Кравченко М. А., Шабанов Д. А. Возможные пути трансформации популяционных систем Pelophylax esculentus cоmpleх (Ranidae, Anura, Amphibia) // Праці Українського герпетологічного товариства. – № 1, 2008. – С. 15-20.
УДК 57.055:597.851
ВОЗМОЖНЫЕ ПУТИ ТРАНСФОРМАЦИИ ПОПУЛЯЦИОННЫХ СИСТЕМ PELOPHYLAX ESCULENTUS COMPLEX (RANIDAE, ANURA, AMPHIBIA)
М. А. Кравченко1, Д. А. Шабанов2
Харьковский национальный университет им. В.Н. Каразина,
пл. Свободы, 4, Харьков, 61077 Украина;
E-mail: 1m_kravchenko@inbox.ru; 2d.a.shabanov@gmail.com
Возможные пути трансформации популяционных систем Pelophylax esculentus complex (Ranidae, Anura, Amphibia). Кравченко М. А., Шабанов Д. А.— Pelophylax esculentus комплекс включает два родительских вида: Pelophylax lessonae (Camerano, 1882) и Pelophylax ridibundus (Pallas, 1771)), а также их гибрид — Pelophylax esculentus (Linnaeus, 1758). Эта группа способна формировать многокомпонентные популяционные системы различного состава, которые существуют благодаря гемиклональному наследованию, характерному для диплоидных и триплоидных гибридных лягушек. Такие популяционные системы могут изменяться во времени. В работе представлена схема возможных путей их трансформации, которая может быть основанием для генеалогической классификации популяционных систем.
Ключевые слова: Pelophylax esculentus (Rana esculenta), гемиклональное наследование, трансформации популяционных систем.
Possible Ways of Transformation of Population Systems of Pelophylax esculentus complex (Ranidae, Anura, Amphibia). Kravchenko M. A., Shabanov D. A.— Pelophylax esculentus complex includes two parental species: (Pelophylax lessonae (Camerano, 1882) and Pelophylax ridibundus (Pallas, 1771)) and their hybrid — Pelophylax esculentus (Linnaeus, 1758). This group has a peculiar ability to form multicomponent population systems of various composition, existing due to the hemiclonal inheritance typical of diploid and triploid hybrid frogs. These population systems can undergo transformation in time. We present a scheme of possible ways of their transformation, which might become a basis of their genealogical classification.
Key words: Pelophylax esculentus (Rana esculenta), hemiclonal inheritance, transformation of population systems.
В состав комплекса среднеевропейских зеленыхлягушек, Pelophylax esculentus complex, входят прудовая (Pelophylax lessonae(Camerano, 1882), старое название— Rana lessonae) иозерная (Pelophylax ridibundus (Pallas, 1771) = Rana ridibunda) лягушки, а также их гибрид[1]— съедобная лягушка (Pelophylax esculentus (Linnaeus, 1758) = Rana esculenta).Все эти три формы способны населять одни местообитания и совместно размножаться, образовывая смешанные популяционные системы (далее — ПС). Одной из причин существования таких ПС является гемиклональное (клональное для отдельных геномов) наследование у гибридных лягушек. В ходе гаметогенеза у гибридов одни из родительских геномов могут элиминироваться, а другие (клональные) — переходить в гаметы без рекомбинации.
ПС P. esculentus complex обозначают заглавными буквами названий форм, которые их формируют (Uzzel, Berger, 1975; Лада, 1995). Например, выделяю тL-, R-, E-, L-R-, L-E-, R- E-, и R-E-L-типы популяций и ПС. Эта классификация не исчерпывает разнообразие известныж ПС зеленых лягушек. Так, в состав некоторый таких систем могут входить триплоидные особи; в таком случае, ПС из ди- и триплоидных гибридов можно отнести к Et-типу. В некоторый ПС гибридные лягушки могут быть представлены только одним полом. Так, R-Em- тип ПС соответствует смешанной системе из P. ridibundus и самцов P. esculentus (m— male— самец), а R-Ef-тип — аналогичной системе, в которой P. esculentus представлены лишь самками (f— female— самка) (Лада, 1995).
