Вегерина А. О., Мелешко Е. В., Пырина И. С., Сапожникова В. А., Бирюк О. В. Определение соотношения диплоидов и триплоидов среди метаморфов зеленых лягушек в Cеверско-Донецком центре разнообразия Pelophylax esculentus complex // Вісник Харківського національного університету імені В.Н.Каразіна. Серія: біологія. – 2013. – Вип.18, №1079. – С. 107–113.
УДК: 597.851
Определение соотношения диплоидов и триплоидов среди метаморфов зеленых лягушек в Cеверско-Донецком центре разнообразия Pelophylax esculentus complex
А.О.Вегерина, Е.В.Мелешко, И.С.Пырина, В.А.Сапожникова, О.В.Бирюк
Харьковский национальный университет имени В.Н.Каразина (Харьков, Украина)
mykhailova.o.v@gmail.com
Проверена гипотеза, что доля триплоидов среди метаморфов представителей Pelophylax esculentus complex в Северско-Донецком центре разнообразия зеленых лягушек является более высокой, чем среди половозрелых особей. Определена плоидность 30 метаморфов, собранных в июле 2013 г. Установлено, что значение средней длины эритроцитов у метаморфов ниже таковой у половозрелых лягушек. Плоидность определялась путем подсчета количества хромосом в метафазных пластинках, полученных из эндотелия кишечника. Для определения плоидности в случае отсутствия делящихся клеток применили Ag-окрашивание, которое позволяет выявить ядрышки в интерфазных ядрах. Все изученные особи оказались диплоидными. С учетом выборки, изученной в 2012 г., один триплоид был зарегистрирован в объединенной выборке из 47 молодых особей; проверяемая гипотеза не подтвердилась. Выдвинуты предположения о причинах низкой доли триплоидов в исследованной выборке.
Ключевые слова: Pelophylax esculentus, триплоиды, эритроциты, Ag-окрашивание, плоидность.
Визначення співвідношення диплоїдів і триплоїдів серед метаморфів зелених жаб в Cіверсько-Донецькому центрі різноманіття Pelophylax esculentus complex
А.О.Вегеріна, О.В.Мелешко, І.С.Пиріна, В.О.Сапожникова, О.В.Бірюк
Харківський національний університет імені В.Н.Каразіна (Харків, Україна)
mykhailova.o.v@gmail.com
Перевірено гіпотезу, що частка триплоїдів серед метаморфів представників Pelophylax esculentus complex в Сіверсько-Донецькому центрі різноманіття зелених жаб є вищою, ніж серед статевозрілих особин. Визначено плоїдність 30 метаморфів, зібраних у липні 2013 р. Встановлено, що значення середньої довжини еритроцитів у метаморфів нижче такої у статевозрілих жаб. Плоїдність визначалася шляхом підрахунку кількості хромосом в метафазних пластинках, отриманих з ендотелію кишечника. Для визначення плоїдності у разі відсутності клітин, що діляться, застосували Ag-фарбування, яке дозволяє виявити ядерця в інтерфазних ядрах. Всі вивчені особини виявилися диплоїдними. З урахуванням вибірки, що була вивчена у 2012 р., один триплоїд був зареєстрований в об'єднаній вибірці з 47 молодих особин; гіпотеза, що перевірялася, не підтвердилася. Висунуто припущення про причини низької частки триплоїдів у дослідженій вибірці.
Ключові слова: Pelophylax esculentus, триплоіды, еритроцити, Ag-фарбування, плоідність.
