
Usova, Kravchenko, Shabanov (2015) Внутрішньопопуляційні онтогенетичні стратегії у зелених жаб
Усова Є. Є., Кравченко М. А., Шабанов Д. А. Внутрішньопопуляційні онтогенетичні стратегії у зелених жаб (Pelophylax esculentus complex) // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2015. …
Усова Е. Е., Кравченко М. А., Шабанов Д. А. Внутрипопуляційні онтогенетичні стратегії у зелених жаб (Pelophylax esculentus complex) // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2015. – Вип. 25. – С. 223-238.
УДК: (57.022+57.032):591.84:591.158.1:597.851
Внутрипопуляційні онтогенетичні стратегії у зелених жаб (Pelophylax esculentus complex)
Е.Е.Усова, М.А.Кравченко, Д.А.Шабанов
Харківський національний університет імені В. Н. Каразіна (Харків, Україна)
e.e.usova@gmail.com, marinakravchenko2106@gmail.com, d.a.shabanov@gmail.com
З використанням скелетохронології досліджено 575 представників Pelophylax esculentus complex (193 P. ridibundus, 348 диплоїдних і 34 триплоїдних P. esculentus) з Харківської області. Різноманітність жаб за віком і довжиною тіла описано за допомогою апроксимації методом найменших квадратів двома лініями регресії, що відповідають відносно дрібнішим та більшим особинам одного віку. Ці дві групи особин порівняно за популяційнобіологічними параметрами та зареєстровано синдроми (комплекси пов’язаних ознак), які дозволяють розглядати їх як прояви внутрішньопопуляційних онтогенетичних стратегій (ВОС) малорозмірності та крупнорозмірності. Малорозмірність/крупнорозмірність характеризуються низькою/високою швидкістю росту, раннім/пізнім початком участі у нересті самок і пізнім/раннім – самців, високою/низькою тривалістю життя, низькою/високою плодючістю самок і великою/малою кількістю сезонів розмноження, в яких бере участь особина. За допомогою імітаційного моделювання показано, що відмінності P. ridibundus і P. esculentus за ВОС можуть забезпечувати їх стійке співіснування в геміклональній популяційній системі.
Ключові слова: Pelophylax esculentus complex, скелетохронологія, стратегія, малорозмірність, крупнорозмірність, імітаційне моделювання, геміклональні популяційні системи.
Внутрішньопопуляційні онтогенетичні стратегії у зелених жаб (Pelophylax esculentus complex)
О.Є.Усова, М.О.Кравченко, Д.А.Шабанов
З використанням скелетохронології дослідили 575 представників Pelophylax esculentus complex (193 P. ridibundus, 348 диплоїдних і 34 триплоїдних P. esculentus) з Харківської області. Різноманітність жаб за віком і довжиною тіла описали за допомогою апроксимації за методом найменших квадратів двома лініями регресії, що відповідають відносно дрібнішим та більшим особинам одного віку. Дрібніших та більших особин порівняли за популяційнобіологічними параметрами і зареєстрували синдроми (комплекси пов’язаних ознак), що дозволяють розглядати їх як прояви внутрішньопопуляційних онтогенетичних стратегій (ВОС) малорозмірності та крупнорозмірності. ВОС малорозмірності/крупнорозмірності характеризуються низькою/високою швидкістю росту, раннім/пізнім початком участі у нересті самок і пізнім/раннім – самців, високою/низькою тривалістю життя, низькою/високою плодючістю самок і великою/низькою кількістю сезонів розмноження, в яких бере участь особина. Із застосуванням імітаційного моделювання показали, що відмінності P. ridibundus і P. esculentus за ВОС можуть забезпечувати їх стійке співіснування в геміклональній популяційній системі.
Ключові слова: Pelophylax esculentus complex, скелетохронологія, стратегія, малорозмірність, крупнорозмірність, імітаційне моделювання, геміклональні популяційні системи.
The water frogs' (Pelophylax esculentus complex) intrapopulation ontogenetic strategies
O.E.Usova, M.O.Kravchenko, D.A.Shabanov
575 representatives of Pelophylax esculentus complex (193 P. ridibundus, 348 diploid and 34 triploid P. esculentus) from Kharkivska oblast have been studied using skeletochronology. With approximation by the LS method, frogs’ diversity on age and body length has been characterized by two regression lines, which correspond to relatively smaller and bigger individuals of the same age. We compared these individuals by population‑biological parameters and registered syndromes (complexes of related traits), allowing their consideration as manifestation of intrapopulation ontogenetic strategies (IOS). IOS of undersized/oversized is characterized by relatively low/high growth rate, high/low life expectancy, low/high fertility of females and relatively large/small number of breeding seasons in which the individual takes part. Participation in spawning begins early/late in undersized/oversized females and vice versa in males. By using simulation it has been shown that differences in IOS between P. ridibundus and P. esculentus can provide their sustainable coexistence in the hemiclonal population system.
Key words: Pelophylax esculentus complex, skeletochronology, strategy, undersized, oversized, simulation, hemiclonal population systems.
Вступ
Гібридогенний комплекс зелених жаб, Pelophylax esculentus complex (=Rana esculenta complex) складається з двох батьківських видів: ставової жаби, Pelophylax lessonae (Camerano, 1882), та озерної жаби, Pelophylax ridibundus (Pallas, 1771), а також їх міжвидових гібридів різної плоїдності (Plötner, 2005; Шабанов, Литвинчук, 2010). Для цих гібридів прийнято використовувати назву, аналогічну видовій: їстівна жаба, Pelophylax esculentus (Linnaeus, 1758). Для P. esculentus характерне геміклональне успадкування, при якому продукувані ними гамети несуть або геном P. ridibundus, або геном P. lessonae, або обидва геноми. Для P. esculentus характерне спільне проживання з представниками батьківських видів у геміклональних популяційних системах, ГПС (Кравченко, 2013; Шабанов, 2015). Відомі також ГПС, що складаються виключно з P. esculentus.
Один із способів позначення типів ГПС пов’язаний із перерахуванням входять у їх склад форм жаб, при цьому літера L означає P. lessonae, R – P. ridibundus, а E – P. esculentus. Наявність у складі ГПС поліплоїдів P. esculentus позначається як Ep. У басейні р. Северський Донец на території Харківської області описано Північно‑Донецький центр різноманіття Pelophylax esculentus complex, що характеризується незвичним складом ГПС (Borkin et al., 2004; Боркін і др., 2005; Mezhzherin et al., 2010; Шабанов, 2015). Для нього характерні R‑E‑ГПС і R‑E‑Ep‑ГПС, а також популяції P. ridibundus, а статево зрілі представники P. lessonae повністю відсутні.
Раніше (Шабанов і др., 2014) була висунута гіпотеза, що різноманіття особин всередині однієї популяції або ГПС (у випадку зелених жаб) може бути описане як прояв внутрішньопопуляційних онтогенетичних стратегій, ВОС. Онтогенетична стратегія – це ієрархія пріоритетів, що проявляються у розвитку організму. Внутрішньопопуляційна стратегія – один із дискретних або об’єднаних у континуум варіантів реалізації видоспецифічної стратегії, що зустрічаються у представників однієї популяції або ГПС. ВОС характеризується певним синдромом (комплексом пов’язаних ознак), що відображає специфіку адаптації до середовища.