ОсобенностьюP. esculentuscomplexявляется высокое разнообразие их ПС даже в пределах относительно ограниченных территорий. Например, в Харьковской обл. зарегистрированы (А. В. Коршунов, устн. сообщ.) системы R-, L-, R-E-, R-Et-, R-Em- и R-E-L-типов (и остается дискуссионным существование систем E-типа). Следует отметить, что подавляющая часть этого региона находится за пределами ареала P. lessonae, и для этой области в целом характерны системы R-E-типа. Неожиданной находкой стало обнаружение в этом регионе значительной доли триплоидов среди гибридных лягушек (Borkin et al., 2004).
Замечательным свойством ПСP. esculentuscomplexявляется их способность к трансформации, которая отмечена в ряде работ и вытекает из теоретических представлений о природе таких систем (Holenweg Peter, Reyer, 2002; Vorburger, Reyer, 2003; Межжерин и др., 2005). Одним из примеров таких преобразований может быть судьба ПС Иськова пруда в окр. биостанции Харьковского университета, подробно рассмотренная в другой публикации (Шабанов и др., 2006). Это означает, что ПС зеленых лягушек следует рассматривать не только в статике, но и в динамике. Мы предложили возможную схему преобразования таких ПС, учитывающую специфику передачи клональных геномов (Шабанов и др., 2006; А. В. Коршунов, устн. сообщ.). В настоящей работе мы представляем расширенный и усовершенствованный вариант этой схемы (рис. 1).
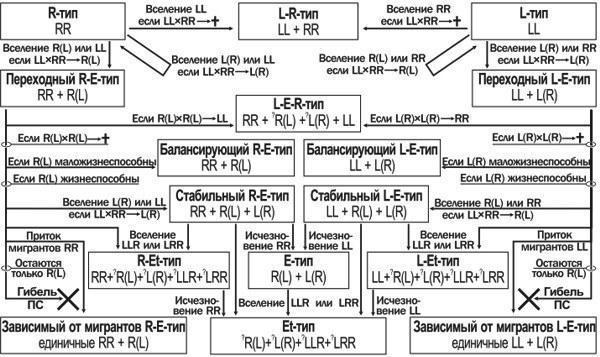
Рис. 1. Возможные пути трансформаций популяционных систем (ПС) Pelophylax esculentus complex.
Обозначения: L— геном Pelophylax Iessonae; R— геном Pelophylax ridibundus; L, R— неклональные (рекомбинирующиеся) геномы; (L), (R) — клональные (нерекомбинирующиеся) геномы
Основным методом данной работы является теоретический анализ литературныж и оригинальных данных о разнообразии популяционныгх систем зеленых лягушек с учетом представлений о закономерностях гемиклонального наследования, характерного для гибридных особей.
Отправной точкой в преобразовании ПС можно считать чистую популяцию родительского вида. Рассматривая возможность вселения в нее лягушек с иными генотипами, можно выделить три вероятных пути преобразования этой ПС. Если в исходную популяцию родительского вида попадут особи иного родительского вида, и образование гибридов между ними по каким-либо причинам окажется невозможным, эти два вида могут остаться в одном местообитании в том соотношении, которое будет определяться их конкурентоспособностью в данных условиях. Возврат к состоянию одновидовой популяции возможен при конкурентном вытеснении одного вида другим. Если же в исходную ПС попадут гибридные особи или особи другого родительского вида, скрещивание с которыми приведет к образованию гибридов, то судьба такой ПС определится тем, какие геномы будут передаваться клонально. Если клональным окажется конспецифичный родительскому геном, уже в следующем поколении система вернется в начальное состояние. Если клональный геном окажется гетероспецифичным, то ПС начнет трансформироваться. Соотношение особей родительского вида и гибридов от поколения к поколению будет изменяться. Из трех типов скрещивания, возможный в такой системе, два (родительский вид — родительский вид и гибрид — гибрид) не меняют соотношение основных форм в ПС, а третье (родительский вид — гибрид) приводит к возрастанию доли гибридов. В скрещиваниях особей родительского вида и гибридов с гетероспецифичными гаметами (RRх R(L) —>R(L), а также LLх L(R) —> L(R)) в потомстве присутствуют только гибриды. Это приведет к возрастанию доли гибридов в составе рассматриваемой ПС.