The determination of the ratio of diploid and triploid green frogs underyearlings from the Pelophylax esculentus complex Seversky Donets diversity center
A.O.Vegerina, O.V.Meleshko, I.S.Pyrina, V.O.Sapoznikova, O.V.Biryuk
The hypothesis has been tested that the percentage of triploids among the metamorphs of Pelophylax esculentus complex representatives in the Seversky Donets green frogs diversity center is higher than that among the mature individuals. The ploidy of 30 metamorphs collected in July, 2013 was defined. It was found that the metamorphs erythrocytes mean length is lower than that in the mature frogs. Ploidy was defined by counting chromosomes in metaphase plates derived from intestine endothelium. To determine the ploidy in the absence of dividing cells the Ag-staining was used, which reveals the nucleoli in interphase nuclei. All the studied specimens were diploid. Given the sample studied in 2012, one triploid was registered in the combined sample of 47 young individuals; the tested hypothesis was not confirmed. The assumptions about the causes of the low percentage of triploids in the studied sample were made.
Key words: Pelophylax esculentus, triploids, red blood cells, Ag-staining, ploidy.
Введение
В состав гибридогенного комплекса зеленых лягушек (Pelophylax esculentus complex) входят два родительских вида: прудовая лягушка Pelophylax lessonae (Camerano, 1882) и озерная лягушка Pelophylax ridibundus (Pallas, 1771), от скрещивания которых образуются межвидовые гибриды. Для обозначения этих гибридов (с определенной степенью условности) используется название, аналогичное видовому – съедобная лягушка Pelophylax esculentus (Linnaeus, 1758). Различные формы гибридов отличаются по плоидности и характеру их гаметогенеза; в типичном случае они гемиклонально (без рекомбинации) передают в гаметы один из родительских геномов (Plötner, 2005). Особи родительских видов обитают и воспроизводятся с гибридами совместно, образуя гемиклональные популяционные системы. Расположенный в бассейне р. Северский Донец регион, характеризующийся высоким разнообразием гибридных зеленых лягушек, в том числе – распространением триплоидных гибридов, был назван Северско-Донецким центром разнообразия P. esculentus complex (Шабанов та ін., 2009).
В первой работе, в которой было сообщено об обнаружении триплоидных гибридов зеленых лягушек на этой территории, зарегистрированная доля триплоидов среди гибридов была относительно высокой и составляла 30% (Borkin et al., 2004). Изученная в этой работе выборка зеленых лягушек включала значительное количество сеголетков. В ряде иных работ, где изучались половозрелые особи, регистрировалась более низкая доля триплоидов в общем количестве гибридов: 9% (Mezhzherin et al., 2010) или 11% (Козак и др., 2012).
Описанное различие в результатах позволяет выдвинуть гипотезу о том, что доля триплоидов среди P. esculentus в Северско-Донецком центре разнообразия зеленых лягушек снижается с возрастом. В пользу этой гипотезы свидетельствует то, что триплоидные гибриды не являются отдельным таксоном и, вероятно, не имеют самостоятельной, независимой от диплоидов генеалогии. Геномы таких лягушек являются результатом эволюции диплоидных представителей родительских видов, и их триплоидность можно рассматривать как цитогенетическую аномалию. Основываясь на этом, можно предположить, что отбор уменьшает долю триплоидов в старших возрастных классах по сравнению с младшими.
В 2012 г. часть авторов данной работы исследовали выборку из 17 сеголетков и неполовозрелых особей, собранных в окрестностях с. Гайдары Змиевского района Харьковской области. Плоидность лягушек в этой работе определяли с помощью метода давленых препаратов (Михайлова и др., 2011). Зарегистрированная в этой работе доля триплоидов оказалась неожиданно низкой: из 17 особей 1 оказалась триплоидной и 16 – диплоидными.
