Shyshkin, 2012. Systemic Conditionality of Morphogenesis and Its Manifestations in the Paleontological Record
Shishkin M. A. Sistemna obumovlenist formoutvorennia y ii proiavlennia v paleontolohichnii litopysi // Paleontolohichnyi zhurnal, 2012, No. 4, p. 3–15.
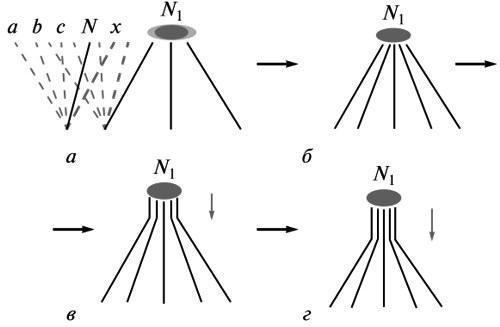
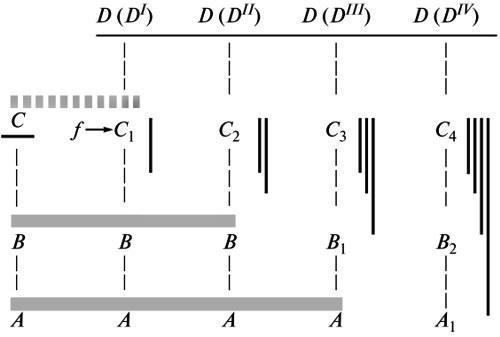
SYSTEMIC CONDITIONING OF MORPHOGENESIS AND ITS MANIFESTATIONS IN THE PALEONTOLOGICAL RECORD © 2012 M. A. Shishkin The general basis of form-formation mechanisms is ensuring the directionality of individual development as a whole toward the adult norm. The stability of this goal-setting is the main property of a living organism that requires explanation. Every step of evolution is a forced reorganization of ontogenesis as a system toward a new ultimate goal (a state of equilibrium), i.e., a new norm. The increase in the stability of its realization across successive life cycles under the influence of selection leads to a restructuring of the development of this evolutionary innovation, spreading in the direction of the early stages. As a result, the difference between the developmental record of the innovation and the initial ancestral ontogenesis is increasingly transformed from a protracted (recapitulative) into a divergent one, corresponding to Baer’s law. Changes associated with this stabilization process are recorded in the paleontological record as a series of characteristic manifestations — the initial instability of newly acquired traits, the restructuring of their morphogenesis over time, and the predominant preservation of recapitulations of ancestral traits in the closest descendants. I. The integrity of development as a controlling factor in morphogenesis The search for an explanation of any phenomenon is possible at different levels of analysis, corresponding to its division into increasingly specific components. The choice of each of these levels may be justified for specific research tasks. However, the substantive laws describing the essence of a phenomenon as a whole pertain to the highest of these levels. Only in this light can the nature of the causal relationships revealed by more specific reductionist approaches be assessed. The essence of scientific knowledge lies precisely in the fact that phenomena that seemed independent are found to be interconnected and subject to general laws. An awareness of this order of things is inevitable for any field of research, and theoretical biology is a striking example of this. Thus, attempts to reduce embryonic development to a mosaic of cellular determination pathways, characteristic of the second half of the 19th century, have given way to the concept of a unified morphogenetic field controlling development at all its stages. Today, the prevailing evolutionary mindset is undergoing a similar transformation; as recently as forty years ago, it believed in the omnipotence of the reductionist principles of neo-Darwinism. This is being replaced by the realization that “macroevolution cannot be reduced to a theory in which the starting concepts are the properties of individual genes”, and that a proper theory must “take into account the requirements of higher levels of analysis” (Maderson et al., 1982). This predetermines the proper approach to understanding the action and formation of the formative mechanisms of individual development—whether we are talking about processes observed in modern organisms or their traces in the paleontological record. The entire diversity of formative acts carried out in the course of normal development, is in one way or another harmonized within its framework, i.e., subject to the laws governing ontogenesis as a whole. Among the latter, the most obvious characteristic is the orientation of the entire process toward a predetermined end result, i.e., the presence of goal-setting (teleonomic) control, as was first emphasized by K. Bear. This integral property of development should serve as the starting point for analyzing all its individual processes. Goal-directed behavior is a property of systemic objects, expressing their capacity for self-regulation. There is no doubt that individual development is a holistic dynamic system, which consistently corrects its state through feedback toward a final equilibrium, i.e., the adult norm (Waddington, 1957; Svetlov, 1972; Shishkin, 2006). All manifestations of developmental integrity identified by embryology are ultimately an expression of its purposefulness. First and foremost, this is the reduction in the range of variability during development, first established by Baer. Second, it is the phenomenon of equifinality or the stability of the outcome of the process with respect to the means of its implementation (Roux, 1896; Driesch, 1908). This property has many manifestations concerning both adults and “programming” provisional stages of development, for example, the mode of gastrulation in many protostomes. These properties can be observed both experimentally (different pathways of individual regeneration, the development of fully formed embryos from separated blastomeres, or the self-assembly of the latter into a single entity) and in normal development. The latter case primarily includes alternative modes of reproduction (sexual and asexual) or types of development (direct or with metamorphosis) in the same or closely related species, whereby the initial stages of the ontogenetic cycle may be extremely different. In a broader sense, the phenomenon of equifinality manifests itself at the level of historical changes, when the same organizational property can be realized in the development of ancestors and descendants on different formative bases (see below). When considering the specific processes that constitute development, their goal-setting nature is most evident in regulatory-type morphogeneses, manifesting itself, for example, in such phenomena as the multiple provision of formative acts or the ability of a competent embryo to exhibit the same reaction in response to a wide range of experimental inductive influences. The main generalization of embryology concerning the directionality of ontogenesis simultaneously expresses its characteristic understanding of the mechanism of this process. This is the concept of holistic determination of development, already present in the zygote and predetermining the formation of all organizational features according to the principle from the general to the particular (Baer, 1828; Gertwig, 1895; Svetlov, 1964). The entire course of events, in the words of P.G. Svetlov, is viewed here as the “inscription” of successive acts of differentiation within the framework of the general developmental plan that controls them. Development is understood as epigenesis, i.e., the accumulation of the diversity of its constituent factors within the framework of their integral interdependence. This principle implies the absence of direct causal relationships between the properties of the organism and the specific