
Usova, Kravchenko, Shabanov (2015) Intrapopulation ontogenetic strategies in water frogs
Usova E. E., Kravchenko M. A., Shabanov D. A. Intrapopulation ontogenetic strategies in water frogs (Pelophylax esculentus complex) // Bulletin of V. N. Karazin Kharkiv National University. Series: Biology. – 2015. &nd...
Usova E. E., Kravchenko M. A., Shabanov D. A. Intrapopulation ontogenetic strategies in water frogs (Pelophylax esculentus complex) // Bulletin of V. N. Karazin Kharkiv National University. Series: Biology. – 2015. – Issue 25. – P. 223-238. UDC: (57.022+57.032):591.84:591.158.1:597.851 Intrapopulation ontogenetic strategies in water frogs (Pelophylax esculentus complex) E.E.Usova, M.A.Kravchenko, D.A.Shabanov V.N. Karazin Kharkiv National University (Kharkiv, Ukraine) e.e.usova@gmail.com, marinakravchenko2106@gmail.com, d.a.shabanov@gmail.com Using skeletochronology, 575 representatives of Pelophylax esculentus complex (193 P. ridibundus, 348 diploid and 34 triploid P. esculentus) from Kharkiv Oblast were studied. The diversity of frogs by age and body length was described by approximation using the least squares method with two regression lines corresponding to relatively smaller and relatively larger individuals of the same age. These two groups of individuals were compared by population-biological parameters, and syndromes (complexes of related traits) were registered that allow considering them as manifestations of intrapopulation ontogenetic strategies (IOS) of undersized and oversized types. Undersized/oversized IOS is characterised by low/high growth rate, early/late onset of female spawning participation and late/early onset for males, high/low life expectancy, low/high female fecundity, and a large/small number of breeding seasons in which the individual participates. Using simulation modelling, it was shown that differences between P. ridibundus and P. esculentus in IOS can ensure their stable coexistence in the hemiclonal population system. Key words: Pelophylax esculentus complex, skeletochronology, strategy, undersized, oversized, simulation modelling, hemiclonal population systems. Intrapopulation ontogenetic strategies in water frogs (Pelophylax esculentus complex) O.E.Usova, M.O.Kravchenko, D.A.Shabanov Using skeletochronology, 575 representatives of Pelophylax esculentus complex (193 P. ridibundus, 348 diploid and 34 triploid P. esculentus) from Kharkiv Oblast were studied. The diversity of frogs by age and body length was described by approximation using the least squares method with two regression lines corresponding to relatively smaller and relatively larger individuals of the same age. The smaller and larger individuals were compared by population-biological parameters, and syndromes (complexes of related traits) were registered that allow considering them as manifestations of intrapopulation ontogenetic strategies (IOS) of undersized and oversized types. The IOS of undersized/oversized type is characterised by low/high growth rate, early/late onset of female spawning participation and late/early onset for males, high/low life expectancy, low/high female fecundity, and a large/small number of breeding seasons in which the individual participates. Using simulation modelling, it was shown that differences between P. ridibundus and P. esculentus in IOS can ensure their stable coexistence in the hemiclonal population system. Key words: Pelophylax esculentus complex, skeletochronology, strategy, undersized, oversized, simulation modelling, hemiclonal population systems. The water frogs' (Pelophylax esculentus complex) intrapopulation ontogenetic strategies O.E.Usova, M.O.Kravchenko, D.A.Shabanov 575 representatives of Pelophylax esculentus complex (193 P. ridibundus, 348 diploid and 34 triploid P. esculentus) from Kharkivska oblast have been studied using skeletochronology. With approximation by the LS method, frogs' diversity on age and body length has been characterized by two regression lines, which correspond to relatively smaller and bigger individuals of the same age. We compared these individuals by population-biological parameters and registered syndromes (complexes of related traits), allowing their consideration as manifeatation of intrapopulation ontogenetic strategies (IOS). IOS of undersized/oversized is characterized by relatively low/high growth rate, high/low life expectancy, low/high fertility of females and relatively large/small number of breeding seasons in which the individual takes part. Participation in spawning begins early/late in undersized/oversized females and vice versa in males.