В данной работе мы проверяли то же предположение, используя большую по численности выборку и сочетая косвенный метод определения плоидности с двумя прямыми. Косвенный метод определения плоидности состоит в измерении среднего размера эритроцитов каждой особи. Как известно, ядра клеток триплоидных гибридов содержат в полтора раза больше хроматина (Plötner, 2005); это приводит к увеличению размеров ядер и клеток в целом. Граница между размерами диплоидных и триплоидных эритроцитов в разных регионах обитания триплоидных P. esculentus может несколько отличаться. Ранее в ходе исследований размеров эритроцитов у полувзрослых и половозрелых зеленых лягушек из Северско-Донецкого центра их разнообразия было установлено, что граничная длина эритроцитов, разделяющая диплоидов и триплоидов, составляет 26–28 μм (Бондарева и др., 2012). Кроме того, было показано, что размер эритроцитов у неполовозрелых лягушек может зависеть от длины тела (Искендерова и др., 2012), а длина эритроцитов у неполовозрелых триплоидов может не достигать граничного размера, установленного для более крупных особей (Темников и др., 2012). Прямыми методами определения плоидности были кариоанализ и исследование ядрышковых организаторов в интерфазных ядрах (см. далее).
Материалы и методы
Для проверки высказанных гипотез нами была исследована выборка из 40 метаморфов (сеголетков, недавно завершивших метаморфоз) зеленых лягушек, отловленная в пойме р. Северский Донец в окрестностях с. Гайдары Змиевского района Харьковской области в конце июля 2013 года. Выборка состояла из особей с длиной тела от 18,5 до 33,3 мм. У отловленных метаморфов был взят мазок крови. 8 особей по техническим причинам исследованы не были. У оставшихся лягушек взяли фрагменты кишечника для кариологических исследований, но у 2 особей материал оказался непригодным для анализа, вследствие чего они также были исключены из дальнейшего исследования.
Получение и исследование мазков крови проводили в соответствии с опубликованной методикой (Бондарева и др., 2012).
Для кариоанализа исследуемым метаморфам делали инъекцию 0,04% раствора колхицина внутрибрюшинно (по 0,04–0,08 мл на особь в зависимости от веса). Через 24 часа животных усыпляли этилацетатом. В ходе вскрытия извлекали фрагмент кишечника. Полученный материал выдерживали в течение 20 мин в гипотоническом растворе (0,07М КCl), а затем переносили в фиксатор Карнуа (3 части метанола и одна часть ледяной уксусной кислоты). Препараты изготавливали методом раскапывания на нагревательном столике при температуре 60°C. Для этого фрагмент ткани опускали в 70% раствор уксусной кислоты, вследствие чего ткань становилась более рыхлой, и исследуемый фрагмент распадался на отдельные клетки, образуя суспензию. Затем с помощью пастеровской пипетки на нагретые предметные стекла наносили капли диаметром до 1 см. Нанесенные капли тут же забирали пипеткой, благодаря чему на стекле оставалось небольшое количество отдельно лежащих клеток. Препараты высушивали и выдерживали в термостате в течение 3 недель при температуре 37°C.
Плоидность определяли путем подсчета количества хромосом (26 для диплоидов и 39 для триплоидов) в не менее чем семи метафазных пластинках для каждой особи. На изготовленных препаратах применяли Ag-окрашивание (серебрение) – метод, позволяющий выявить и определить положение района ядрышкового организатора (РЯО, сайты 18S + 28S рДНК) в хромосоме (Birshtein, 1984) (рис. 4). В ядрах соматических клеток, находящихся на стадии интерфазы, у диплоидных особей в результате Ag-окрашивания обнаруживаются ядрышки, заметные как два отчетливо окрашенных участка (рис. 3) (Schmid, 1982). У триплоидных особей обнаруживается три таких участка, что делает указанный метод применимым для определения плоидности. Для особей, на кариологических препаратах которых не оказалось метафазных пластинок, пригодных для анализа, плоидность определяли методом подсчета количества ядрышек не менее чем в 20 ядрах.