structures of the germ cell. As is well known, the search for such relationships constituted the essence of early hypotheses about the mechanism of development, based on “cellular” preformationism, i.e., reducing the entire ontogenesis to a mosaic aggregate of types of cellular differentiation. It was assumed that these were controlled separately by certain cellular components—either cytoplasmic or nuclear (Roy Lancaster’s theory or Ru-Weismann’s mosaic theory; see Svetlov, 1978). This reductionist approach, having proved incompatible with the facts of embryology, subsequently found new ground in chromosomal genetics — during a period when its primary task was the direct mapping of genes to traits of the adult organism using hybrid analysis. As genetics became involved in the analysis of developmental processes, these views were transformed (theoretically) toward greater adaptation to the actual picture of ontogenetic events 1. Today, they are reflected in concepts of gene networks, where the action of higher-order regulatory genes triggers sequential stages ofcascading expression of target genes (e.g., Gilbert et al., 1997). Yet despite the complexity of such schemes compared to the straightforward preformationism of the past, they remain within the framework of what R. Goldschmidt (Goldschmidt, 1940) called the principle of “integrated gene action,” i.e., the notion that adult organization arises from the interaction of independent factors. The fact that the role of a trigger mechanism is assigned in these models to only a small number of initiating genes does not take them beyond the idea of mosaic determinism of development. The latter remains at odds with the idea of holistic control of the ontogenetic process. In fact, the very postulate of the independence of factors that coordinately trigger development, by definition, precludes an answer to the question: what coordinates them? However, in the real world of science, a researcher’s way of interpreting facts may deviate significantly from the theoretical postulates they accept (cf. Shishkin, 2010). For this reason, the current body of genetic knowledge contains numerous generalizations that express an alternative idea of holistic determination. T. Morgan (1924) already suggested that the discreteness of hereditary factors does not necessarily contradict the idea that every property of an organism is determined by the entire germplasm. He also proposed the idea that the differential activity of genes in different cell types is linked to the placement of equipotential nuclei into different cytoplasms—a view now shared by most geneticists (Korochkin, 2002). In essence, this is a recognition that the behavior of a cell’s genome is controlled by its holistic properties—whether these are called the cytoplasmic environment, the cellular environment, or something else. This conclusion is supported by numerous experimental findings regarding the cell’s influence on the structure and function of its genetic apparatus. It has been shown that all stages of matrix synthesis are potentially ambiguous, including both various possibilities for transcript editing and their subsequent translation and post-translational modifications of protein molecules—depending on the physiological conditions within the cell (Inge-Vechtomov, 1976; Golubovsky, 2001). Consequently, the outcome of synthesis is always determined by the nature of its intracellular regulation throughout the entire process. Furthermore, many experiments, particularly on single-celled organisms, have shown that a cell is capable of locally reorganizing its genome to meet the demands of its functioning in a specific situation (Cairns et al., 1988; Hall, 1990). These phenomena reflect the cell’s evolutionarily developed capacity for adaptive modifications at the genetic level (although to the geneticists who discovered it, it usually appears as striking evidence of “adaptive mutations” purposefully carried out by the organism). Thus, the cell is “capable of responding to environmental challenges through active genetic exploration,” i.e., “acting purposefully” in accordance with prevailing conditions (Golubovsky, 2001). All of this shows that the genetic activity of a cell throughout its life is governed, in principle, by the same systemic regulatory mechanism (which determines the appropriate course of events), as is ontogenesis as a whole. This seems to us to be further evidence that the entire body of genetic generalizations will inevitably be reinterpreted over time in light of the laws governing individual development (cf. Shishkin, 2006, 2010). As it pertains to our objectives, we may conclude that in both cases under comparison, a controlling factor of integrity is present from the outset. At the cellular level, its specificity evidently corresponds to their specific generations, while in the organism as a whole it appears as a sequence of states characterizing the successive phases of ontogenesis. This latter conception corresponds to A. Gurwitsch (Gurwitsch, 1922) regarding a transformable biological field, each state of which is an invariant determining the course of development for its immediate segment. Consequently, the change in the field’s characteristics during development is a function of the path traversed by its substrate. The field concept appears to be, for embryology, “the only concept for a holistic causal study of the course of ontogenesis,” allowing us to explain its directionality (Svetlov, 1964). Evidently, it is here that we should seek the mechanism of ontogenetic goal-setting, which determines the course and coordination of individual formative processes in normal development. II. Evolution as a Change in Goal-Setting within the System of Ontogenetic Realization “Purposiveness is not an inherent property of the living” (Schmalhausen, 1982) and must be the subject of an evolutionary explanation. If evolution is the transformation of ontogenesis as a whole, then each of its steps signifies a restructuring of the developmental system toward a new final equilibrium (“goal,” or adaptive norm) in place of the lost one. The system thereby restores its temporarily disrupted stability. According to the epigenetic theory of evolution, which postulates such a course of events, this occurs on the basis of the “available possibilities” of the initial reaction norm, i.e., through the selective preservation of one of the terminal labile aberrations feasible within the framework of this norm (Shmalgauzen, 1982; Shishkin, 1988a, b, 2006, 2010). This aberration is the model of the new nearest state of equilibrium (Fig. 1, a). It determines the direction of correction of the properties of the developmental system during selection. Fig. 1. Evolutionary change as the stabilization of a new “goal” of development: a — the space of possible developmental paths within the initial norm N (left) and within the initial shift toward the new norm N1 (center); formation of a new norm based on similar