Кравченко М. А., Шабанов Д. А., Владимирова М. В., Жолткевич Г. Н. Исследование устойчивости гемиклональных популяционных систем гибридогенного комплекса зеленых лягушек при помощи имитационного моделирования // Вісник Дніпропетровського університету. Біологія. Екологія. – 2011. – Вип. 19, т. 1. – С. 51–64.
УДК: 004.942:597.851
М. А. Кравченко, Д. А. Шабанов, М. В. Владимирова, Г. Н. Жолткевич
Харьковский национальный университет имени В. Н. Каразина
ИССЛЕДОВАНИЕ УСТОЙЧИВОСТИ ГЕМИКЛОНАЛЬНЫХ ПОПУЛЯЦИОННЫХ СИСТЕМ ГИБРИДОГЕННОГО КОМПЛЕКСА ЗЕЛЕНЫХ ЛЯГУШЕК ПРИ ПОМОЩИ ИМИТАЦИОННОГО МОДЕЛИРОВАНИЯ
Проанализировано понятие устойчивости в экологии и математике. Обосновывается трактовка устойчивости как способности системы оставаться в ограниченной зоне фазового пространства, соответствующей определенному типу рассматриваемых систем. Этот подход применен для описания изменений состава гемиклональных популяционных систем гибридогенного комплекса зеленых лягушек (Pelophylax esculentus complex). Использована имитационная модель, построенная на рекуррентных разностных уравнениях в MS Excel. Описана динамика переходов в части фазового пространства гемиклональных популяционных систем зеленых лягушек. Проанализирована типология состояний устойчивости биосистем.
М. О. Кравченко, Д. А. Шабанов, М. В. Владимирова, Г. М. Жолткевич
Харківський національний університет імені В. Н. Каразіна
ДОСЛІДЖЕННЯ СТІЙКОСТІ ГЕМІКЛОНАЛЬНИХ ПОПУЛЯЦІЙНИХ СИСТЕМ ГІБРИДОГЕННОГО КОМПЛЕКСУ ЗЕЛЕНИХ ЖАБ ЗА ДОПОМОГОЮ ІМІТАЦІЙНОГО МОДЕЛЮВАННЯ
Проаналізовано поняття стійкості в екології та математиці. Обґрунтовується трактування стійкості як здатності системи залишатися в обмеженій зоні фазового простору, що відповідає певному типу систем, що розглядаються. Цей підхід застосований для опису змін складу геміклональних популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex). Використана імітаційна модель, яка побудована на рекурентних різницевих рівняннях в MS Excel. Описана динаміка переходів у частині фазового простору геміклональних популяційних систем зелених жаб. Проаналізовано типологію станів стійкості біосистем.
INVESTIGATION OF THE STABILITY OF HEMICLONAL POPULATION SYSTEMS OF WATER FROGS HYBRIDOGENEOUS COMPLEX BY THE MEANS OFSIMULATION MODELING
M. O. Kravchenko, D. A. Shabanov, M. V. Vladimirova, G. M. Zholtkevych
V. N. Karazin Kharkiv National University
The concept of stability used in ecology and mathematics is analyzed. Stability is interpreted as the ability of the system to remain in a restricted zone of a phase space that corresponds to a certain type of systems. This approach is applied to describe changes in the structure of hemiclonal population systems of hybridogenous complex of water frogs (Pelophylax esculentus complex). Simulation model of these population systems built on a recursive difference equations in MS Excel is used. The dynamics of transitions in the part of phase space of systems hemiclonal populations of water frogs is described. Typology of stabile states of biosystems is analyzed.
Введение
Для подавляющего большинства организмов с половым размножением характерно образование видовых популяций, которые входят в состав биотических сообществ. Особи внутри популяции объединены общностью генофонда и совместно участвуют в популяционном воспроизводстве. Популяции в сообществе состоят в разнообразных отношениях, от конкуренции до эксплуатации и мутуализма. Часть сообщества, объединяющая конкурирующие популяции, использующие один и тот же ресурс, могут образовывать гильдии. Перечисленные уровни организации биосистем (популяция, гильдия, сообщество) можно считать типичными. Однако для некоторых групп организмов характерны и иные, необычные уровни организации биосистем. К их числу принадлежат европейские зеленые лягушки, Pelophylax esculentus complex.
