|
|
|||
|
БІОСИСТЕМИ. БІОСФЕРА
ЕКОСИСТЕМИ. ПОПУЛЯЦІЇ
ОРГАНІЗМИ У ДОВКІЛЛІ
ЛЮДСТВО ТА ЙОГО ДОЛЯ
|
|||
|
IV-13. Правило конкурентного виключення Гаузе |
|||

IV-13. Правило конкурентного виключення Гаузе
Чим більш подібні два види, тим більше перетинаються їх потреби у ресурсах і тим більше конкуренція між ними. Найгостріша конкуренція — внутрішньовидова. Але особи одного виду, хоч і конкурують дуже гостро за ресурси, спільно формують наступне покоління. А що буде, якщо конкуренція двох різних видів досягне тієї ж гостроти, як і внутрішньовидова?
У 1931–1935 роках молодий радянський біолог Георг Францович Гаузе намагався експериментально перевірити рівняння конкуренції, виведені В. Вольтерра. Гаузе зрозумів, що для експериментів у галузі популяційної біології добре підходять найпростіші: їм потрібно зовсім невеликий обсяг середовища, а зміна їх поколінь відбувається набагато швидше, ніж у багатоклітинних тварин. Експерименти Гаузе, яке прославило його ім'я, виконані на інфузорії туфельці (Paramecium caudatum) та її найближчих родичах (рис. IV-12.1).
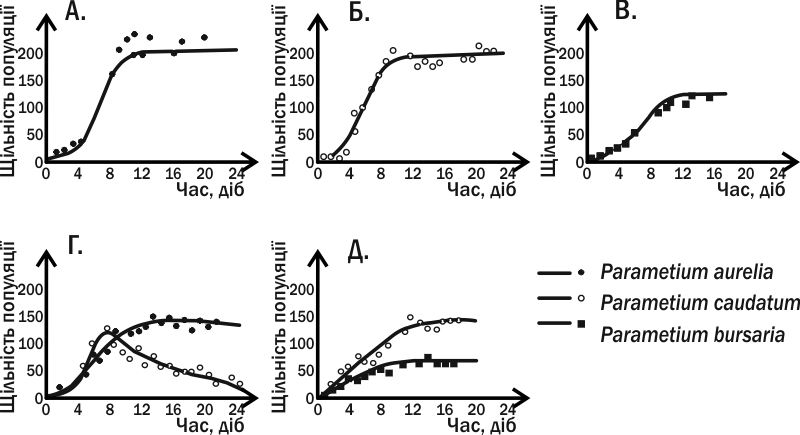
Рис. IV-13.1. Динаміка чисельності трьох видів інфузорій у дослідах Г.Ф. Гаузі. У варіантах експериментів А., Б. і В. види вирощували поодинці; у випадках Г. та Д. у пробірку заселяли відразу два види інфузорій
Гаузе вирощував три види інфузорій роду Paramecium у пробірках, куди він додавав вівсяне борошно і звідки періодично видаляв відходи. На вівсяніому борошні розвивалися дріжджі та бактерії, якими і харчувалися інфузорії. Три види роду Paramecium успішно розвивалися у монокультурі за таких умов. При сумісному утриманні P. aurelia завжди повністю витісняла P. caudatum. Однак при спільному культивуванні P. aurelia і P. bursaria два види могли існувати разом. Як встановив Гаузе, ці два види відрізняються за своїм способом харчування: перший вид харчувався бактеріями в товщі рідини, другий дріжджовими клітинами біля дна.
На підставі описаних спостережень був сформульований принцип конкурентного виключення, або правило Гаузе: два види, що займають ту саму екологічну нішу, не можуть стійко співіснувати в одному оселищі; співіснування видів можливе завдяки поділу їх ніш.
Правило конкурентного виключення є однією з можливих причин, що пояснюють надзвичайну різноманітність видів у природі. Якщо дві популяції одному оселищі не можуть займати одну нішу, їм доводиться її розділяти. Згодом кожна з таких популяцій спеціалізується у своїй ніші і може дати початок новому виду.