individual variations preserved by selection (right); b, c, d — successive stages of stabilization of the new norm’s development as the process spreads (across generations) from the final stage to increasingly earlier ones. Notations: a, b, c — potential terminal aberrations of the initial adult norm; N — initial norm; N1 — new norm at different stages of its formation; x — one of the aberrations of the initial norm, preserved by selection under new conditions. Horizontal arrows — stages of stabilization of change; vertical arrows — direction of historical stabilizing transformations. Thus, the path of formation of each new type of goal-setting (modified norm) is the history of increasing the precision of “guiding” development toward a new selected goal (Fig. 1, a–d). Conversely, moving back to the early stages of this process, we should observe an ever-increasing dispersion of individual developmental variants around this future goal. The initial stage should correspond to the maximum lability of the aberration preserved by selection. This is expressed in the extreme ambiguity of the reproduction of its phenotype in direct offspring. From this, the direction of change in the evolution of morphogenesis becomes clear. Each step toward stabilizing a new goal involves the preferential survival of individuals who have successfully replicated the phenotype of their previously selected parents. Thus, preference is given to those individual “equifinal” variants of the offspring, in which the immediate formative basis of the preserved phenotype proved to be the least sensitive to fluctuations in developmental factors (a process described by Schmalhausen as a transition from the dependent implementation of innovation to an increasingly autonomous one). Among the descendants of these variants, those in which the innovation’s resistance to interference proved to be greatest will again gain a selective advantage—due to the involvement of increasingly earlier ontogenetic processes in its integrated morphogenesis (Fig. 1, b, c). Thus, as the new adaptive norm stabilizes, the restructuring of its morphogenesis spreads across generations toward earlier stages of development. As this process deepens, its morphological expression inevitably spreads in the same direction (Shishkin, 1988a, b, 2006, 2010). In short, the history of the emergence of any innovation is a chain of successive tests by the selection of its carriers based on their ability to reproduce themselves regardless of disturbances in the course of development. The selection process acts here as a sequence of damped correction cycles leading to the stabilization of a new norm. The fact that the morphogenetic transformations of long-standing structures proceed in the manner described above, encompassing increasingly earlier stages of development, is well known from the experience of comparative embryology and paleontology (although here it is usually not elementary evolutionary events that are available for assessment, but their cumulative result, unified by a common vector of change). The most obvious pattern observed in these cases is that organizational properties, which arose under the conditions of the ancestor’s adult life and were undoubtedly morphogenetically dependent on them, manifest in descendants much earlier and more autonomously—long before the onset of conditions conducive to the functioning of these properties. For example, the shell and foot in mollusks, which arose as adaptations of the adult animal to life on the seabed, usually appear in modern forms as early as the pelagic larval stage, unrelated to the conditions of their evolutionary origin. Similarly, in modern birds, the specificfeatures of the wing skeleton, which developed in the course of adaptation to flight in their ancestors, are already evident at the embryonic stage, unrelated to flight. Stabilizing transformations of this kind, reflecting a profound restructuring of the morphogenesis of the adult structure while preserving the final plan of its structure, are described in the literature under a variety of names — such as acceleration, condensation, adultation, embryonization 2, etc. (Shishkin, 2006). More indirect, yet equally clear, evidence of this evolutionary path of morphogenesis lies in the numerous parallels between variations within certain taxa and diagnostic features in their closest relatives. Obviously, the second state can be interpreted as the result of the historical stabilization of the first (see below). This direction of events in stabilization processes sheds light on the true meaning of Baer’s law (the increase in ontogenetic divergence between related forms during development). Many authors consider it “fundamentally incompatible” with E. Haeckel’s biogenetic law, arguing that Haeckel’s recapitulations of adult ancestral states are in fact merely manifestations of the embryonic similarity between ancestor and descendant, described by de Beer’s rule (de Beer, 1971; Gould, 1977). In fact, both of these generalizations are valid, characterizing in an extremely idealized form the two sides of the evolutionary process (Shishkin, 1988b). De Beer’s law raises no questions only with regard to developmental types that diverged from a common ancestor; but when comparing ontogenies belonging to a single phylogenetic line, the possibility of their divergence (rather than the prolongation of one by the other) is not self-evident and requires explanation. Evolution begins with terminal changes in organization. The first steps of this selective process, which fixes one of the later aberrations of the existing norm, initially create thereby only a new modification of the latter, realizable only under these altered conditions (Schmalhausen, 1982; cf. above). That is, in accordance with Haeckel’s principle, the previous norm is largely preserved here as a subterminal stage of development, serving as the basis for a new adaptive response (Fig. 2, C–D(D I )). However, as this reaction transforms into an unconditional, stable norm, its autonomy from the environmental factors that initially induced its morphogenesis increases. Thus, the ancestral stage that served as the starting point for the new norm—which was more or less fully reproduced at the previous stage—must be transformed to provide the formative mechanism for this change (Fig. 2, D(D II )). As the new norm further stabilizes in descending descendants, this restructuring of morphogenesis will intensify, simultaneously extending to increasingly earlier stages (see above). As a result, the recapitulation of the ancestral state will be increasingly erased, while the divergence between the initial and derived variants of ontogenesis will deepen (Fig. 2, C–D (D IV ) ). Fig. 2. Deepening divergence between the development of the ancestor and that of descending descendants during the restructuring of the morphogenesis of an evolutionary innovation. Symbols: A, A 1 ; B, B 1 , B 2 ; C, C 1–4 — developmental stages; D(D