Мы использовали метод серебрения (Birshtein, 1984) с изменениями. Для окрашивания приготовили 30% водный раствор AgNO3, который перед применением фильтровали через шприцевой фильтр Simplepure NY (поры 0,22 μм). Затем 200 мг желатина растворяли в10 мл теплой воды. После растворения в желатин добавляли 100 μл муравьиной кислоты. На препараты наносили 75 μл раствора желатина, поверх которого наносили 150 μл раствора AgNO3. Затем препараты накрывали покровными стеклами и помещали во влажную светонепроницаемую камеру, которую оставляли в термостате на 3 мин при температуре 60°C, после чего препараты вынимали, промывали под проточной водой и высушивали. Далее препараты окрашивали в 2% растворе красителя Гимза. После высыхания препараты микроскопировали и подсчитывали количество ядрышек в интерфазных ядрах.
Результаты и обсуждение
На основании значения средней длины эритроцитов каждого метаморфа было сделано предположение, что все исследуемые особи являются диплоидными. Средние размеры эритроцитов метаморфов находились в диапазоне от 15,38 до 22,56 мкм, что гораздо ниже установленной границы между размерами эритроцитов ди- и триплоидов в 26–28 μм (Бондарева и др., 2012). Для сравнения, средние размеры эритроцитов взрослых диплоидных особей в норме составляют 21–26 мкм. Следует отметить, что изученный нами ранее один неполовозрелый триплоид из выборки в 17 особей, собранной в том же районе исследований, при длине тела 48 мм имел среднюю длину эритроцитов 23,5 мкм, что превышает средний размер эритроцитов всех рассмотренных в данной работе лягушек, но значительно меньше границы установленной для половозрелых особей (Темников и др., 2012).
В изученной выборке зависимость между размерами тела лягушек и средней длиной их эритроцитов (рис. 1) оказалась незначимой (р=0,36).
На мазках крови метаморфов мы наблюдали как мелкие овальные эритроциты, так и более крупные округлые клетки. Такая картина может быть связана с тем, что рассматриваемые особи только прошли метаморфоз и, возможно, еще имели кровяные клетки, характерные для головастиков (Hollyfield, 1966) (рис. 2).
Таким образом, определение плоидности неполовозрелых особей по средней длине эритроцитов оказывается ненадежным. Метод требует дополнительных исследований и определения границы между размерами эритроцитов разной плоидности у особей одного размерного класса или одной стадии развития. В то же время Ag-окрашивание кариологических препаратов соматических тканей позволяет определить плоидность даже при условии недостатка или отсутствия метафазных пластинок.