Метою даної роботи було з’ясування того, чи відображає внутрішньопопуляційне різноманіття представників Pelophylax esculentus complex з Харківської області різницю їх ВОС. Якщо це припущення вірне, на одних і тих самих етапах онтогенезу у одних особин пріоритет має підвищення їхньої власної життєздатності, а у інших – розмноження. Для досягнення мети роботи вирішувалися наступні завдання:
– визначити форми, до яких належать жаби, виміряти їх довжину, встановити за допомогою скелетохронології їх вік і ретроспективно визначити їх динаміку росту протягом останніх років життя;
– вивчити емпіричне різноманіття досліджених особин за розміром їх тіла у певному віці, як за провідною ознакою (тобто ознакою, яка може відображати ВОС);
– порівняти групи жаб, що відрізняються за провідною ознакою, за характеристиками, що відображають пріоритети в їх онтогенезі: швидкість росту, тривалість життя, плодючість самок, час початку участі у розмноженні;
– у випадку значних відмінностей між групами, виділеними за провідною ознакою, за характеристиками, пов’язаними з онтогенетичними пріоритетами, описати особливості ВОС, реалізованих представниками цих груп;
– встановити можливе значення різноманіття особин за їх ВОС для стійкості ГПС представників Pelophylax esculentus complex.
Матеріали та методи
Склад вивченої нами сукупності зелених жаб охарактеризовано в табл. 1. Усі жаби були зібрані в Харківській області, 550 – у Північно‑Донецькому центрі різноманіття Pelophylax esculentus complex (486 – у R‑E‑Ep‑ГПС і 64 – у R‑E‑ГПС), а 25 – у басейні Дніпра (7 – у R‑E‑ГПС і 18 – у L‑E‑R‑ГПС). Збір жаб проводився під час нересту в темний час доби.
Етапи обробки матеріалу детально описані у публікації, що стосується частини вивченого в даній роботі матеріалу (Усова, 2014). Належність особи до однієї з форм Pelophylax esculentus complex визначали за зовнішніми ознаками та розміром еритроцитів (Бондарева і др., 2012). Склад геномів ряду вивчених нами особин підтвердили за допомогою проточної ДНК‑цитометрії (Borkin et al., 2004). Проточну цитометрію проводили С. Н. Литвинчук і Ю. М. Розанов у ЦІН РАН (м. Санкт‑Петербург, РФ).
Таблиця 1.
Склад вивченої сукупності зелених жаб за формою (належність до батьківського виду, диплоїдним або триплоїдним гібрідам) та статтю
| Форма | Непзрілих, екз. | Самок, екз. | Самців, екз. | Всього, екз. |
|-------|----------------|------------|-------------|--------------|
| P. ridibundus | 13 | 149 | 31 | 193 |
| P. esculentus, 2n | 2 | 134 | 212 | 348 |
| P. esculentus, 3n | 2 | 12 | 20 | 34 |
| Всього | 17 | 295 | 263 | 575 |
У жаб видаляли другий (найдовший) палець задньої ноги. Його четверту фалангу очищали, декальцинували і отримували зрізи середньої частини діафізу (товщиною 20–22 мкм) на заморожуючому мікротомі. Зрізи фарбували гематоксиліном Ерліха, розміщували в гліцерині під покривним склом і фотографували під мікроскопом USB‑камерою. На електронних фотографіях у програмі Adobe Photoshop CS5 підраховували та вимірювали лінії склеювання. Результати вимірювань переводили в мікрометри, використовуючи встановлений за допомогою фотографування об’єкт‑мікрометра коефіцієнт перерахунку. Результати вимірювань збирали та обробляли у базі даних, створеній у програмі Statistica 8, StatSoft Inc.
При інтерпретації скелетохронологічних препаратів необхідно прийняти два рішення. По‑перше, треба визначити, які лінії склеювання утворилися під час зимувань, а які пов’язані зі зупинкою росту в теплий сезон. По‑друге, слід припустити, скільки ліній склеювання було резорбовано (зруйновано) через розширення кістковомозкової порожнини, розташованої всередині кістки. Після певного віку розширення кістковомозкової порожнини припиняється, і вона заповнюється ендостальною кістковою тканиною зі своїми лініями склеювання. Ці лінії важливо не плутати з лініями склеювання в периостальній частині кістки (Смирина, 1983).
Точно визначити, скільки ліній склеювання в периостальній частині кістки було резорбовано при розширенні кістковомозкової порожнини, у ряді випадків неможливо. Однак обґрунтоване припущення про кількість таких ліній можна зробити, порівнюючи препарати, отримані від різних індивідів з одного або близьких місць проживання. Підставами для прийняття гіпотези про кількість резорбованих ліній є поперечник межі між ендостальною та периостальною частиною кістки (чим він більший, тим більше ліній склеювання в периостальній кістці могло бути резорбовано); розмір першої повністю збереженої лінії склеювання (яка за розмірами може відповідати першій, другій або наступним лініям склеювання, що утворюються у жаб у досліджуваному місці); видимі на препараті залишки частково резорбованих ліній. Прийняті нами на цих підставах припущення про кількість резорбованих ліній у вивченої сукупності жаб відображені на рис. 1.
Визначення довжини тіла жаби під час минулих зимувань базується на припущенні, що P/L = li/Lj, де P – середній зовнішній поперечник кістки (її периостальної частини) у момент дослідження, L – довжина тіла у момент дослідження, li – середній поперечник лінії склеювання, утвореної під час певного зимування, Lj – довжина тіла, яку особина мала під час відповідного зимування. У такому випадку Lj = L × li/P (Усова, Шабанов, 2009). На підставі оцінок розміру тіла особини під час двох послідовних зимувань можна встановити відносний річний приріст довжини тіла gi = (Lj − Lj‑1)/Lj‑1.
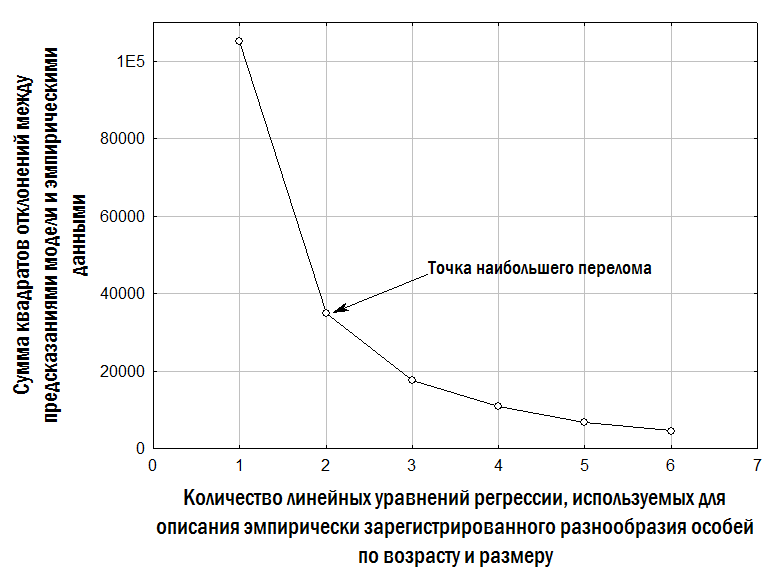
Пошук оптимальних значень коефіцієнтів для кожної з моделей, що апроксимують зафіксоване нами емпіричне різноманіття жаб, здійснювали за допомогою надбудови «Пошук рішення» в Microsoft Excel. Для цього обчислювали суму квадратів відхилень між спостережуваною та розрахованою (за кожною моделлю) довжиною тіла кожної особини. Потім надбудова «Пошук рішення» мінімізувала суму квадратів відхилень, змінюючи значення коефіцієнтів, підставлених у відповідні рівняння моделей.