То, чем закончатся такие изменения, зависит от нескольких факторов. Первым из них является возможность выживания особей иного родительского вида, которые выщепляются при скрещивании гибридов, т. н. «гибридолизе» (Plotner, 2005): R(L) х R(L) —>- LLи L(R) х L(R) —>- RR. Если эти особи жизнеспособны, то система переходит к L-E-R-типу ПС. Однако зачастую особи, несущие два одинаковых клональных генома, оказываются нежизнеспособными. Этот эффект рассматривают как следствие так называемого «храповика Мюллера» — накопления неблагоприятных мутаций в геномах, которые передаются без рекомбинации (Хедрик, 2003).
Если жизнеспособность гибридов и особей родительского вида сравнима, трансформация ПС R-E- или L-E-типа может привести к полному вытеснению родительских особей. При условии нежизнеспособности потомства от скрещивания гибридов это означает гибель ПС. Она может сохраниться в нескольких случаях.
Во-первых, если гибриды обладают меньшей по сравнению с особями родительского вида жизнеспособностью. В этом случае преимущество гибридов в воспроизводстве может быть скомпенсировано отбором в пользу родительского вида. Соотношение двух форм в такой ПС будет определяться балансом двух процессов, меняющих его в противоположных направлениях.
Во-вторых, при появлении в ПС из родительского вида и гетероспецифичных ему гибридов гибридных особей с иным гаметогенезом такая ПС может перейти в стабильное состояние. Так, для ПС из RR, R(L) и L(R) (а также LL, L(R) и R(L))должны существовать стабильные соотношения названных форм, при которых состав каждого следующего поколения оказывается идентичен предыдущему. Такое стабильное состояние может быть достигнуто при вселении в ПС как диплоидных, так и, вероятно, триплоидных гибридов. Интересно, что вселение гибридов с конспецифичными родительскому виду гаметами в популяцию родительского вида не приведет ни к каким изменениям. Напротив, в системе из родительского вида и гибридов с гетероспецифичными ему гаметами вселение таких особей может привести к переходу ПС в стабильное состояние. Такая разница в реакции ПС на одно и то же воздействие — следствие частот- нозависимого отбора.
В-третьих, гибель рассмотренной ПС может предотвратить постоянный приток мигрантов, наблюдающийся между различными локальными популяциями в составе метапопуляцийP. esculentuscomplex. Результатом может быть возникновение зависимой от мигрантов ПС, состоящей из гибридов с одинаковой формой гаметогенеза. Такие ПС описаны как для R-E- (Шабанов и др., 2006), так и для L-E-систем (Межжерин и др., 2005).
Наконец, следует добавить, что потери особей родительского вида в ПС, включающих гибридов с различным гаметогенезом, могут приводить к возникновению «чистого» E- или Et- (при наличии триплоидов) типа ПС. Вселение в ПС определенных форм лягушек или их потеря в силу различных причин может обеспечить и иные, кроме рассмотренных, трансформации ПСP. esculentuscomplexв пределах указанной схемы.
Рассматривая показанные на рисунке 1 типы ПС, мы можем установить, что они отличаются по характеру своей устойчивости. Так, ПС родительского вида находится в состоянии неустойчивого равновесия — она может воспроизводиться в течение длительного времени, но вселение в нее даже одной гибридной особи с гетероспецифичными гаметами может запустить процесс ее необратимых трансформаций в переходной системе R-E- или L-E-типа. Состав ПС L-R-типа, а также R-E- и L-E-типов с пониженной жизнеспособностью гибридов отражает баланс противонаправленных процессов ее изменения. Зависимые от мигрантов системы с критично низкой репродуктивной численностью находятся в состоянии деградации, а ПС, включающие различные формы гибридов, которые отличаются по характеру гаметогенеза, могут быть по-настоящему стабильными.
Необходимо подчеркнуть, что в представленной на рисунке 1 не отражены следующие пять факторов, существенных с точки зрения трансформации ПС P. esculentus complex.
1. Причины возникновения в ПС в той или иной формы форме гибридных лягушек. Авторам не известно удовлетворительное объяснение причин того, какой геном при скрещивании родительских видов становится клональным. Существующие гипотезы (большая склонность к клональной передаче генома одного из родительских видов; клональная передача того генома, который принадлежит материнской особи и т. д.) не могут объяснить наблюдаемую совокупность фактов. Возникновение той или иной формы гибридов в показанной на рисунке 1 может быть результатом скрещивания родительских видов, миграции соответствующих особей из иных локальных популяций или (гипотетически) изменения характера клонального генома в ходе воспроизводства гибридов.