Екологічно перевірити дію правила Гаузе у природних умовах досить важко, проте деякі приклади його дії біологам відомі. З кінця XIX століття на більшій частині Європи широкопалий рак (Astacus astacus) витіснений довгопалим раком (A. leptodactylus). Екологічна ніша довгопалого раку перекриває нішу короткопалого, і при цьому вид-переможець є більш плодючим. Плітка (Rutilus rutilus) витісняє у багатьох водоймах краснопірку (Scardinius erythrophthalmus) та окуня (Perca fluviatilis). Екологічні ніші дорослих особин роз'єднані, а мальків — перекриваються. Мальки плітки виявляються більш конкурентноспроможними. Проте наявність багатьох прикладів конкурентного витіснення не доводить, що воно відбувається завжди. За той час, протягом якого біологи досліджують дію правила Гаузе, вдалося знайти як приклади його дії, так і ситуації, коли воно не спрацьовує.
Наприклад, в угрупованні планктонних водоростей поділ ніш не спостерігається. Причиною цього є шкидкі зміни середовища, де швидкоплинні умови дають перевагу то одному, то іншому виду, а також інтенсивний вплив з боку хижаків (зоопланктону та різноманітних фільтраторів), які не дають конкурентному виключенню стати головним чинником у формуванні угруповання. Однак у деяких випадках конкурентне витіснення відбувається і в планктонном угрупованні. Це відбувається при так званому «цвітінні» води, коли планктонні водорості вириваються з-під дії хижаків, які обмежують їх чисельність. В результаті «цвітіння» води в ній стають масовими лише кілька видів планктонних водоростей, які відрізняються один від одного за своїм способом життя — тобто відбувається конкурентне виключення.
Деякі види відносин можуть пов'язувати не тільки особин з різних популяцій, а й родичів, з однієї популяції. Наприклад, внутрішньовидова і міжвидова конкуренції схожі тим, що обидві обмежують кількість ресурсів, доступних для особини. Однак їх вплив на ширину екологічних ніш спрямований протилежно. Внутрішньовидова конкуренція розширює екологічні ніші: всім особинам не вистачає оптимальних для виду ресурсів, і деякі витісняються на край, де є хоч якісь більш-менш відповідні ресурси. Міжвидова конкуренція, згідно з правилом Гаузе, навпаки, звужує екологічні ніші. Ширина спостережених у природі ніш є, таким чином, результатом врівноваження двох протилежно спрямованих процесів — їх розширення через внутрішньовидової конкуренцію і звуження внаслідок міжвидової.
Цікавий приклад конкурентного виключення був досліджений у дослідах Томаса Парка з борошняними хрущаками. Два види цих жуків із сімейства чорнотілок (Tribolium confusum і T. castaneum) утримували в ящиках з борошном. Ці види конкурували за їжу (борошно) і, крім того, могли харчуватися один одним, причому жуки обох видів поїдали переважно особин виду-конкурента, а не свого виду (таке поєднання конкуренції з хижацтвом називається антагонізмом). Парку вдалося підібрати умови, за яких закономірно перемагав перший чи другий вид. Однак найцікавіше виявилося досліджувати результат конкуренції за проміжних умов, які не забезпечували безумовної переваги одного з видів (таблиця IV-13.1.).
Таблиця IV-13.1. Частка перемог двох видів борошняних хрущаків в експериментах Томаса Парка
|
«Клімат» (умови експерименту) |
% перемог |
|
|
Tribolium confusum |
Tribolium castaneum |
|
|
Спекотний вологий |
0 |
100 |
|
Помірний вологий |
14 |
86 |
|
Холодний вологий |
71 |
29 |
|
Спекотний посушливий |
90 |
10 |
|
Помірний посушливий |
87 |
13 |
|
Холодний посушливий |
100 |
0 |
Повторюючи дослідження за однакових умов, експериментатор реєстрував перемогу одного, то іншого виду з певною частотою. Зміна умов вирощування змінювала шанси кожного виду на перемогу, але в досить широкому діапазоні температури та вологості з певною ймовірністю перемогти могли обидва види. Постійним залишалося лише одне: через якийсь час у середовищі залишався лише один вид, а другий зникав.