By using simulation it has been shown that differences in IOS between P. ridibundus and P. esculentus can provide their sustainable coexistence in the hemiclonal population system.
Key words: Pelophylax esculentus complex, skeletochronology, strategy, undersized, oversized, simulation, hemiclonal population systems. Introduction The hybridogenous complex of water frogs, Pelophylax esculentus complex (=Rana esculenta complex), consists of two parental species: the pool frog, Pelophylax lessonae (Camerano, 1882), and the marsh frog, Pelophylax ridibundus (Pallas, 1771), as well as their interspecific hybrids of various ploidy levels (Plötner, 2005; Shabanov, Litvinchuk, 2010). The name analogous to a species name is conventionally used for these hybrids: the edible frog, Pelophylax esculentus (Linnaeus, 1758). P. esculentus is characterised by hemiclonal inheritance, in which the gametes they produce carry either the genome of P. ridibundus, or the genome of P. lessonae, or both genomes. P. esculentus is characterised by cohabitation with representatives of the parental species in hemiclonal population systems, HPS (Kravchenko, 2013; Shabanov, 2015). HPS consisting exclusively of P. esculentus are also known.
One method of designating HPS types involves listing the frog forms composing them, with the letter L denoting P. lessonae, R denoting P. ridibundus, and E denoting P. esculentus. The presence of polyploid P. esculentus in an HPS is denoted as Ep. In the basin of the Seversky Donets River within Kharkiv Oblast, the Seversky-Donets Diversity Centre of Pelophylax esculentus complex has been described, characterised by an unusual HPS composition (Borkin et al., 2004; Borkin et al., 2005; Mezhzherin et al., 2010; Shabanov, 2015). It is characterised by R-E-HPS and R-E-Ep-HPS, as well as P. ridibundus populations, while sexually mature P. lessonae individuals are entirely absent.
Previously (Shabanov et al., 2014), a hypothesis was proposed that the diversity of individuals within a single population or HPS (in the case of water frogs) can be described as a manifestation of intrapopulation ontogenetic strategies, IOS. An ontogenetic strategy is a hierarchy of priorities manifesting in the development of an organism.
An intrapopulation strategy is one of the discrete or continuum-combined variants of the species-specific strategy found in representatives of a single population or HPS. IOS is characterised by a specific syndrome (complex of related traits) reflecting the specifics of adaptation to the environment.
The aim of this study was to determine whether the intrapopulation diversity of Pelophylax esculentus complex representatives from Kharkiv Oblast reflects differences in their IOS. If this assumption is correct, at the same ontogenetic stages, some individuals prioritise enhancement of their own viability, while others prioritise reproduction. To achieve the aim of the study, the following tasks were addressed: – to identify the forms to which the frogs belong, to measure their length, to establish their age using skeletochronology, and to retrospectively determine their growth dynamics over recent years of life; – to study the empirical diversity of the examined individuals by body size at a given age, as the leading trait (i.e., the trait that may reflect IOS); – to compare groups of frogs differing in the leading trait by characteristics reflecting priorities in their ontogenesis: growth rate, life expectancy, female fecundity, and age at onset of reproduction; – in the case of significant differences between groups distinguished by the leading trait in characteristics related to ontogenetic priorities, to describe the IOS features realised by representatives of these groups; – to establish the possible significance of individual IOS diversity for the stability of HPS of Pelophylax esculentus complex representatives. Material and Methods The composition of the studied sample of water frogs is characterised in Table 1.
All frogs were collected in Kharkiv Oblast; 550 from the Seversky-Donets Diversity Centre of Pelophylax esculentus complex (486 from R-E-Ep-HPS and 64 from R-E-HPS), and 25 from the Dnieper basin (7 from R-E-HPS and 18 from L-E-R-HPS). Frogs were collected during the spawning period at night. The stages of material processing are described in detail in a publication concerning part of the material studied in the present work (Usova, 2014). Assignment of individuals to one of the forms of Pelophylax esculentus complex was determined by external characteristics and erythrocyte size (Bondareva et al., 2012).