2. Различие мужских и женских клональных геномов. У лягушек гетерогаметным полом является мужской, и поэтому мужские клональные геномы у диплоидных лягушек могут нести только самцы, а женские — как самки, так и самцы. Если в генофонде ПС все клональные геномы являются мужскими, следует ожидать, что гибридные особи в ней будут представлены лишь самцами. Наличие ПС, где все гибриды являются самками, не может быть вызвано аналогичной причиной и требует особого объяснения.
3. Возможность неполной клональной передачи геномов с возникновением рекомбинантных особей (Межжерин и др., 2005). При скрещивании рекомбинантных гибридов с особями родительских видов возникает возможность передачи генетической информации через видовой барьер. Этот механизм, наряду с различными формами эволюции клональных геномов, должен приводить к увеличению разнообразия клональных геномов, передающихся внутри ПС.
4. Существование гибридов, производящих смесь гамет P. lessonae и P. ridibundus в определенном, специфичном для особи соотношении (Боркин и др., 2005).
5. Отличия особей двух родительских видов по параметрам, влияющим на воспроизводство ПС. Показанная на рисунке 1 схема симметрична, хотя в ряду P. lessonae—P. esculentus—P. ridibundus увеличивается средний размер половозрелых особей, растет привлекательность самок и снижается агрессивность самцов при спаривании. Следствием этой асимметрии является различная динамика преобразований и различные равновесные частоты разных форм лягушек в балансирующих и стабильных ПС.
С точки зрения авторов, приведенная гипотетическая схема нуждается в ее проверке как в ходе долгосрочных полевых наблюдений и проведения скрещивания и выращивания лягушек в экспериментальных условиях, так и в ходе математического моделирования.
Авторы выражают искреннюю благодарность А. И. Зиненко, А. В. Коршунову, Г. А. Мазепе и С. Ю. Морозову-Леонову за совместные полевые исследования, Л. А. Атраментовой, Л. Я. Боркину, Г. А. Ладе и С. Н. Литвинчуку за ценную критику и обсуждение результатов, а также М. В. Владимировой, Г. Н. Жолткевичу и А. А. Луцику за помощь в формализации представлений о популяционных системах лягушек и за математическое моделирование их трансформаций.
Боркин Л. Я., Зиненко А. И., Коршунов А. В. и др. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины // Матеріали Першої конф. Українського герпетол. т-ва. — К. : Зоомузей ННПМ НАНУ, 2005. — С. 23—26.
Лада Г. А. Среднеевропейские зеленые лягушки (гибридогенный комплекс Rana esculenta): введение в проблему // Флора и фауна Черноземья. — Тамбов, 1995. — С. 88—109.
Межжерин С. В., Морозов-Леонов С. Ю, Некрасова О. Д. и др. Пространственная структура гибридогенного комплекса зеленых лягушек Rana esculenta (Anura, Ranidae) на территории Украины // Матеріали Першої конф. Українського герпетол. т-ва. — К. : Зоомузей ННПМ НАНУ, 2005. - С. 110-144.
Хедрик Ф. Генетика популяций. М. : Техносфера, 2003. — 592 с.
Шабанов Д. А., Зиненко А. И., Коршунов А. В. и др. Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы // Вісн. ХНУ ім. В. Н. Каразіна. Сер. Біологія. — 2006. — Вип. 3 (№ 729). — С. 208-220.
Borkin L. J., Korshunov A. V., Lada G. A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology. — 2004. —11,N 3. — P. 194—213.
Holenweg Peter A.-K, Reyer H.-U, Abt Tietje G. Species and sex ratio differences in mixed populations of hybridogenetic water frogs: the influence of pond features // Ecoscience. — 2002. —9.— P. 1—11.
Plotner J. Die westpalaarktichen Wasserfrosche. — Bielefeld : Laurenti, 2005. — 161 s.
Uzzell T. M, Berger L. Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenic associate Rana esculenta // Proc. Acad. Nat. Sci. Phila. — 1975. —127.— P. 13—24.
Vorburger C., Reyer H.-U. A genetic mechanism of species replacement in European waterfrogs? // Conservation Genetics. — 2003 —4.— P. 141—155.
[1]В данном случае термином «гибрид» обозначается эволюционно-таксономическая единица видового ранга, имеющая научное название Pelophylax esculentus (Dubois, 1991, 1998; но см. Frost, 2006.) — Прим. редакции.