По результатам кариологического анализа все метаморфы, изученные в данной работе, также оказались диплоидными. При этом на препаратах, полученных от 13 особей, мы обнаружили метафазные пластинки, содержащие по 26 хромосом, что соответствует диплоидному набору. Для остальных 17 метаморфов заключение о плоидности было сделано на основании наличия двух ярко окрашенных ядрышек в интерфазных ядрах (рис. 3).
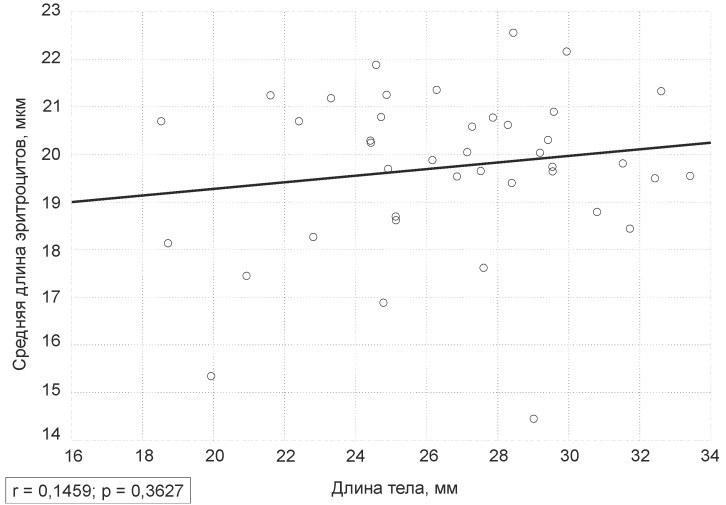
Рис. 1. График зависимости средней длины эритроцитов лягушек от длины их тела
Fig. 1. The plot of the frogs’ average red blood cells length dependence on their body length
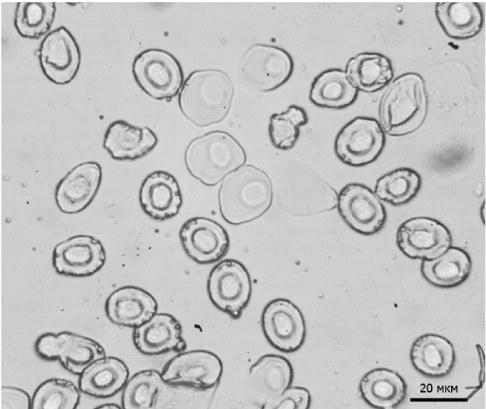
Рис. 2.Эритроциты, отличающиеся по форме и размеру, в мазке крови одной особи
Fig. 2.The red blood cells of different shapes and sizes in the blood smear of one individual
Обсуждая вопрос о причинах отсутствия триплоидов в изученной выборке, следует отметить, что, в отличие от процитированных выше работ (Borkin et al., 2004; Mezhzherin et al., 2010; Козак и др., 2012), в данной работе выборка состояла не только из гибридов, но и, вероятно, из сеголетков P. ridibundus. Определение видовой принадлежности метаморфов по морфологическим признакам крайне ненадежно в силу недостаточной проявленности их диагностичных признаков. Среди сеголетков всего гибридогенного комплекса зелёных лягушек триплоиды должны встречаться реже, чем в выборке половозрелых гибридов.
В районе, где были собраны метаморфы, соотношение представителей P. ridibundus и P. esculentus близко к 1:1 (Д.А.Шабанов, личное сообщение; Москалев и др., 2012). Объединив выборки, изученные в работе 2012 г. (Темников и др., 2012) и в данном исследовании, мы можем установить, что в объединенной группе из 47 сеголетков триплоидной оказалась 1 особь, а диплоидными – 46.
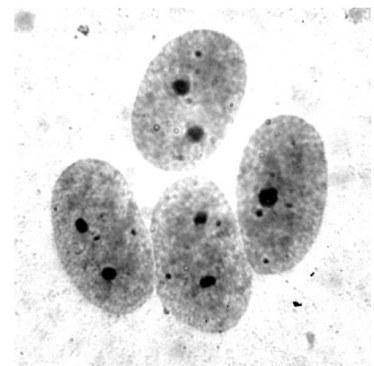
Рис. 3. Эпителиоциты тонкого кишечника с окрашенными нитратом серебра ядрышками
Fig. 3. Small intestine epithelial cells with silver nitrate stained nucleoli

Рис. 4. Метафазная пластинка (26 хромосом) с окрашенными нитратом серебра РЯО
Fig. 4. Metaphase plate (26 chromosomes) with silver nitrate stained NORs
Полученный результат дает основание на нынешнем этапе исследования отклонить гипотезу, которая проверялась в данной работе. Чтобы согласовать низкую долю триплоидов, зарегистрированную в нашем исследовании, с результатами публикаций, где указывается намного более высокая доля триплоидов среди P. esculentus, необходимо предположить действие каких-то неучтенных нами факторов. Изученная нами совокупность метаморфов или существенно отличается по доле входящих в ее состав триплоидов, или существенно отличается по соотношению P. ridibundus и P. esculentus от изученных ранее выборок более старших особей.
Для объяснения полученного результата можно выдвинуть два предположения.