Плодовитість самок визначали, отримуючи кладки ікри в штучних умовах. Кладки фотографували цифровим фотоапаратом, а потім підраховували ікринки за фотографією.
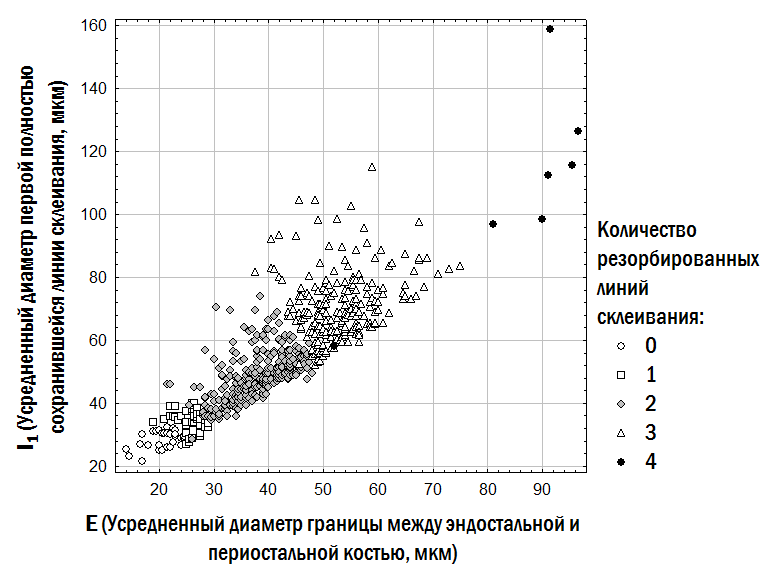
Рис. 1. Прийняті в даній роботі припущення про кількість резорбованих ліній склеювання в периостальній кістці у вивченої сукупності жаб
Вивчення стійкості ГПС Pelophylax esculentus complex проводили з використанням імітаційної моделі, розробленої згідно з поставленим нами завданням А. О. Леоновим за участі М. В. Владимирової, Г. Н. Жолткевича (ХНУ імені В. Н. Каразіна), J. Newman та Q. Mair (Glasgow Caledonian University). Ця модель є консольним застосунком для Java, який імітує зміни складу ГПС зелених жаб. На підставі заданих початкових параметрів модель покроково проводить перебудову описуваної модельної ГПС (рис. 2).
До початкових параметрів, що задаються при моделюванні, належать:
– початковий склад (чисельність особин різних форм і вікових груп) модельної ГПС;
– популяційно‑біологічні параметри всіх розглянутих у ній груп жаб (за генотипом і віком), що включають їх життєздатність (ймовірність виживання в неконкурентному середовищі), конкурентоспроможність (ймовірність збереження під час конкурентного виключення), ймовірність утворення пари з партнером під час розмноження, вік першого розмноження, максимальну тривалість життя, плодючість та потребу в ресурсах;
– варіанти всіх можливих схрещувань з вказанням ймовірностей появи різних генотипів у потомстві;
– місткість середовища (кількість доступних у ньому ресурсів);
– сценарії імміграції та еміграції (за потреби).
На кожному кроці роботи моделі доля кожної особини в моделюваній ГПС (її виживання та розмноження) визначається випадковими процесами і описується ймовірностями, заданими у початкових параметрах моделі. Для кожного набору початкових умов виконувалося 10 імітацій. Результати імітацій класифікували залежно від складу форм жаб, присутніх у модельній ГПС через 500 циклів (що відповідає 500 рокам), після чого визначали розподіл ймовірностей різних результатів імітацій залежно від початкового складу ГПС та прийнятих параметрів життєздатності.
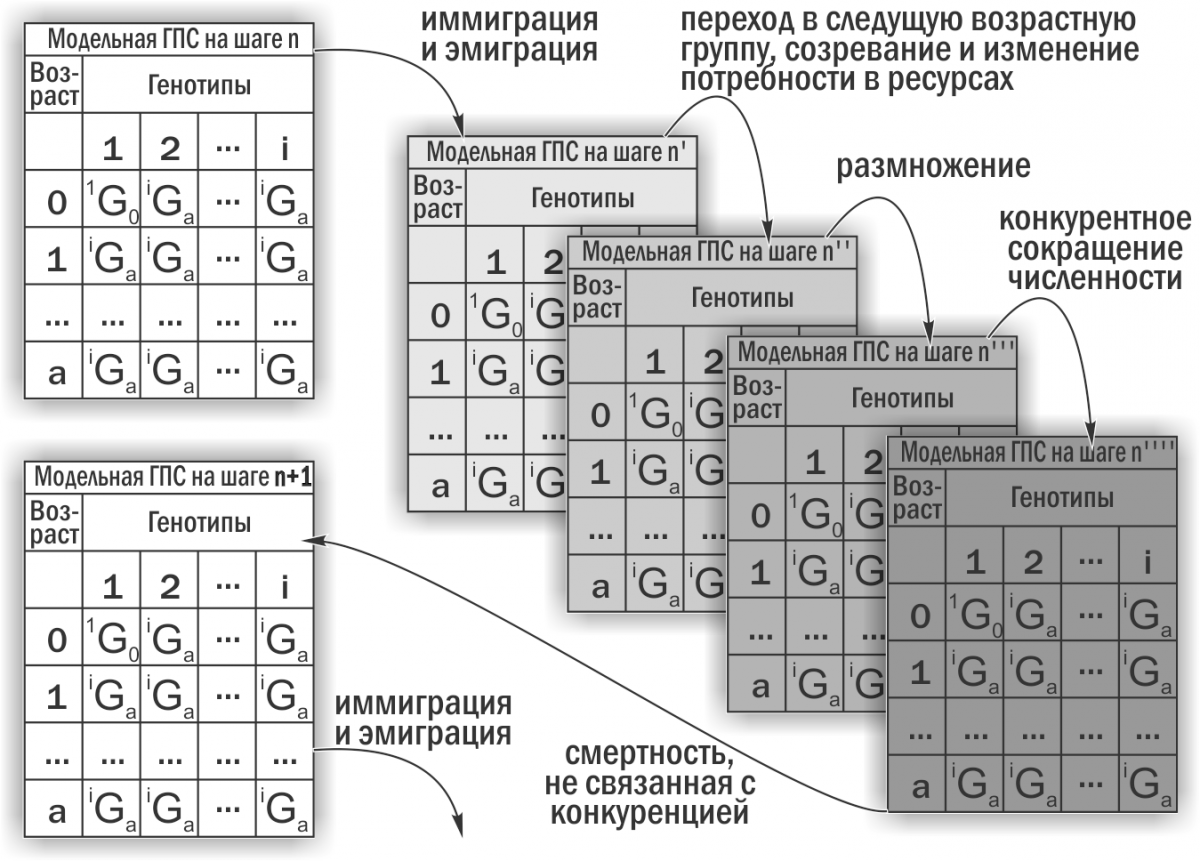
Рис. 2. Обчислення в кожному циклі роботи імітаційної моделі (Шабанов, 2015)
Результати та обговорення
1. Опис залежності розміру жаб від їх віку. Залежність розміру вивчених жаб від їх віку показана на рис. 3. Відображене на цьому рисунку різноманіття особин може бути описане різними моделями (табл. 2).