I )–D(D IV ) — historical stages of innovation stabilization; f — external factor initially influencing form formation. Gray horizontal lines — successive levels of divergence; dark vertical lines — accumulation of stabilizing changes (after Shishkin, 1988b, with modifications). Thus, the evolution of new acquisitions is the history of the continuous transformation of their ontogenetic record from a prolonging (according to Haeckel) to a divergent (according to Ber) one in comparison with the ontogenesis of the original ancestral form (Shishkin, 1988b). From the above, it is clear that the smaller the phylogenetic distance between ancestor and descendant, the greater the likelihood that traces of the ancestral adult state may be preserved in the descendant’s late ontogenesis, and vice versa. This pattern, well known from the study of fossil remains (see, for example, Ruzhentsev, 1960), explains why the main criticism of the biogenetic law comes from specialists in extant organisms, whereas paleontologists view it much more positively. Indeed, in an analysis that includes modern forms, the objects of comparison are organisms separated by vast intervals of evolutionary transformation, not to mention the predominantly nonlinear nature of their phylogenetic relationships. In contrast, the paleontologist often deals with more or less continuous links within a singlephylogenetic lineage, where the repetition of ancestral traits in the late ontogenesis of a descendant appears in many cases as an obvious fact. III. Evidence of the evolution of morphogenesis, as recorded in fossil remains From the above, a set of patterns in the evolution of morphogenesis emerges, the manifestations of which can be expected with a high degree of probability when comparing fossil organisms. These include, first and foremost, the following. (1) The emergence of innovations in the form of unstable variations—not only in terms of the frequency and degree of their manifestation, but also in relation to other features of organization. (2) The historical stabilization of these acquisitions, expressed (in addition to the actual increase in their stability) in a restructuring of their mode of development and in their increasingly early visible manifestation in ontogenesis 3. (3) A high probability of the preservation of adult ancestral traits in the late ontogenesis of relatively close descendants, and thereby—the possibility of identifying in them ancient recapitulations that have been erased in later types of development. As already noted, in the last of these cases, fossil material has an indisputable advantage over recent material in terms of its informativeness; but with regard to patterns (1) and (2), this difference is not so obvious. The paleontological record preserves mainly only established morphotypes, i.e., those that were able to persist over time and space. At the same time, the early stages of their formation, characterized by rapid transformation and occurring in local populations within limited territories, leave little direct evidence. These stages are detected mainly indirectly—in the form of fading traces of ancestral variations preserved in the development of more stabilized descendants (especially in pedomorphic lineages). Similar caveats are necessary regarding the stabilizing restructuring of the development of new acquisitions. Evidence of this process, as observed in the fossil record, is limited mainly to its morphological manifestations and the extension of these to earlier stages. The actual changes in the factors of morphogenesis (available for comparative study in extant forms) can be traced only in rare cases. Furthermore, as noted above, the material for assessing stabilization processes in the fossil record is usually not individual evolutionary shifts, but their cumulative outcomes within unidirectional sequences of phylogenetic changes.
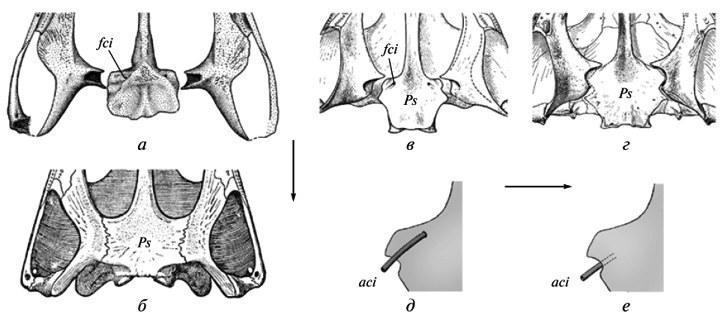
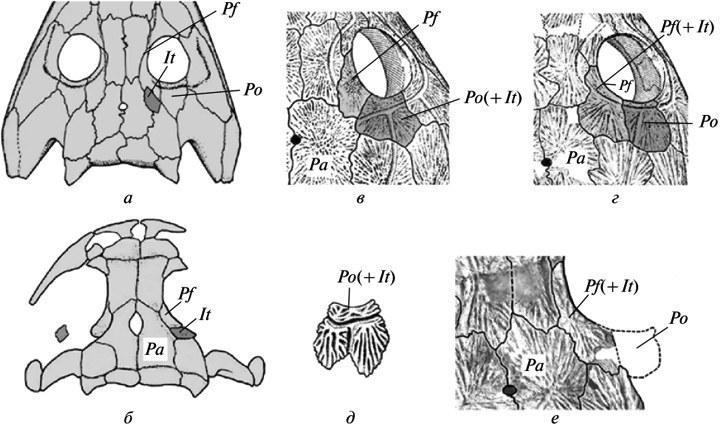
Sources of data on the course of developmentavailable in the fossil record can be summarized into several main categories (Makridin, Shishkin, 1988). These are, first, age series or their individual key stages (e.g., larval morphotypes in insects or embryos in amniotes). Rough models of age series, which in some cases provide information on qualitative transformations (often unavailable in natural growth sequences), can be derived from comparisons of related forms with varying degrees of pedomorphic underdevelopment (Shishkin, 1973). Another extensive source consists of changes associated with the accretionary growth of the skeleton, i.e., the process of adding new parts without resorption of the previous ones. This type of skeletal formation is known to varying degrees in most groups of invertebrates. It suffices to mention the development of the shell in cephalopods — a source of classic examples of heterochrony and recapitulation in ancient organisms. Closely related to accretion is skeletal growth through the addition of new elements, as is characteristic of echinoderms. A distinct variant of accretion is the growth of colonial organisms (corals, bryozoans, graptolites, etc.), where the object of study may be both the life cycles of the individuals comprising the colony (hysterogenesis) as well as the colony’s development as a whole (astogenesis). In this context, the course of historical changes in astogenesis can be assessed in the same categories—in particular, such as heterochrony — as in the evolution of individual development. A specific subject of analysis may be developmental anomalies, which represent natural analogues of experimental disruptions in morphogenesis studied in modern organisms. For each developmental system, such deviations collectively express its specificity (“phase portrait”), or the space of potentially realizable ontogenetic trajectories. It is always bounded, structured, and is a mediated product of the evolution of normal development, constituting its broader characterization (Waddington, 1957; Shishkin, 1988a, 2006, 2010). Evolutionary changes in this space imply a greater or lesser similarity in the pattern of possible developmental trajectories among related taxa, with each of them potentially being stable (normal) in one instance and aberrant in another 4 . In fossil organisms, examples of anomalies or simply variations that replicate the normal features of related taxa are just as common as in modern ones (see, for example, Shishkin, Makridin, 1988, Fig. 50). In particular, among ammonites, there are known cases where the shell sculpture on one side is normally developed, while on the other it follows a pattern known from a closely related species (Tasnadi-Kubacska, 1962, Fig. 82). In addition to demonstrating hidden developmental potentials, such cases also shed light on features of form formation not captured in the traces of typical ontogenesis. Thus, in this example, the autonomous functioning of the mantle epithelium (forming the ornamentation) on