Название Pelophylax esculentus (Linnaeus, 1758) принадлежит не виду, а гибриду, который образуется при скрещивании двух родительских видов: Pelophylax lessonae (Camerano, 1882) и Pelophylax ridibundus (Pallas, 1771). Для P. esculentus характерно гемиклональное наследование, при котором в гаметы (половые клетки) переходит или геном (хромосомный набор) P. lessonae, или геном P. ridibundus, а не рекомбинантные геномы, состоящие из смеси родительских геномов, как это происходит у подавляющего большинства организмов с половым размножением [17, 20]. Мы принимаем для генома P. lessonae обозначение L, а для генома P. ridibundus — R, причем клональность мы обозначаем заключением символа генома в скобки: (L) или (R). При скрещивании с особями родительских видов тех гибридов, которые передают гетероспецифичный геном (т.е. геном другого родительского вида), все потомство состоит из гибридов: RR × R(L) → R(L). Клональный геном (L) в обоих поколениях одинаков, а рекомбинантные геномы — R — «пересобираются» заново при передаче из поколения в поколения у особей родительских видов.
Для зеленых лягушек характерно образование популяционных систем, где совместно обитают и размножаются как представители родительских видов, так и разнообразные гемиклональные гибриды. Например, в Северско-Донецком центре разнообразия зеленых лягушек [17], помимо особей с генотипом RR (т.е. представителей P. ridibundus) встречаются диплоидные гибриды (L)R, L(R), (L)(R), а также триплоидные гибриды LLR и LRR. Среди неполовозрелых особей встречаются одиночные тетраплоиды LLRR и особи LL (т.е. P. lessonae). Важно, что особи P. lessonae, возникающие от скрещивания особей (L)R, оказываются нежизнеспособными. Нежизнеспособность особей родительских видов, получивших оба своих генома от родителей-гибридов, регистрировались и в других регионах [20].
Очевидно, что подобные системы, где из поколения в поколение передаются как клональные, так и рекомбинантные геномы разных видов, принадлежат к особой группе биосистем и заслуживают особого названия. Мы предложили [16, 17] называть их гемиклональными популяционными системами — ГПС.
ГПС не являются популяциями, так как они включают особи разных видов. ГПС не являются гильдиями и сообществами, так как объединены процессом совместного воспроизводства. Динамика ГПС отличается от динамики популяций, гильдий и сообществ. Одним из следствий того, что ГПС является особым уровнем организации биосистем, существование которого было осознано совсем недавно, является их значительно меньшая изученность по сравнению с типичными уровнями биосистем.
Целью данной работы является анализ различных состояний ГПС с точки зрения их устойчивости. Предварительным условием такого анализа является рассмотрение смысла, который вкладывается в понятие «устойчивость».
Анализ понятия «устойчивость»
Понятие устойчивости стало одним из важнейших экологических понятий с ранних этапов развития этой науки. Оно было важным для теории климакса растительных сообществ Ф. Клементса, появившейся в начале XX века [19, цит. по 13]. В. Н. Сукачев [14] определял устойчивость растительного сообщества как способность «сохранять свой состав и строй в течение продолжительного времени. Эта устойчивость обусловлена способностью сообщества к самовозобновлению».
Несмотря на интерес к понятию устойчивости в экологии, считать его ясным, увы, преждевременно. Приведем два высказывания авторитетных авторов. «К сожалению, термин «устойчивость» слишком часто оставался расплывчатым и неопределенным» [11, с. 324]. «Экологи удивительно невежественны во всем, что касается устойчивости природных систем: какие внутренние механизмы сообществ в этом участвуют и как они действуют?» [12, с. 394].