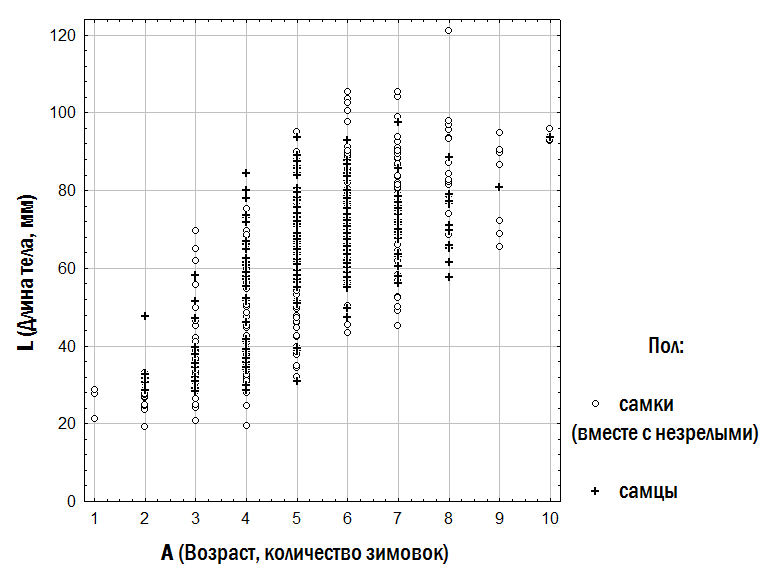
Рис. 3. Залежність розміру жаб від їх віку
Таблиця 2.
Результати апроксимації спостережуваних відповідностей віку (A) та довжини тіла (L) зелених жаб за допомогою трьох різних моделей
| Модель | Апроксимація | Сума квадратів відхилень |
|--------|--------------|--------------------------|
| Лінійна | L = a + b×A | L = 16,6 + 8,6×A | 105 087,5 |
| Квадратична | L = a + b×A + c×A² | L = –3,7 + 17,2×A – 0,8×A² | 99 351,1 |
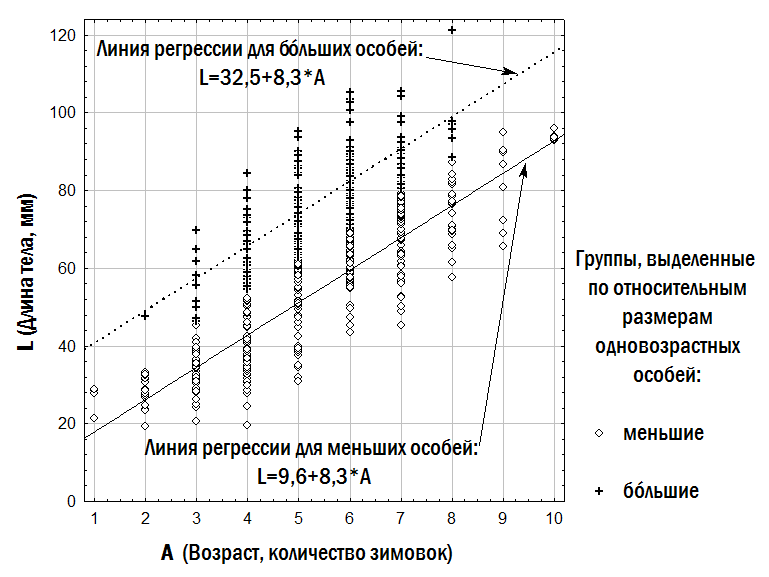
| Дві лінійні залежності для відносно менших і відносно більших одновозрастних особин | lL = la + lb×A; bL = ba + bb×A | lL = 9,7 + 8,3×A; bL = 32,5 + 8,3×A | 34 906,5 |
| Три лінійні залежності для відносно менших, проміжних і більших особин | lL = la + lb×A; mL = ma + mb×A; bL = ba + bb×A | lL = 3,7 + 8,4×A; mL = 14,6 + 9,1×A; bL = 32,9 + 9,1×A | 17 559,7{
"text": "Третя з використаних нами моделей передбачає, що різноманіття особин одного віку за їхнім розміром може бути описане за допомогою двох ліній регресії, одна з яких відповідає відносно меншим, а інша – відносно більшим одновіковим особинам. Позначення, використані в табл. 2, пов’язані з англійськими словами lesser – менший і bigger – більший. Під час апроксимації