1. Возможно, среди метаморфов и сеголетков доля триплоидов может быть не большей, а, наоборот, меньшей, чем в более старших возрастных классах. Известно, что при скрещивании гибридов, клонально передающих гаметы одного и того же родительского вида, выщепляются особи этого родительского вида. Этот феномен получил название гибридолиза. Однако диплоидные особи, возникшие в результате гибридолиза, оказываются маложизнеспособными и гибнут до достижения половозрелости (Plötner, 2005). Если этот процесс происходит в изученной нами популяционной системе достаточно часто, он может приводить к увеличению доли триплоидов в более старших возрастных классах за счет избирательной гибели определенной части диплоидных особей (представителей родительских видов, выщепившихся вследствие скрещивания гибридов). Поскольку значительная часть гамет, производимых и P. esculentus в изучаемой популяционной системе, принадлежит P. ridibundus (Боркин и др., 2005), существенная доля гибридолизных особей, принадлежащих к этому виду, может гибнуть после метаморфоза.
2. Полученный результат может быть следствием неравномерного распределения по пойме сеголетков, относящихся к родительскому виду или к триплоидным гибридам. Ранее было показано, что распределение половозрелых лягушек по территории поймы не является случайным, не зависимым от их генотипа. Представители разных форм и разных генотипов концентрируются в различных микроместообитаниях и демонстрируют различное поведение (Коршунов, 2010; Москалев и др., 2012).
Высказанные предположения в дальнейшем могут быть проверены на более широком материале с применением методов, использованных в данной работе.
Благодарности
Авторы выражают искреннюю благодарность доценту Д.А.Шабанову, как автору идеи и научному руководителю данной работы.
Список литературы
Бондарева А.А., Бибик Ю.С., Самило С.М., Шабанов Д.А. Цитогенетические особенности эритроцитов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex // Вісник Харківського національного університету імені В.Н.Каразіна. Серія: біологія. – 2012. – Вип.15 (№1008) – С. 116–123. /Bondareva A.A., Bibik Yu.S., Samilo S.M., Shabanov D.A. Tsitogeneticheskiye osobennosti eritrotsitov zelenykh lyagushek iz Seversko-Donetskogo tsentra raznoobraziya Pelophylax esculentus complex // Vіsnyk Kharkіvs'kogo natsіonal'nogo unіversytetu іmenі V.N.Karazіna. Serіya: bіologіya. – 2012. – Vyp.15 (№1008) – S. 116–123.
Боркин Л.Я., Зиненко А.И., Коршунов А.В. и др. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины // Мат. I конф. Українського Герпетологічного Товариства – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26. /Borkin L.Ya., Zinenko A.I., Korshunov A.V. i dr. Massovaya poliploidiya v gibridogennom komplekse Rana esculenta (Ranidae, Anura, Amphibia) na Vostoke Ukrainy // Mat. I konf. Ukrains'kogo Gerpetologіchnogo Tovarystva – K.: Zoomuzey NNPM NANU, 2005. – S. 23–26.
Искендерова Н.Э., Павлюк О.В., Чебукина М.В. Зависимость размеров эритроцитов от размеров зеленых лягушек (Pelophylax esculentus complex) // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 256–257. /Iskenderova N.Ye., Pavlyuk O.V., Chebukina M.V. Zavisimost' razmerov eritrotsitov ot razmerov zelenykh lyagushek (Pelophylax esculentus complex) // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – Kh.: FOP Shapovalova T.M., 2012 – S. 256–257.
Козак Н.А., Гладкова Ю.Д., Кулымова М.Д. Определение доли триплоидов среди Pelophylax esculentus в НПП «Гомольшанские леса» и его окрестностях // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 261–262. /Kozak N.A., Gladkova Yu.D., Kulymova M.D. Opredeleniye doli triploidov sredi Pelophylax esculentus v NPP «Gomol'shanskiye lesa» i yego okrestnostyakh // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – Kh.: FOP Shapovalova T.M., 2012 – S. 261–262.