The genomic composition of several studied individuals was confirmed using flow DNA cytometry (Borkin et al., 2004). Flow cytometry was performed by S.N. Litvinchuk and Yu.M. Rozanov at the Cytology Institute of the Russian Academy of Sciences (St.
Petersburg, Russia). Table 1. Composition of the studied sample of water frogs by form (membership in parental species, diploid or triploid hybrids) and sex 263 Form Immature, specimens Females, specimens Males, specimens Total, specimens P. ridibundus 13 149 31 193 P. esculentus, 2n 2 134 212 348 P. esculentus, 3n 2 12 20 34 Total 17 295 263 575 The 2nd (longest) toe of the hind foot was removed from frogs. Its 4th phalanx was cleaned, decalcified, and cross-sections of the mid-diaphysis (20–22 µm thick) were obtained on a freezing microtome.
Sections were stained with Ehrlich's haematoxylin, mounted in glycerol under a coverslip, and photographed under a microscope with a USB camera. On digital photographs in Adobe Photoshop CS5, lines of arrest of growth (LAGs) were counted and measured. Measurement results were converted to micrometres using a conversion factor established by photographing a stage micrometre.
Results were collected and processed in a database created in Statistica 8, StatSoft Inc.
When interpreting skeletochronological preparations, two decisions must be made. First, it is necessary to determine which LAGs formed during hibernation periods and which are related to growth arrest during the warm season. Second, an assumption must be made as to how many LAGs were resorbed (destroyed) due to expansion of the medullary cavity located inside the bone. After a certain age, expansion of the medullary cavity ceases and it fills with endosteal bone tissue with its own LAGs.
It is important not to confuse these LAGs with those in the periosteal part of the bone (Smirina, 1983). It is sometimes impossible to determine precisely how many LAGs in the periosteal part of the bone were resorbed during expansion of the medullary cavity.
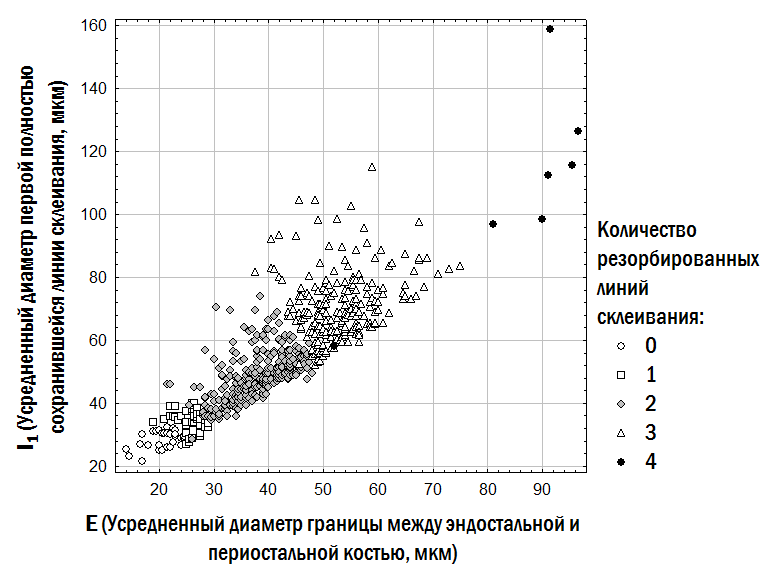
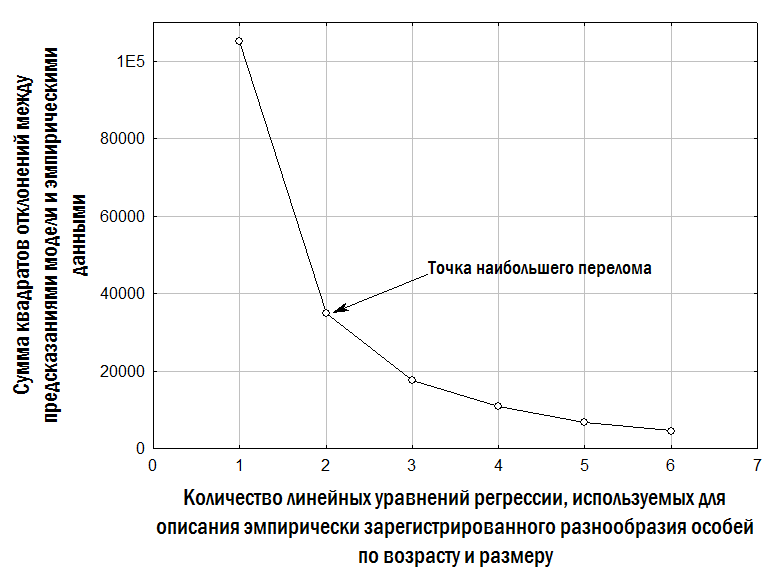