both sides of the shell’s axis of symmetry is self-evident. Sometimes anomalies allow us to document early features of normal morphogenesis, which, in typical cases, are no longer evident at the stage of hard skeleton formation. For example, in Triassic amphibians of the Tupilacosauridae, the components of diplospondyl vertebrae ossify as discs that do not retain traces of division into two antimeres. However, this initial paired structure can be detected in cases of abnormal vertebral fusion—when individual discs are represented by only one half. All these traces of morphogenic processes provide a basis for documenting the evolutionary patterns listed above. Let us consider some examples. Fig. 3. The transition from the superficial arrangement of the internal carotid arteries (a, c, e) to their longitudinal ingrowth into the skull base (b, d, f) in the evolution of temnospondyl amphibians. The base of the skull (a–d) and part of the parasphenoid (e, f) from the palatal side: a — primitive state with superficial arrangement of arterial traces (Onchiodon); b — derived state (Benthosuchus); c, d — primitive state in the trimerorachoid Dvinosaurus primus and advanced state in D. egregius; e — reconstruction of the artery segment in the region where it enters the skull in Dvinosaurus primus; f — the same in D. egregius. Abbreviations: aci — internal carotid artery; fci — artery entrance; Ps — parasphenoid. Arrows indicate the direction of changes (a — after: Boy, 1990, with modifications; b — after: Bystrov, Efremov, 1940; c–d — after: Shishkin, 1973). Initial instability of evolutionary changes. This phenomenon manifests itself in various aspects of the skull evolution of ancient temnospondyl amphibians. In particular, the consolidation of its palatal ossifications led to a change in the topography of the internal carotid arteries. Initially, before entering the endocranial cavity, they were located openly on the palate (Fig. 3, a, e); but subsequently their channels became entirely incorporated into the parasphenoid in the preauricular region—a condition typical of Triassic forms (Fig. 3, b, e). The transition from the first structural stage to the second occurred rapidly in various ways; therefore, it usually leaves no traces in the age-related stages available for study. However, in the development of early forms of the second type (Permian Acanthostomatops: Witzmann, Schoch, 2006, Fig. 5) variable recapitulations of the initial proportions may persist. Variability in the transitional adult state (asymmetry of arterial openings) is known in the Permian melosaurid Tryphosuchus. Finally, an indirect expression of the instability of the transition to the advanced type is observed in neotenic Dvinosauridae, where ambiguity regarding the adult state exists at the genus level. Specifically, in the genus Dvinosaurus, two alternative types of arterial arrangement, which normally characterize different evolutionary stages, appear as merely one of the species-level differences (Fig. 3, c-e) — against the backdrop of a preserved primitive skull structure. These relationships constitute a typical example of “archaic diversity” (Mamkaev, 1968)—a phenomenon characterizing the early, unstable stages of the formation of new organizational plans. Although the Dinosauridae themselves are a terminal Late Permian group, they most likely gave rise to some Laurasian Triassic “brachiopids.” Another similar example is the loss in various lineages of Permian-Carboniferous temnospondyls of the intertemporal bone in the cranial vault (through its normal fusion with the parietal bone). The intermediate stages of this process have not been identified. In the earliest stages of ontogenesis available for study (the onset of ossification), its normal recapitulations have already been lost—even in forms studied through fairly complete age series, such as the brachiosaurs. Nevertheless, as a rare variation, these bones may sometimes retain their independence in juvenile individuals—on one or both sides of the skull (Fig. 4, a, b). In other words, their mesenchymal primordia persisted during early development, sometimes reaching ossification due to random delays or disruptions in fusion processes (Boy, 1988, Fig. 2B; Schoch, 2004, Fig. 1A, “po”). Fig. 4. Instability of the early stages of intertemporal bone loss in the evolution of the skull roof in temnospondyl amphibians: a — preservation of the bone as a variation in the eriopoid Sclerocephalus; b — in the branchiosaur Apateon; c — fusion with the postorbital bone in one species of Dvinosaurus (D. primus); d — fusion with the postfrontal bone in the species D. egregius; e — preservation of traces of the intertemporal bone’s distinctness within a complex postorbital bone in a juvenile D. primus; f — fusion of the intertemporale with the postfrontale as a variation in the same species. Abbreviations: It — intertemporale; Pa — parietale; Pf — postfrontale; Pf(+It) — fused postfrontale and intertemporale; Po — postorbital; Po(+It) — fused postorbital and intertemporal (a — after: Boy, 1988, with modifications; b — after: Schoch, 2004, with modifications; c, d — after: Shishkin, 1973; e — after: Bystrow, 1938). This initial instability of evolutionary change, obscured by the incompleteness of the record against a backdrop of rapid transformations, is again much more clearly demonstrated in the neotenic Dvinosaurus, where changes proceeded more slowly. The preservation of the intertemporal bone in the early stages of development is evident from the presence of traces of its separate ossification center in late juvenile specimens, even despite the loss of the bone’s independence (Fig. 4, e). It is particularly interesting that in the basal species D. primus, the latter is typically assimilated in a manner unusual for Temnospondyli, through fusion with the preorbital bone (Fig. 4, c). At the same time, an alternative fusion with the postfrontal bone occurs as a variation, which becomes the norm in the subsequent species D. egregius (Fig. 4, d, e). All of this suggests that the multidirectionality and instability of events on the path toward the eventual loss of the intertemporale were in fact typical of this transformation in the history of the Temnospondyli group as a whole. Evidently, such a developmental path is characteristic of any evolutionary innovations. The latter, in fact, is what is implied by the principle of “archaic diversity.” Stabilization of evolutionary acquisitions. Manifestations of increased variability in the early history of evolutionary acquisitions already in themselves indicate the gradual nature of their stabilization. This applies both to the examples cited above and to many others known from the fossil record (Shishkin, 1988a, p. 168; 1988b, p. 193). An even more frequently observed consequence of stabilization processes is the acceleration of the development of new features, leading to their appearance at earlier ontogenetic stages (see Section II).