Коршунов А.В. Экологические закономерности распределения Pelophylax esculentus complex в биотопах бассейна верхнего течения реки Северский Донец. Автореф. дисс. … к.б.н./03.00.16 – экология. – Днепропетровский национальный университет имени Олеся Гончара. – Днепропетровск, 2010. /Korshunov A.V. Ekologicheskiye zakonomernosti raspredeleniya Pelophylax esculentus complex v biotopakh basseyna verkhnego techeniya reki Severskiy Donets. Avtoref. diss. … k.b.n./03.00.16 – ekologiya. – Dnepropetrovskiy natsional'nyy universitet imeni Olesya Gonchara. – Dnepropetrovsk, 2010.
Михайлова О.В., Кечеджи А.Е., Шабанов Д.А. Изучение сперматогенеза у диплоидных Pelophylax esculentus (Amphibia, Anura) при помощи кариоанализа в раздавленных препаратах // Праці Українського герпетологічного товариства. – 2011. – №3. – С. 120–127. /Mikhaylova O.V., Kechedzhi A.E., Shabanov D.A. Izucheniye spermatogeneza u diploidnykh Pelophylax esculentus (Amphibia, Anura) pri pomoshchi karioanaliza v razdavlennykh preparatakh // Pratsі Ukrains'kogo gerpetologіchnogo tovarystva. – 2011. – №3. – S. 120–127.
Москалёв В.Б., Каширин О.О., Меламед Р.Г. Зависимость состава выборок зеленых лягушек от места и способа их сбора // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 271–272. /Moskalyov V.B., Kashirin O.O., Melamed R.G. Zavisimost' sostava vyborok zelenykh lyagushek ot mesta i sposoba ikh sbora // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – H.: FOP Shapovalova T.M., 2012 – S. 271–272.
Темников А.В., Вегерина А.О., Мелешко Е.В., Михайлова О.В. Изучение плоидности и размеров эритроцитов у неполовозрелых представителей Pelophylax esculentus complex // «Біологія: від молекули до біосфери». Мат. VII Міжнар. конф. мол. науковців. – Х.: ФОП Шаповалова Т.М., 2012 – С. 276–277. /Temnikov A.V., Vegerina A.O., Meleshko Ye.V., Mikhaylova O.V. Izucheniye ploidnosti i razmerov eritrotsitov u nepolovozrelykh predstaviteley Pelophylax esculentus complex // «Bіologіya: vіd molekuly do bіosfery». Mat. VII Mіzhnar. konf. mol. naukovtsіv. – H.: FOP Shapovalova T.M., 2012 – S. 276–277.
Шабанов Д.А., Коршунов О.В., Кравченко М.О. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми // Біологія та валеологія. – Вип.11. – Харків: ХДПУ, 2009. – С. 116–125. /Shabanov D.A., Korshunov O.V., Kravchenko M.O. Yakі zh zelenі zhaby naselyayut' Kharkіvs'ku oblast'? Termіnologіchnyy і nomenklaturnyy aspekty problemy // Bіologіya ta valeologіya. – Vyp.11. – Kharkіv: KhDPU, 2009. – S. 116–125.
Borkin L. J., Korshunov A. V., Lada G. A., Litvinchuk S. N., Rosanov J. M., Shabanov D. A., Zinenko A. I. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology, 2004. – Vol. 11, No 3. – P. 194–213.
Birstein V.J. Localization of NORs in karyotypes of four Rana species // Genetica. – 1984. – №64. – P. 149–154.
Hollyfield J.G. Erythrocyte replacement at metamorphosis in the frog Rana pipiens // Journal of Morphology. – 1966. – №119. – P. 1–6.
Mezhzherin S.V., Morozov-Leonov S.Yu., Rostovskaya O.V. et al. The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol.44, №4. – P. 212–216.
Plötner J. Die westpaläarktichen Wasserfrösche. – Bielefeld: Laurenti-Verlag, 2005. – 161S.
Schmid M. Analysis of the structure and variability of NORs in Anura // Chromosoma. – 1982. – №87. – P. 327–344.