спостережуваного різноманіття віково-розмірних відповідностей жаб використаний нами алгоритм обчислював значення коефіцієнтів двох лінійних залежностей, мінімізуючи суму квадратів відстаней від кожної точки до найближчої до неї прямої. Ми задали обмеження, згідно з якими ba ≥ la та bb ≥ lb (позначення див. у табл. 2). Результат опису емпіричних даних за допомогою двох ліній регресії, що відповідають відносно меншим і відносно більшим одновіковим особинам, показано на рис. 4. Модель з трьома лініями регресії (табл. 2) побудована аналогічним чином; у ній додається лінія регресії, що відповідає проміжним (medium) за відносним розміром особинам.\n
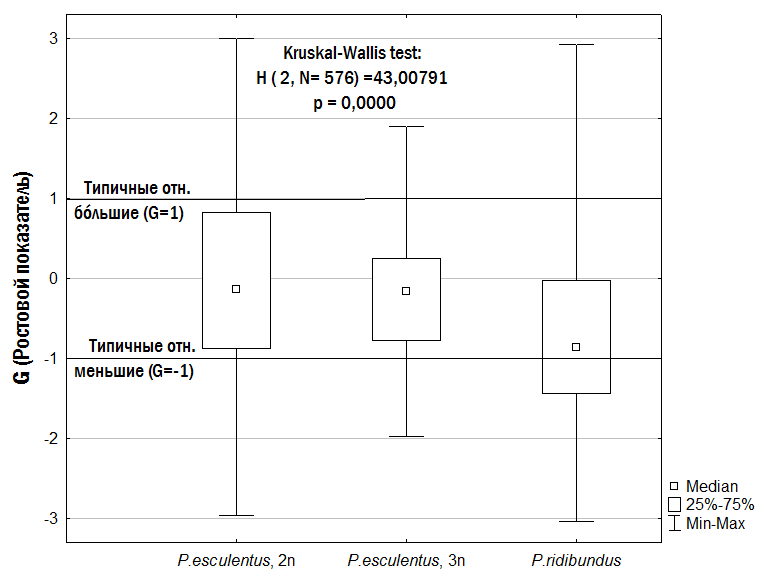
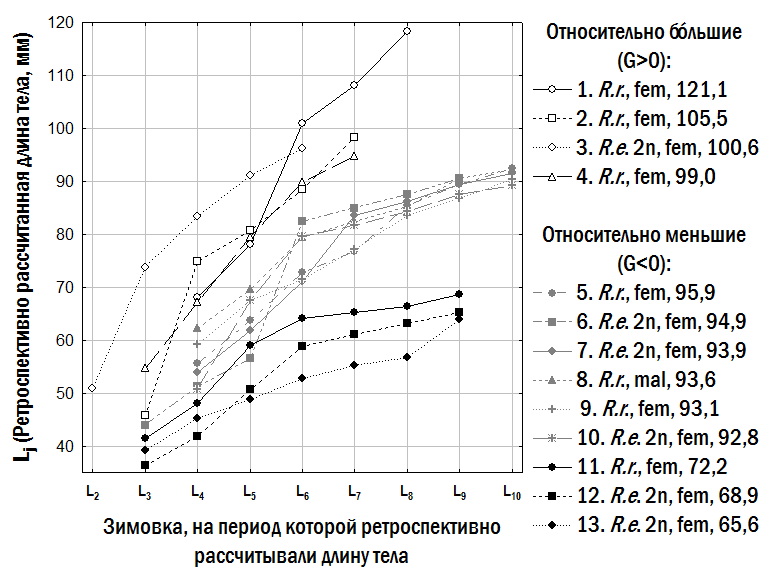
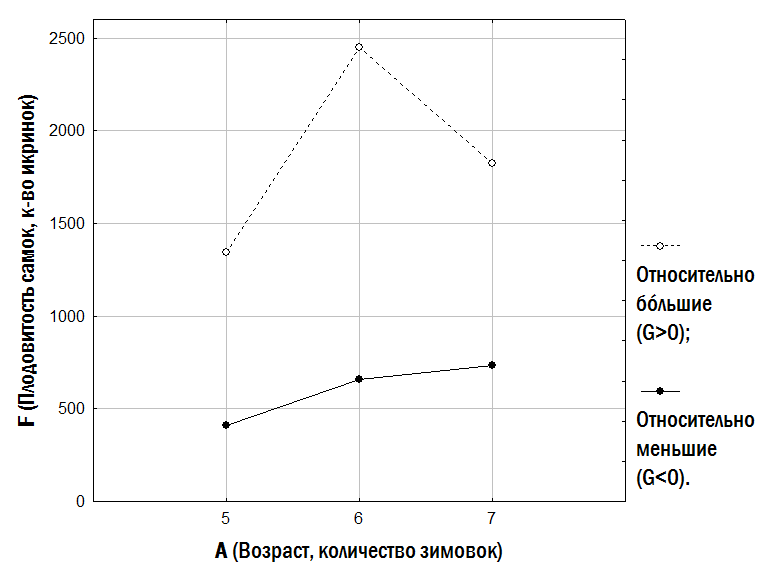
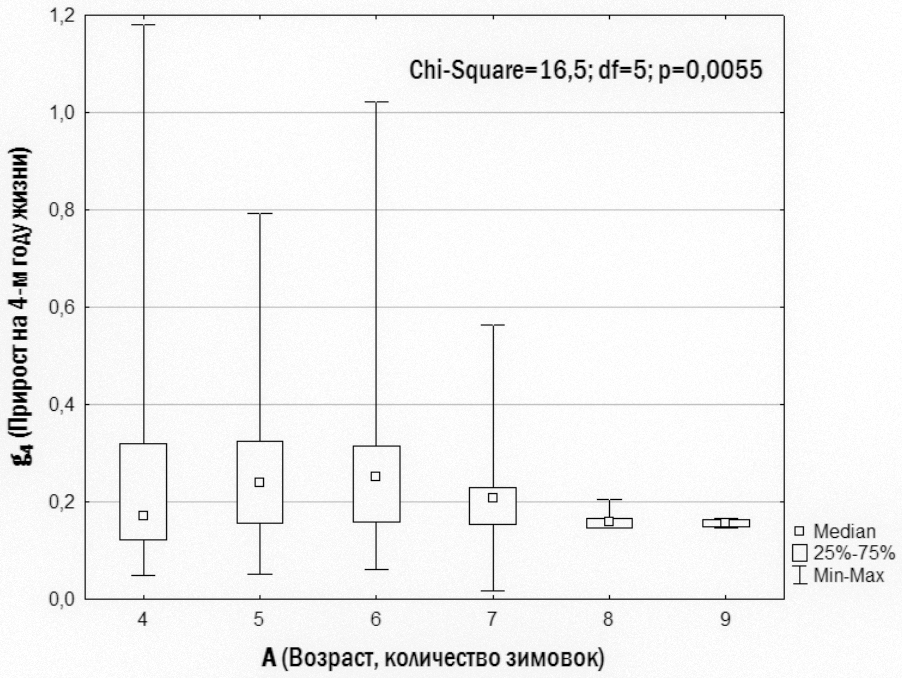
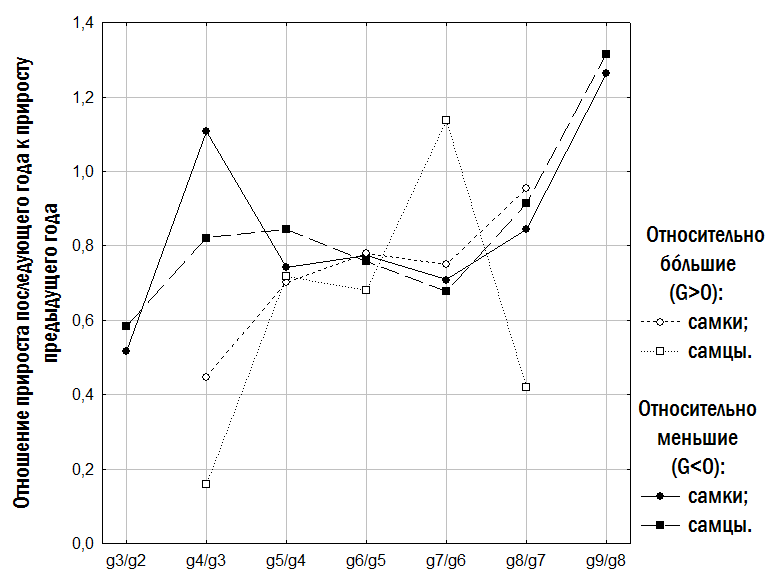
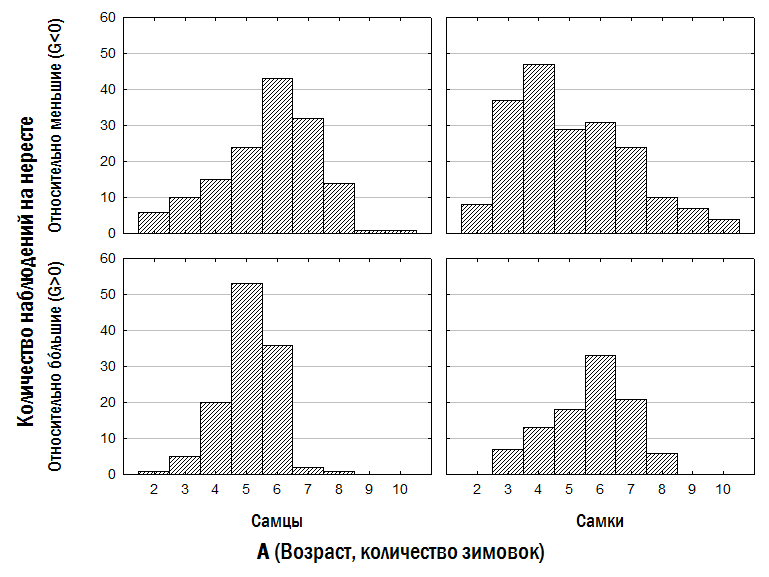
}Слід особливо підкреслити, що малорозмірні (G<0) і крупнорозмірні (G>0) особи зустрічаються в одних і тих самих місцевих середовищах і належать як до P. ridibundus, так і до диплоїдів і триплоїдів P. esculentus. Проте, як встановлено нами (рис. 6), у P. esculentus ВОС крупнорозмірності спостерігається значно частіше, ніж у P. ridibundus.