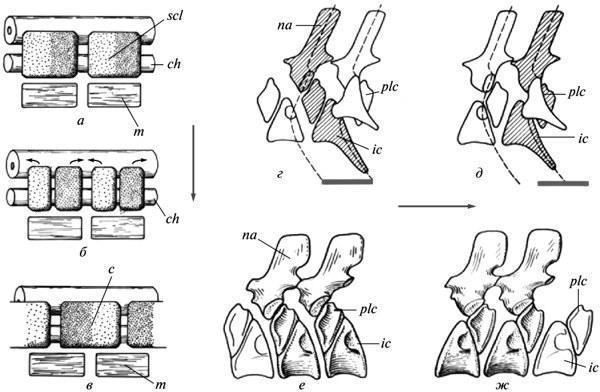
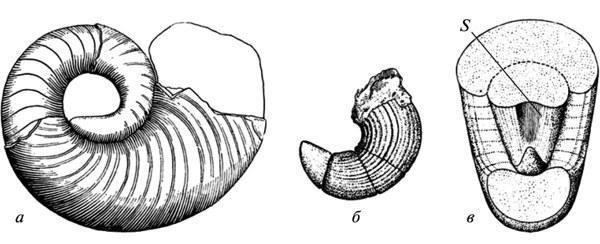
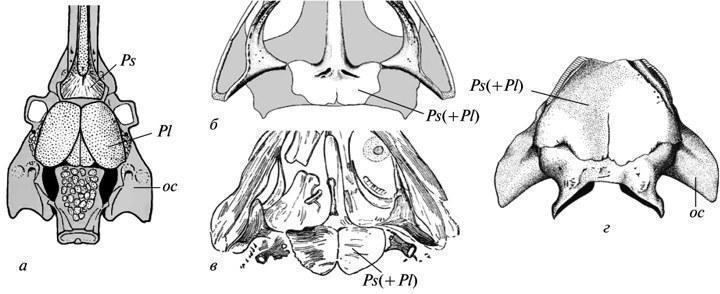
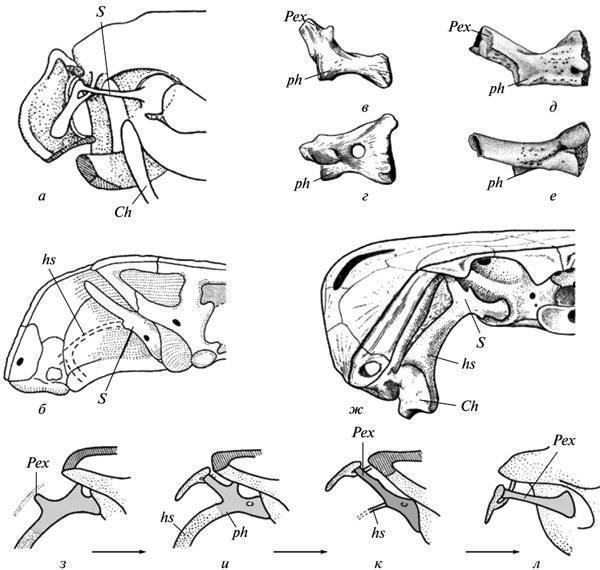
Both of these patterns imply a restructuring of form formation, but the possibilities for directly assessing its mechanisms based on fossil remains are quite limited. One of the rare cases allowing us to observe the autonomization of the morphogenetic process from the exogenous factor that initially induced it was cited in the 19th century by A. Hayett regarding the evolution of the shell of Paleozoic nautiloids. It was shown that the appearance of indentations on the dorsal side of a growing shell was initially caused by its tight contact with the preceding whorl. However, in later forms, this feature no longer exhibits such a dependence and is established directly from the initial stages of growth, not yet affected by the closure of whorls (Fig. 5). Similar manifestations of developmental autonomy from initial inducing factors are well known in the evolution of modern morphogenesis as well (Shishkin, 1988b, p. 176). Fig. 5. Autonomization of the development of longitudinal indentation in the shell wall in Paleozoic nautiloids: a — early form (the trait appears as a result of the closure of a new shell whorl with the previous one); b, c — latest form (the appearance of the feature precedes the closure). Legend: s — longitudinal indentation on the dorsal wall of the initial whorl of the shell (after: Hyatt, 1893 from Gould 1977, with modifications). Changes in morphogenetic relationships can also be illustrated by alterations in the development of the cranial bones bearing the lateral line organs during the transition from fish to ancient amphibians. In the lobe-finned ancestors of tetrapods, as in most modern bony fish, the placodes of the lateral line organs undoubtedly served as inductors for the formation of the “canal-bearing” cover bones developing around them. Accordingly, the chains of seismic sensory organs in the head were located in these fish within channels passing through the centers of formation of the corresponding bones. In Paleozoic amphibians, this connection was lost, i.e., the bones were laid down independently of the lateral line system. The latter formed more superficially in the epidermis and could come into contact with the bones only at later stages of development. The coordination of these two morphogenetic processes was partially preserved, since the lateral line grooves still mainly passed over the centers of development of the former “canal” bones. However, this relationship was no longer as rigid, allowing for the displacement of individual grooves onto other bones (cf. Shishkin, 1973, fig. 65, p. 170). Recapitulation of ancestral traits (palingenesis) preserved in primitive developmental patterns. Among the features of adult organization characteristic of ancient vertebrates, many either left no traces at all in modern ontogenies, or these traces do not lend themselves to unambiguous interpretation. But in the more recent descendants of ancient forms, such features could have been imprinted in development much more clearly. This is particularly evident from the following data on the development of ancient amphibians. As shown by E. Jarvik, the parasphenoid in tetrapods arose through the fusion of the parasphenoid proper of their lobe-finned ancestors (which underlay the trabecular region of the skull) with the posterior paired parietal plates belonging to the exoskeleton of the hyoid arch (Fig. 6, a). In the development of modern tetrapods, traces of the paired nature of the posterior part of the complex parasphenoid have been lost, including in primitive Urodela (cf. Lebedkina, 1979). In Paleozoic temnospondyl amphibians, by the onset of ossification, this condition normally also left no traces; but it undoubtedly persisted at the stage of mesenchymal formation of the palatal exoskeleton. This is evidenced by variations in which the phase of cleavage of the posterior part of the parasphenoid was individually prolonged, persisting during the stages of palatal ossification. Such examples are not uncommon in the development of branchiosaurs (Fig. 6, b, c). Among eriopoids, a similar delay has been documented in one case in a fully formed juvenile specimen with an ossified endocranium (Fig. 6, d). Fig. 6. Traces of recapitulation of the parietal plates of the palate in lobe-finned fish in the development of the parasphenoid in temnospondyl amphibians. Cranial structures from below (a–d): a — the crossopterygian Eusthenopteron (occipito-auricular and part of the ethmosphenoid regions of the endocranium); b, c — the branchiosaur Apateon and a closely related form (posterior part of the palate); d — juvenile erioid “Actinodo n” (occipito-auricular region of the endocranium). Abbreviations: oc — auricular capsule; Pl — parotico-temporal plate; Ps — parasphenoid; Ps(+Pl) — tetrapod parasphenoid complex (a — after: Jarvik, 1954; b — after: Boy, 1978; c — after: Watson, 1963; d — after: Boy, 1971; all with modifications). Among the long-standing problems of evolutionary morphology discussed in the context of developmental data is the question of the homology of the auditory ossicle in tailless amphibians. Unlike the reptilian stapes, which develops from the hyoid arch, in tailless amphibians this bone arises from the wall of the ear capsule (or independently), without connection to the hyoid. Therefore, many researchers have regarded it as a neoplasm (“otostapes”) that arose independently of the “hyostapes” of reptiles. Meanwhile, in the direct ancestors of tailless amphibians—the Paleozoic batrachomorphs (Temnospondyli)—the pattern of stapes development, being much more archaic, clearly indicates the hyoid origin of this element. In adult forms, the