Відмінності між видами однієї гільдії (сукупності видів, які використовують той самий ресурс) можуть бути опосередковані впливом хижака. Так, явна відмінність від сусіда може вивести вид зі сфери уваги хижака, що харчується сусідом! У Південно-Східній Азії сітчасті пітони живуть просто в поселеннях людини, крадуть і їдять кіз, свиней та собак і вкрай рідко нападають на людину. Це пов'язано з тим, що людина зовнішньо не схожа на своїх чотириногих родичів і не сприймається пітонами як жертва.
Внутрішньовидова конкуренція призводить до розширення ніші, що прагне розширитися до меж фундаментальної і навіть далі (внаслідок еволюції).
Оцінити ступінь конкуренції між видами можна, припустивши наявність певного лімітуючого рівня подібності. Якщо рівні відмінностей між сусідніми членами гільдії при порівнянному рівні внутрішньовидової мінливості приблизно однакові, це свідчить про високу роль конкуренції в диференціації ніш.
Джордж Хатчинсон продемонстрував подібність рівня дивергенції морфологічних ознак видів однієї гільдії на прикладі співвідношення розмірів ротових частин (тісно пов’язаних із параметрами здобичі) у співіснуючих комах, птахів та ссавців. З'ясувалося, що однотипні структури кожного такого виду більші, ніж у попереднього в 1,1–1,4 рази, в середньому — в 1,28 рази. Це різниця в розмірах, яка забезпечує різницю у вазі у 2 (2,09) рази. Правило Хатчинсона стосується розмірів харчодобувних структур і всього тіла у найрізноманітніших тварин: павуків, жуків-скакунів, ящірок, саламандр, білок, кажанів, пустельних гризунів, плодоядних голубів. З'ясовується, що при коефіцієнті варіації (CV) 5,5 для двох видів із відношенням розмірів 1,28 перекривання ніш становитиме 1–2%. Види розподіляються вздовж градієнта ресурсу невипадково. Співвідношення, близьке до 1,3, застосовується для опису відмінностей у серіях магнітофонів, верстатів і велосипедів! Можливо, воно відображає особливості нашого сприйняття, внаслідок яких інженери та маркетологи створюють класи виробів, які «природно» відрізняються один від одного.
Слід врахувати, що, на відміну від верстатів і велосипедів, представники видів із більшими розмірами тіла найчастіше проходять під час онтогенезу через розмірні класи, властиві меншим видам. У правилі Хатчинсона не враховані закономірності аллометричного зростання.
Угруповання можна уявити як n-мірний простір, зайнятий екологічними нішами різних видів. За умов дифузної конкуренції кожен вид взаємодіє з безліччю інших, конкуруючи з ними з різних чинників середовища. Успіх чи невдача закріплення виду в угрупованні насамперед залежить від відношень конкуренції та експлуатації з іншими видами. Уявіть собі ящик, заповнений надутими повітряними кульками: кожен із них тисне на решту. Якщо популяція якогось виду (кулька) «програє», тобто не зможе себе відтворювати (лусне), ефект від її зникнення призведе до зсуву рівноваги між рештою популяцій (переміщення всіх інших кульок).
З іншого боку, пари видів, що сильно перекриваються по одному з вимірів ніші, можуть істотно відрізнятися по іншому, послаблюючи цим конкуренцію. Можна говорити про диференціальне перекриття, при якому дифузно конкуруючі види ділять нішу за різними параметрами. Наприклад, хижаки частіше ділять ресурси за часом доби, оскільки їх видобутку також можуть характеризуватись певною добовою активністю. Для рослиноїдних тварин поділ за часом доби мало властивий. Поїкілотермні тварини більш диференційно використовують добу, ніж гомойотермні.
Що ближче сумарна чисельність членів гільдії до ємності середовища, то більшу роль може грати конкуренція. Можна грубо припустити, що у високих трофічних рівнях конкуренція гостріша.
Ступінь поділу екологічних ніш рослин загалом нижчий, ніж тварин.
Оскільки внутрішньовидова конкуренція виявляється сильнішою за міжвидову, розвиток з метаморфозом або екологічний статевий диморфізм можна розглядати як пристосування, що зменшують внутрішньовидову конкуренцію.