8. Можливе значення різноманітності ВОС, реалізованих представниками Pelophylax esculentus complex в одній ГПС. Оскільки ВОС малорозмірності і крупнорозмірності зустрічаються у зелених жаб, що спільно мешкають в одній ГПС, ми розглядаємо це різноманіття як адаптацію до внутріпопуляційних взаємодій. Для вивчення його значення може бути корисне імітаційне моделювання. Описаний нижче феномен був вперше зафіксований нами під час експериментів з імітаційною моделлю ГПС зелених жаб, виконаною в середовищі Microsoft Excel (Kravchenko, 2013). Потім його було детальніше досліджено нами з використанням описаної вище моделі А. О. Леонова (Shabanov et al., 2015).
Ми розглядали R‑E‑ГПС, що складаються з P. ridibundus і диплоїдних P. esculentus, які передають у гаметах жіночий геном P. lessonae. Для позначення геномів P. ridibundus ми використовуємо літеру R, а для позначення геномів P. lessonae — літеру L. Чоловічі геноми позначаються надстрочним знаком Y, а жіночі — X (гетерогаметним статтю у зелених жаб є чоловіча; Plötner, 2005). Клональний характер передачі геному позначається заключенням його символу в дужки. Використовуючи ці позначення, можна перерахувати всі типи схрещувань, які можуть відбуватися в описуваній ГПС:
– відтворення батьківського виду: ♀XRXR×♂XRYR → ♀♀XRXR : ♂♂XRYR;
– схрещування представників батьківського виду з гібридами, при якому весь нащадок складається з гібридів: ♀XRXR×♂YR(XL) → ♀♀XR(XL); ♀XR(XL)×♂XRYR → ♀♀XR(XL) : ♂♂YR(XL);
– схрещування гібридів з утворенням нежиттєздатних представників відсутнього в даній ГПС батьківського виду: ♀XR(XL)×♂YR(XL) → ♀♀(XL)(XL) → † (Plötner, 2005; Shabanov, 2015).
Трансформації описаної R‑E‑ГПС можуть призвести до одного з трьох результатів:
результат №1: перехід до популяції P. ridibundus; внаслідок зникнення P. esculentus;
результат №2: збереження R‑E‑ГПС внаслідок співіснування P. ridibundus і P. esculentus;
результат №3: зникнення ГПС внаслідок зникнення в ній P. ridibundus.
Якщо популяційно‑біологічні параметри P. ridibundus і P. esculentus, прийняті при моделюванні, ідентичні, частка P. esculentus у ГПС безперервно зростає, внаслідок їх більш ефективного відтворення. У такому випадку трансформації R‑E‑ГПС приводять до результату №3. Такий результат означає, що уявлення про ідентичність параметрів життєздатності у P. ridibundus і P. esculentus не відповідає дійсності. Якби це припущення було справедливим, R‑E‑ГПС існували б відносно недовго і закономірно зникали. Водночас нам відомо, що такі системи широко поширені в басейнах р. Мжа і р. Уди, правих притоках р. Северського Дону (Shabanov, 2015).
Ми припустили, що стійке співіснування P. ridibundus і P. esculentus може пояснюватися нижчою життєздатністю P. esculentus. Відповідно до цього припущення, зниження життєздатності гібридів має компенсувати перевагу в їх відтворенні. Як показано на рис. 12, це припущення не виправдало себе: залежно від співвідношення смертності P. ridibundus і P. esculentus реалізується або результат №1, або результат №3.
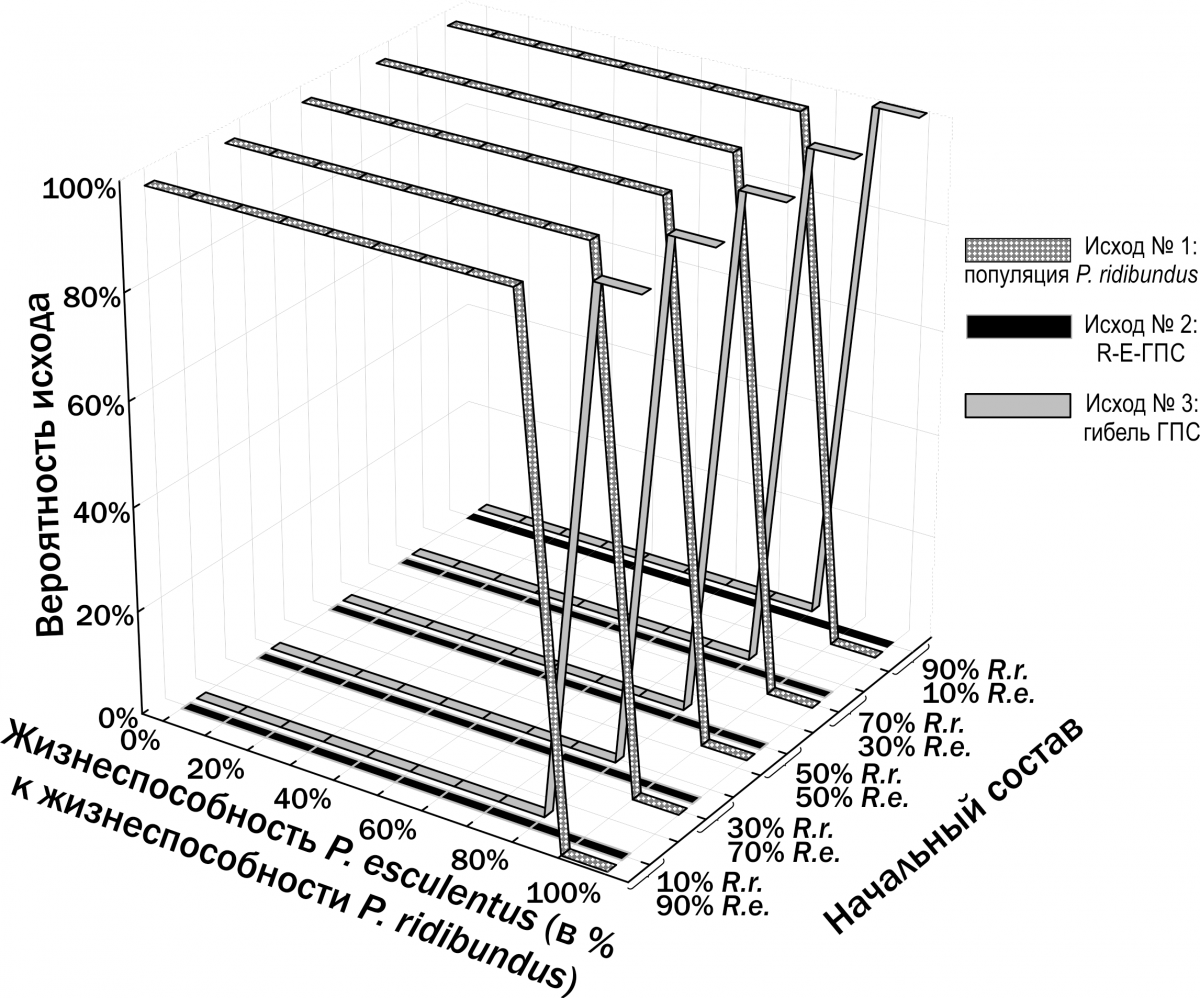
Рис. 12. Результати моделювання трансформацій R‑E‑ГПС при припущенні, що P. ridibundus і P. esculentus відрізняються лише за ймовірністю їхньої загибелі
Інший результат спостерігається у випадку, якщо P. ridibundus і P. esculentus реалізують різні ВОС. На рис. 13 показані результати моделювання у випадку, якщо P. ridibundus реалізує ВОС тугорослості, а P. esculentus — ВОС скороспілості (Shabanov et al., 2014). У цьому випадку виникає зона співвідношення життєздатностей двох форм, де можливе їх стале співіснування.