latter is quite similar in structure and topography to its homolog in Anura (Fig. 7, a, b, k, l); the hyostapediary connection could have persisted here only as a rudimentary ligament. But in early stages, and in neotenic forms—sometimes even in the adult state— it was represented by a robust cartilaginous infrastapediary process of the stapes, which had a broad base and recapitulated the hyomandibular stem of the palmate-footed ancestors (Fig. 7, c–g). Fig. 7. Recapitulation of the hyostapediac connection in the ontogeny of ancient batrachomorphs (b–k): a—middle ear region in modern tailless frogs (Rana); b—ear region of the occiput in temnospondyls (Lydekkerina); c–e — the auditory ossicle in the same group in neotenic forms (Dvinosaurus: c, d) and at early stages of development (Sclerocephalus: e, f), viewed from the rear (c, e) and from above (d, f); g — reconstruction of the hyostapediary connection in the skull of Dvinosaurus; h–l — reconstruction of the path of transformation of the hyomandibula into the auditory ossicle in batrachomorphs — from the earliest amphibians (h, i) to the typical temnospondyl structure (k) and the relationships in tailless (l). Abbreviations: Ch — ceratohyale; hs — hyostapediary cartilage or ligament; Pex — processus externus (extrastapedialis); ph — processus hyoideus (infrastapedialis); S — stapes (a — after: Eaton, 1959; b — after: Parrington, 1948; d, e — after: Boy, 1988; c, d, f–l — after Shishkin, 1973, with modifications). One of the controversial features of the embryogenesis of modern amniotes is the resegmentation of their spine. The vertebral body primordia arise here from two adjacent halves of the sclerotomes of neighboring segments, as a result of which each vertebra develops in association with two consecutive myotomes (Fig. 8, a–c). The origin of this rearrangement cannot be explained by comparing development in modern tetrapods. In recent amphibians, it is obscured by the late formation of sclerotomes and is therefore often questioned. The key to understanding the nature of this phenomenon is almost certainly provided by the relationships among ancient apsisospondyl (complex-vertebrate) amphibians, specifically within their batrachomorphic clade—the Temnospondyli. In these forms, the vertebral body hemicenters could merge into functional units via two alternative pathways—intrasegmentally and transsegmentally, i.e. by integrating semi-centers from adjacent metameres (Fig. 8, d–e). The transition from the first type to the resegmented second type occurred repeatedly in the history of the Temnospondyli, taking place during late development; meanwhile, in the neotenic Dvinosaurus, both variants are present, represented by different species. Occasionally (in Triassic metapodials), a transition from the original variant to the derived one can be observed within the same vertebral column (Shishkin, 1987). Fig. 8. Embryonic resegmentation of sclerotomes in amniotes as a reflection of the recombination of vertebral hemicenters in ancientapsidospondyl amphibians: a–c — stages of resegmentation in modern amniotes (schematic); d, e — intrasegmental and transsegmental vertebral metameres in temnospondyl amphibians (schematic); f — intrasegmental vertebrae in Dvinosaurus egregius; g — transsegmental vertebrae in D. purlensis. Symbols: c — vertebral body primordium; ch — notochord; ic — intercenter; m — myotome; na — neural arch; plc — pleurocenter; scl — sclerot; arrows indicate the direction of resegmentation; gray lines — extent of the functional segment (a–c — after: Romer, 1968; d–g — after: Shishkin, 1987). Given the vast gap between modern amniotes and ancient amphibians, as well as the fact that Temnospondyli are not the ancestors of amniotes, one might certainly question the continuity between the two types of resegmentation discussed. However, it is more reasonable to assume that in early terrestrial reptiloid amphibians (the ancestors of amniotes), resegmentation began in the same way as in batrachomorphs, but then the acceleration of this process led to its embryonization as early as the Paleozoic. The presence of both “batrachomorphic” types of vertebral integration in the limbed ancestors of tetrapods also indirectly supports this view (Shishkin, 1987). From this perspective, we must conclude that while the initial objects of resegmentation were the bony or cartilaginous components of the definitive apidospondylous vertebra, then its acceleration and intensification led to a state where the material of embryonic sclerotomes became its direct substrate. This restructuring made the entire process independent of the formation of vertebral semicenters, of which in amniotes practically only one (plevrocenter). Taken together, this provides a vivid example of the equifinality of a stable evolutionary acquisition, where, while remaining historically unchanged in its main outcome, it radically alters the manner of its realization. All the examples of morphogenesis in ancient organisms discussed here (evaluated by comparing them with one another or with modern developmental patterns) shed light on some of the main patterns of the historical stabilization of form formation. The latter is traced here both in the form of traces of the initial instability of future typical organizational features and in examples of large-scale historical changes leading to a qualitative restructuring of the mechanism of development of such features. As shown above, these phenomena are characteristic and expected manifestations of the holistic evolution of ontogenesis as a system. REFERENCES Bystrov A.P., Efremov I.A. Benthosuchus sushkini Efr. — a labyrinthodont from the Eotriassic of the Sharzhenge River. // Proceedings of the Paleontological Institute of the USSR Academy of Sciences. 1940. Vol. 10. No. 1. 152 pp.