However, a reasoned assumption about the number of such lines can be made by comparing preparations obtained from different individuals from the same or closely related habitats. The basis for accepting a hypothesis about the number of resorbed LAGs includes: the diameter of the boundary between the endosteal and periosteal parts of the bone (the larger it is, the more LAGs in the periosteal bone may have been resorbed); the size of the first fully preserved LAG (which by its dimensions may correspond to the first, second, or subsequent LAGs formed in frogs at the studied locality); and visible remnants of partially resorbed LAGs on the preparation. The assumptions adopted on these grounds regarding the number of resorbed LAGs in the studied sample are reflected in Fig. 1. Calculation of frog body length during past hibernations is based on the assumption that P/L=li/Lj, where P is the averaged outer diameter of the bone (its periosteal part) at the time of study, L is the body length at the time of study, li is the averaged diameter of the LAG formed at a given hibernation, and Lj is the body length the individual had during the corresponding hibernation. In this case, Lj=L×li/P (Usova, Shabanov, 2009). Based on estimates of the individual's body size during two consecutive hibernations, the relative annual body length increment can be established: gi=(Lj-Lj-1)/Lj-1. The search for optimal coefficient values for each of the models approximating the empirical diversity of frogs we recorded was carried out using the Solver add-in in Microsoft Excel. For this purpose, the sum of squared deviations between the observed and calculated (according to each model) body length of each individual was computed. The Solver add-in then minimised the sum of squared deviations by changing the values of the coefficients substituted into the equations corresponding to the models used. Female fecundity was determined by obtaining clutches in artificial conditions. Clutches were photographed with a digital camera, and eggs were then counted from the photographs. Fig. 1. Assumptions adopted in this study regarding the number of resorbed LAGs in the periosteal bone for the studied sample of frogs The study of HPS stability in Pelophylax esculentus complex was conducted using a simulation model developed according to our specifications by A.O. Leonov with the participation of M.V. Vladimirova, G.N. Zholtkevich (V.N. Karazin Kharkiv National University), J. Newman and Q. Mair (Glasgow Caledonian University). This model is a console application for Java that simulates changes in the composition of the HPS of water frogs.