Ми припускаємо, що стале існування R‑E‑ГПС пов’язане з різноманіттям внутрі‑популяційних онтогенетичних стратегій особин у їх складі.
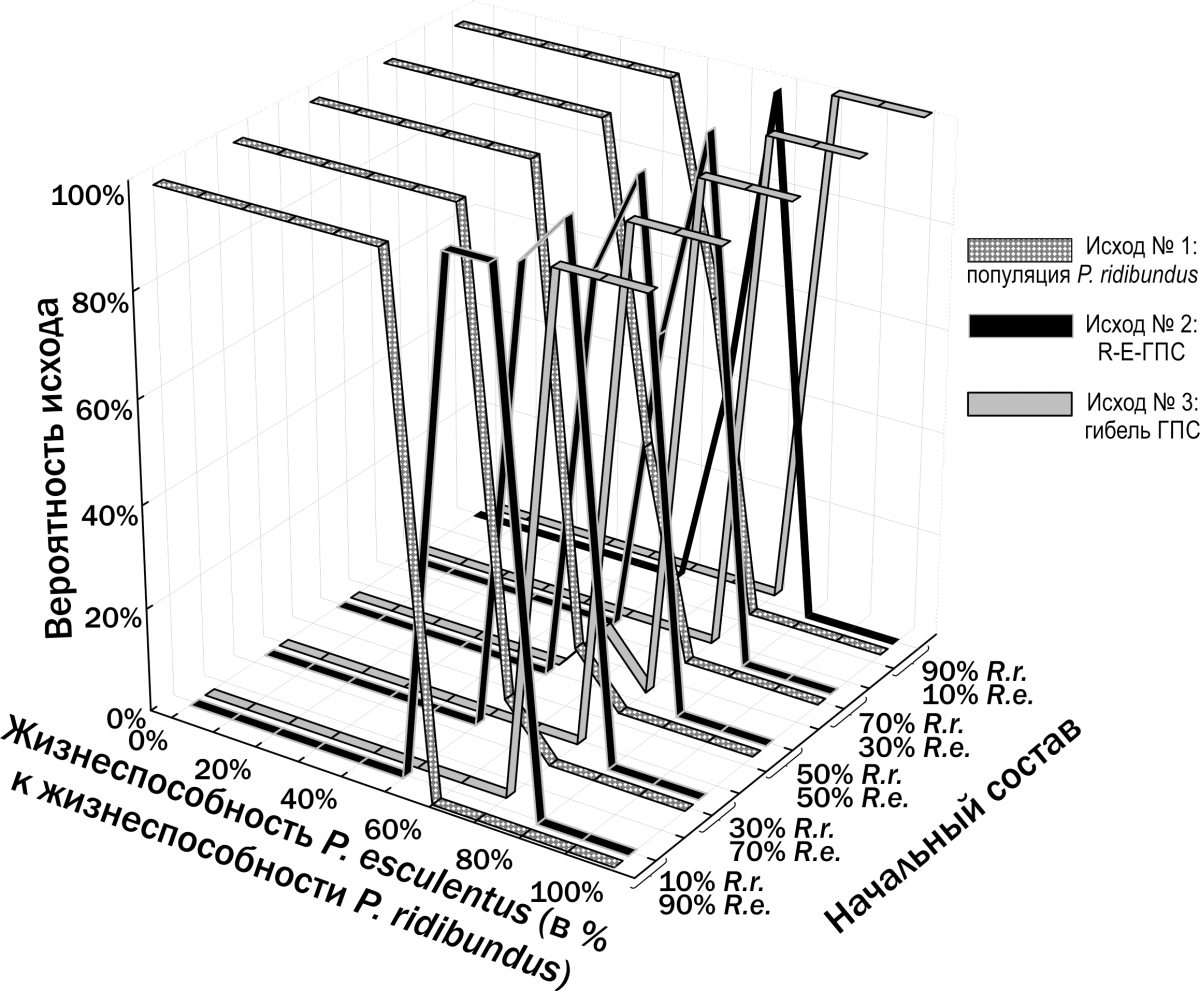
Рис. 13. Результати моделювання трансформацій R‑E‑ГПС при припущенні, що P. ridibundus і P. esculentus відрізняються за їх ВОС
Подяки
Автори висловлюють глибоку подяку класикові скелетохронологічних досліджень амфібій Е. М. Смирині (м. Москва) за допомогу в освоєнні методики, використаної в роботі, О. В. Біруку за кариогенетичні дослідження ряду вивчених нами особин жаб, членам групи популяційної екології амфібій Харківського національного університету імені В. Н. Каразіна А. В. Коршунову, Є. В. Мелешко та іншим за допомогу у зборі та обробці матеріалу, співробітникам ЦІН РАН (м. Санкт‑Петербург) С. Н. Литвинчуку і Ю. М. Розанову за визначення генотипів ряду вивчених особин з використанням проточної ДНК‑цитометрії, а також А. О. Леонову, М. В. Владимировій, Г. Н. Жолткевичу (ХНУ імені В. Н. Каразіна), J. Newman і Q. Mair (Glasgow Caledonian University) за допомогу в імітаційному моделюванні. Крім того, автори дякують А. А. Атемасову за конструктивну критику даної роботи.
Список літератури
Бондарева А. А., Бибик Ю. С., Самило С. М., Шабанов Д. А. Цитогенетичні особливості еритроцитів зелених жаб з Північно‑Донецького центру різноманіття Pelophylax esculentus complex // Вісник Харківського національного університету імені В. Н. Каразіна. Серія «Біологія». – 2012. – Вип. 15 (№ 1008). – С. 116–123. /Bondareva A.A., Bibik Yu.S., Samilo S.M., Shabanov D.A. Tsitogeneticheskiye osobennosti eritrotsitov zelenykh lyagushek iz Seversko-Donetskogo tsentra raznoobraziya Pelophylax esculentus complex // Vіsnyk Kharkіvs'kogo natsional'nogo unіversitetu іmenі V.N.Karazіna. Serіya «Bіologіya». – 2012. – Vyp.15 (№1008). – S. 116–123.