Gertwig O. Contemporary Controversial Issues in Biology. No. 1. Evolution or Epigenesis. Moscow, 1895. 125 pp. Gilbert S.F., Opitz D.M., Reif R.A. A New Synthesis of Evolutionary Biology and Developmental Biology // Ontogenesis. 1997. Vol. 28. No. 5. pp. 325–343. Golubovsky M.D. Non-canonical Hereditary Changes // Nature. 2001. No. 8. pp. 3–9. Inge-Vechtomov S.G. The Genotype System // Physiological Genetics. L.: Meditsina, 1976. pp. 57–113. Korotchkin L.I. Biology of Individual Development. Moscow: Moscow State University Press, 2002. 263 pp. Lebedkina N.S. Evolution of the Amphibian Skull. Moscow: Nauka, 1979. 283 pp. Makridin V.P., Shishkin M.A. Study of the Ontogenesis of Extinct Organisms // Modern Paleontology / Ed. by V.V. Menner, V.P. Makridin. Moscow: Nedra, 1988. Vol. 1. pp. 197–211. Mamkaev, Yu.V. Comparison of morphological differences in lower and higher groups of a single phylogenetic lineage // Journal of General Biology. 1968. Vol. 29. pp.
48–55. Meyen, S.V. Problems of the Phylogenetic Classification of Organisms // Modern Paleontology / Edited by V.V. Menner, V.P. Makridin. Moscow: Nedra, 1988. Vol. 1. pp. 497–511. Morgan T. The Structural Basis of Heredity. Moscow–Leningrad: Gosizdat, 1924. 310 pp. Ruzhentsev V.E. Principles of Systematics, System, and Phylogeny of Paleozoic Ammonoids // Proceedings of the Paleontological Institute of the USSR Academy of Sciences. 1960. Vol. 83. pp. 1–331. Svetlov, P.G. On the Holistic and Elementalist Methods in Embryology // Archives of Anatomy, Histology, and Embryology. 1964. Vol. 46, No. 4. pp. 3–26. Svetlov P.G. Ontogenesis as a purposeful (teleonomic) process // Arch. Anat. Histol. Embryol. 1972. Vol. 63. No. 8. pp. 5–16. Svetlov P.G. Physiology (Mechanics) of Development. L.: Nauka, 1978. Vol. 1. 279 pp.; Vol. 2. 262 pp.
Shishkin M.A. Morphology of ancient amphibians and problems of the evolution of lower tetrapods // Proceedings of the Paleontological Institute of the USSR Academy of Sciences. 1973. Vol. 137. 260 pp. Shishkin, M.A. The Evolution of Ancient Amphibians // Proceedings of the Paleontological Institute of the USSR Academy of Sciences. 1987. Vol. 225. 142 pp. Shishkin M.A. Evolution as an epigenetic process // Modern Paleontology / Ed. by Menner V.V., Makridin V.P. Moscow: Nedra, 1988a. Vol. 2. pp. 142–169. Shishkin, M.A. Patterns of Ontogenetic Evolution // Modern Paleontology. / Edited by V.V. Menner, V.P. Makridin. Moscow: Nedra, 1988b. Vol. 2. pp. 169–209. Shishkin, M.A. Individual Development and the Lessons of Evolutionism // Ontogenesis. 2006. Vol. 37. No. 3. pp. 179–198. Shishkin, M.A. Evolutionary Theory and Scientific Thinking // Paleontological Journal. 2010. No. 6. pp. 3–17. Shishkin M.A., Makridin V.P. Paleopathological and Paleoteratological Studies // Modern Paleontology / Ed. by Mener V.V., Makridin V.P. Moscow: Nedra, 1988. Vol.
1. Pp. 219–229. Shmalgauzen I.I. The Organism as a Whole in Individual and Historical Development. Moscow: Nauka, 1982. 228 pp. Alberch P. Developmental constraints in evolutionary processes // Life Sci. Res. Rep. 1982. No. 22. pp. 313–332. Baer, K. On the History of Animal Development. Vol. I. Königsberg: Kupfertaf, 1828. 271 pp. Beer, G.R. de. Embryos and Ancestors.
Oxford: Clarendon Press, 1971. 191 pp. Boy J. A Remarkable Skull Fragment of an Early Permian Labyrinthodont (Amphibia) from the Saar Region // Abh. hess. Land.-Amt. Bodenforsch. 1971. Vol. 60. pp. 31–43. Boy J. The tetrapod fauna (Amphibia, Reptilia) of the Saar-Palatinate Rothliegend (Lower Permian, SW Germany). 1. Branchiosaurus // Mainz Geological Bulletin 1978. Vol. 7. pp. 27–76. Boy J.A. On some representatives of the Eryopoidea (Amphibia: Temnospondyli) from the European Rotliegend (?Upper Carboniferous–Permian). 1. Sclerocephalus // Paläontol. Z. 1988. Vol. 62. No. 1/2. pp. 107–132. Boy J. On some representatives of the Eryopoidea (Amphibia: Temnospondyli) from the European Rotliegend (?Upper Carboniferous–Permian).