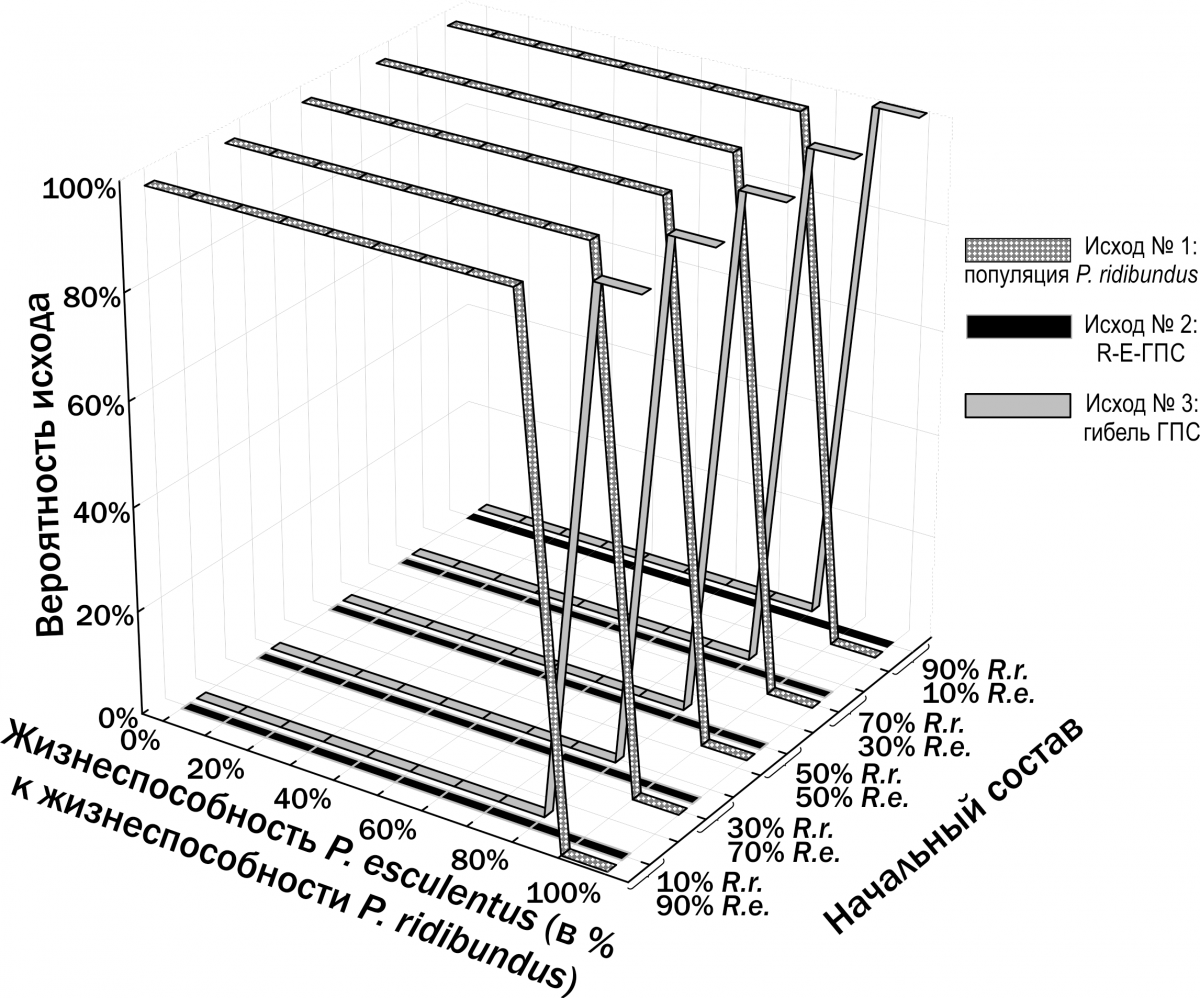
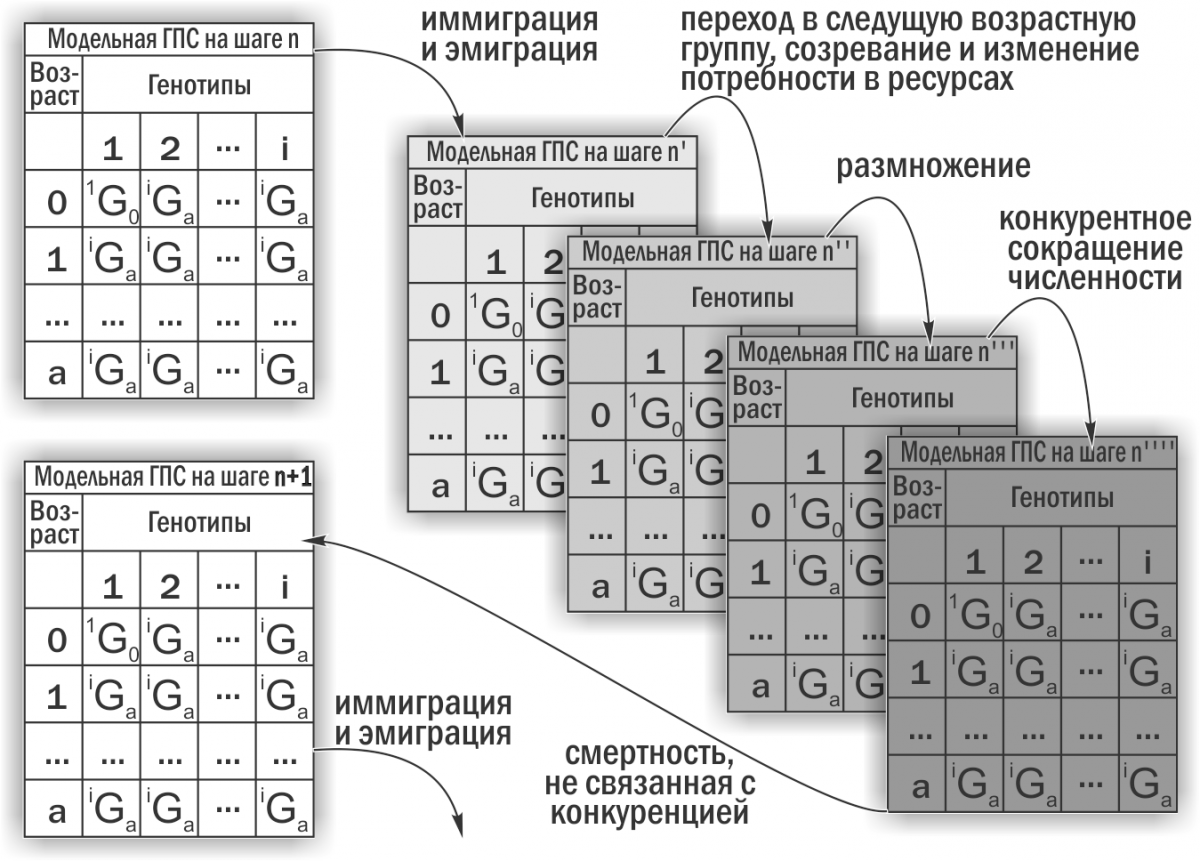
Based on specified initial parameters, the model performs step-by-step restructuring of the described model HPS (Fig.
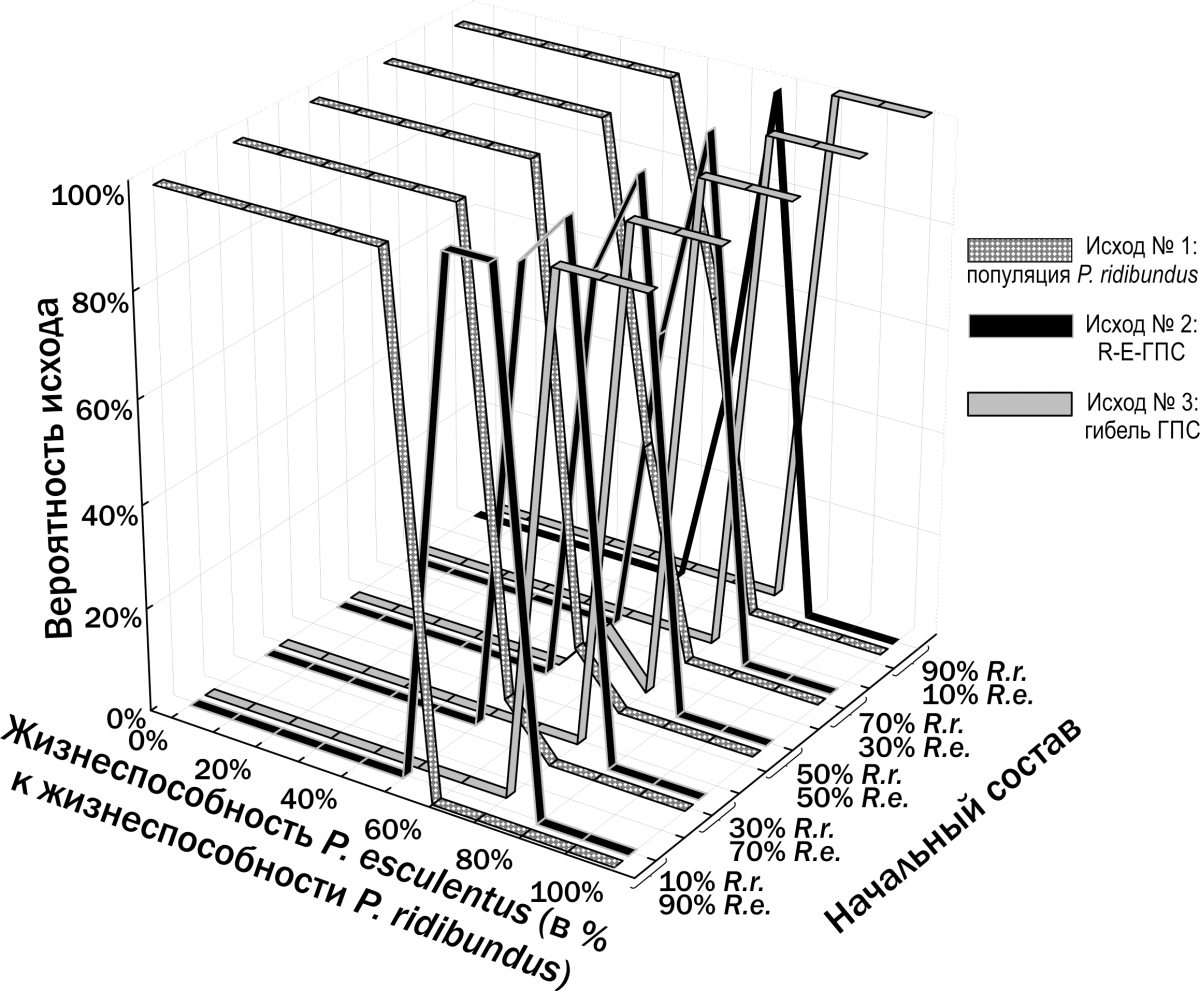
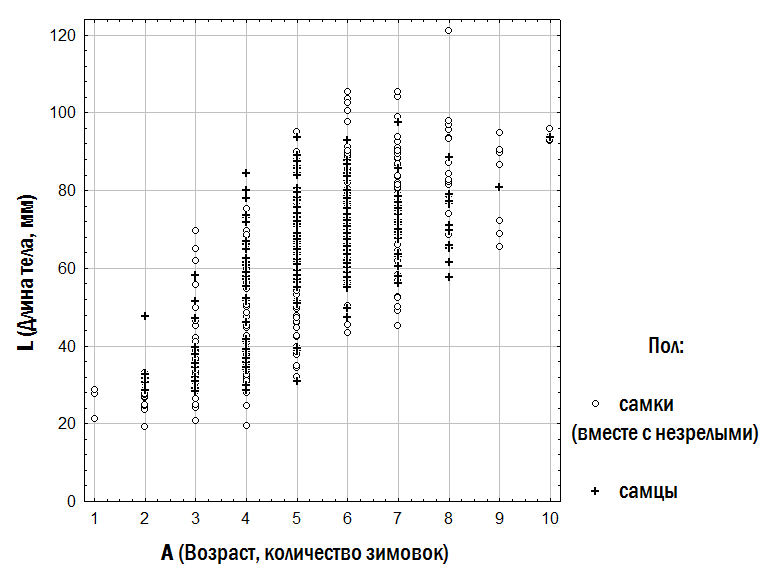
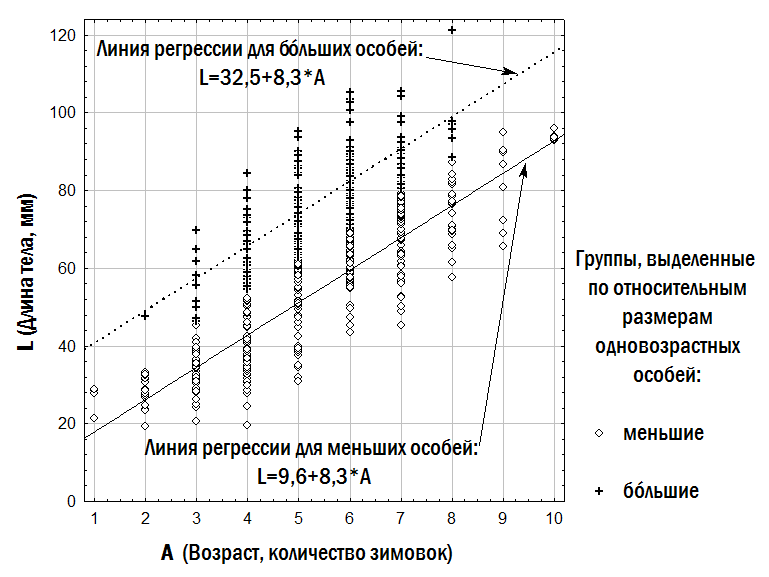
2). Initial parameters specified during modelling include: – initial composition (numbers of individuals of different forms and ages) of the model HPS; – population-biological parameters of all groups of frogs considered (differing in genotype and age), including viability (probability of survival in a non-competitive environment), competitive ability (probability of persistence during competitive