Боркин Л. Я., Зиненко А. І., Коршунов А. В. та ін. Масова поліплоїдія в гібридогенному комплексі Rana esculenta (Ranidae, Anura, Amphibia) на Сході України // Матеріали I конференції Українського Герпетологічного Товариства – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26. /Borkin L.Ya., Zinenko A.I., Korshunov A.V. i dr. Massovaya poliploidiya v gibridogennom komplekse Rana esculenta (Ranidae, Anura, Amphibia) na Vostoke Ukrainyi // MaterIali I konferentsiyi Ukrayinskogo Gerpetologichnogo Tovaristva – K.: Zoomuzey NNPM NANU, 2005. – S. 23–26.
Кравченко М. О. Екологічна стійкість популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex). Автореф. дис. … канд. біол. наук / 03.00.16 – екологія. – Дніпропетровськ, 2013. – 20 с. /Kravchenko M.O. Ekologіchna stіykіst' populyatsіynykh system gіbrydogennogo kompleksu zelenykh zhab (Pelophylax esculentus complex). Аvtoref. dys. … kand. bіol. nauk / 03.00.16 – ekologіya. – Dnіpropetrovs'k, 2013. – 20s.
Маро А. Н., Шабанова А. В., Шабанов Д. А. Чи можуть умови розвитку гусениць Bufo bufo визначати темпи постметаморфічного зростання і дозрівання жаб? // Вопроси герпетологии. Матеріали III з'їзду Герпетологічного товариства ім. А. М. Нікольського. – СПб, 2008. – С. 274–280. /Maro A.N., Shabanova A.V., Shabanov D.A. Mogut li usloviya razvitiya golovastikov Bufo bufo opredelyat' tempy postmetamorficheskogo rosta i sozrevaniya zhab? // Voprosy gerpetologii. Materialy III s'yezda Gerpetologicheskogo obshchestva im. A.M.Nikol'skogo. – SPb, 2008. – S. 274–280.
Смирина Е. М. Передсмертне визначення віку і ретроспективна оцінка розмірів тіла сірої жаби (Bufo bufo) // Зоол. журн. – 1983. – Т. 63, № 3. – С. 437–444. /Smirina E.M. Prizhiznennoye opredeleniye vozrasta i retrospektivnaya otsenka razmerov tela seroy zhaby (Bufo bufo) // Zool. zhurn. – 1983. – T.63, №3. – S. 437–444.
Усова Є. Є. Вік і швидкість зростання зелених жаб (Pelophylax esculentus complex) Нижнього Добрицького ставка (Зміївський район Харківської області) // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2014. – Вип. 20, № 1100. – С. 204–212. /Usova Ye.Ye. Vozrast i skorost' rosta zelenykh lyagushek (Pelophylax esculentus complex) Nizhnego Dobritskogo pruda (Zmiyevskoy rayon Khar'kovskoy oblasti) // Vіsnyk Kharkіvs'kogo natsional'nogo unіversytetu іmenі V.N. Karazіna. Serіya: bіologіya. – 2014. – Vyp.20, №1100. – S. 204–212.
Усова Є. Є., Шабанов Д. А. Про оптимізацію методики ретроспективної оцінки динаміки розмірів тіла представників Pelophylax esculentus complex (Amphibia, Ranidae) за допомогою скелетохронології // Zoocenosis-2009. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ: Ліра, 2009. – С. 278–280. /Usova Ye.Ye., Shabanov D.A. Ob optimizatsii metodiki retrospektivnoy otsenki dinamiki razmerov tela predstaviteley Pelophylax esculentus complex (Amphibia, Ranidae) pri pomoshchi skeletokhronologii // Zoocenosis-2009. Bіorіznomanіttya ta rol' tvaryn v ekosistemakh. – Dnіpropetrovs'k: Lira, 2009. – S. 278–280.
Халафян А. А. Statistica 6. Статистичний аналіз даних. – М.: ТОВ «Біном‑Пресс», 2007. – 512 с. /Khalafyan A.A. Statistica 6. Statisticheskiy analiz dannykh. – M.: OOO «Binom-Press», 2007. – 512s.
Шабанов Д. А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України. Автореф. дис. … д‑ра біол. наук / 03.00.16 – екологія. – Дніпропетровськ, 2015. – 36 с. /Shabanov D.A. Evolyutsіyna ekologіya populyatsіynykh system gіbrydogennogo kompleksu zelenykh zhab (Pelophylax esculentus complex) Lіvoberezhnogo lіsostepu Ukrayiny. Avtoref. dys. … d-ra bіol. nauk / 03.00.16 – ekologіya. – Dnіpropetrovs'k, 2015. – 36s.
Шабанов Д. А., Коршунов А. В., Кравченко М. А. та ін. Внутріпопуляційні онтогенетичні стратегії скороспілості і тугорослості: визначення на прикладі безхвостих амфібій // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2014. – Вип. 22, № 1126. – С. 115–124. /Shabanov D.A., Korshunov A.V., Kravchenko M.A. i dr. Vnutripopulyatsionnyye ontogeneticheskiye strategii skorospelosti i tugoroslosti: opredeleniye na primere beskhvostykh amfibiy // Vіsnyk Kharkіvs'kogo natsional'nogo unіversitetu іmenі V.N. Karazіna. Serіya: Bіologіya. – 2014. – Vyp.22, №1126. – S. 115–124.
Шабанов Д. А., Литвинчук С. Н. Зелені жаби: життя без правил чи особливий спосіб еволюції? // Природа. – 2010. – № 3. – С. 29–36. /Shabanov D.A., Litvinchuk S.N. Zelenyye lyagushki: zhizn' bez pravil ili osobyy sposob evolyutsii? // Priroda. – 2010. – №3. – S. 29–36.
Biriuk O., Usova O., Meleshko O., Shabanov D. Composition and characteristic of subadult water frogs sample (Pelophylax esculentus complex) // Book of abstracts of the 3rd International workshop‑conference: Research and conservation of European herpetofauna and its environment: Bombina bombina, Emys orbicularis, and Coronella austriaca. – Daugavpils, Latvia, 2015. – P. 8.
Borkin L.J., Korshunov A.V., Lada G.A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology. – 2004. – Vol. 11, No 3. – P. 194–213.
Plötner J. Die westpaläarktichen Wasserfrösche. – Bielefeld: Laurenti‑Verlag, 2005. – 161 s.
Mezhzherin S.V., Morozov‑Leonov S.Yu., Rostovskaya O.V. et al. The ploidy and genetic structure of hybrid populations of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol. 44, No 3. – P. 212–216.
Shabanov D., Usova O., Kravchenko M. et al. Sustainable coexistence of the parental species and hemiclonal interspecific hybrids is provided by the variety of ontogenetic strategies // Herpetological Facts Journal. – 2015. – Vol. 2. – P. 35–43.
Представлено: Т.Ю. Маркіна / Presented by: T.Yu.Markina
Рецензент: А. А. Атемасов / Reviewer: A.A.Atemasov
Подано до редакції / Received: 1.10.2015