exclusion), probability of pair formation with a partner during reproduction, age at first reproduction, maximum lifespan, fecundity, and resource requirements; – variants of all possible crosses with specified probabilities of different genotype frequencies in offspring; – carrying capacity (quantity of resources available in the environment); – immigration and emigration scenarios (when required). At each step of the model, the fate of each individual in the modelled HPS (its survival and reproduction) is determined by stochastic processes and is described by probabilities specified in the initial parameters of the model. For each set of initial conditions, 10 simulations were performed. The outcomes of simulations were classified according to the composition of frog forms present in the model HPS after 500 cycles (corresponding to 500 years), after which the probability distribution of different simulation outcomes was determined as a function of the initial HPS composition and the adopted viability parameters. Fig. 2. Computations in each cycle of the simulation model (Shabanov, 2015) Results and Discussion 1. Description of the dependence of frog size on age. The dependence of the size of studied frogs on their age is shown in Fig. 3. The individual diversity reflected in this figure can be described using different models (Table 2). Fig. 3. Dependence of frog size on age Table 2. Results of approximating the observed age (A) and body length (L) correspondences of water frogs using three different models | Model | Approximation | Sum of Squared Deviations | |--------|--------------|--------------------------| | Linear | L = a + b×A | L = 16.6 + 8.6×A | 105,087.5 | | Quadratic | L = a + b×A + c×A² | L = –3.7 + 17.2×A – 0.8×A² | 99,351.1 | | Two linear dependencies for relatively smaller and relatively larger individuals of the same age | lL = la + lb×A; bL = ba + bb×A | lL = 9.7 + 8.3×A; bL = 32.5 + 8.3×A | 34,906.5 | | Three linear dependencies for relatively smaller, intermediate, and larger individuals | lL = la + lb×A; mL = ma + mb×A; bL = ba + bb×A | lL = 3.7 + 8.4×A; mL = 14.6 + 9.1×A; bL = 32.9 + 9.1×A | 17,559.7