Р. Риклефс проводит анализ понятия «устойчивость». «Устойчивость, присущую данной системе, можно определить как отношение между изменчивостью в среде и изменчивостью в самой системе» [12, с. 378], подчеркивая, что это определение (фактически, говорящее о буферных свойствах системы) трудно приложить к популяции и сообществу. Проще понять этот подход на примере, относящемся к организменному уровню: Риклефс приводит данные о динамике климатических параметров и динамике толщины годовых приростов деревьев. Выясняется, что с этой точки зрения деревья могут быть более или менее устойчивыми к изменению внешних условий. Затухание возмущений (падение количества осадков на 50% приводит к снижению растительной продукции на 25% и к падению численности растительноядных животных на 10%) позволяет говорить о работе внутренних механизмов, поддерживающих устойчивость экосистемы.
Риклефс выделяет три типа состояний, связанных с устойчивостью: устойчивое равновесие (возмущения компенсируются), неустойчивое равновесие (возмущения усиливаются) и нейтральное равновесие (возмущения вызывают в системе несущественные для ее функционирования изменения). В конечном итоге, Риклефс формулирует определение, характеризующее двойственность рассматриваемого понятия: «Устойчивость – это внутренне присущая системе способность выдерживать изменение, вызванное извне, или восстанавливаться после него» [12, с. 379].
Р. Уиттекер трактует устойчивость прежде всего как буферность. «Мы будем называть процессы, которые сокращают потери популяции, когда среда становится более неблагоприятной, буферностью популяций» [15, с. 58].
Понятийное выражение двойственности феномена устойчивости, на которое указывал Р. Риклефс, дает Ю. Одум [10, с. 66], рассматривающий резистентную устойчивость (способность сопротивляться пертурбациям) и упругую устойчивость (способность восстанавливаться после нарушения) как проявления стабильности.
М. Бигон и соавторы [3, с. 325-327, а также 19, p. 576-577], анализируя понятие устойчивости, выделяют три пары понятий:
— упругость (resilence) как мера способности возвращаться в исходное состояние и сопротивление (resistance) как мера способности избегать изменений;
— локальная (local) и общая устойчивость (global stability), различающиеся по уровню возмущений, которые способны компенсировать рассматриваемые системы;
— динамическая хрупкость (dynamic fragility) или прочность (robustness) отражают отношения рассматриваемой системы с внешней средой: способность системы сохраняться в узком наборе внешних условий или в их широком диапазоне.
Авторы приводят графические аналогии для пояснения использованных ими терминов. Как указывают Бигон и соавторы, изучение устойчивости сообществ ведется в значительной мере с использованием математического моделирования.
В. Г. Стороженко [13] приводит обширный перечень параметров, сохранение которых является проявлением устойчивости биогеоценозов. В их числе количество видов и сами эти виды, взаимосвязи внутри системы, численность популяций, особенности функционирования, подавление вредных факторов и т.д. Главной задачей анализа, который проводит В. Г. Стороженко, является поиск критериев устойчивого лесного сообщества. Автор не рассматривает и не классифицирует те состояния системы, которые далеки от климакса.
Как видно из приведенного обзора, большинство работ, посвященных проблеме экологической устойчивости, характеризует условия, при которых те или иные сообщества (прежде всего – лесные) оказываются устойчивыми. Однако кроме состояния истинной устойчивости (устойчивого равновесия), как наиболее ценного состояния биосистемы, следует исследовать и иные ее возможные состояния. Можно ли разделить совокупность возможных состояний системы на типы, отличающиеся с точки зрения их динамики и устойчивости?
Для решения этой проблемы необходимо привлечь аппарат анализа устойчивости, развитый в математике. Рассматриваемому понятию «не повезло» и здесь: «устойчивость – термин, не имеющий четко определенного содержания» [9, с. 604]. Классическим определением понятия «устойчивость» является то, которое было дано в 1892 г. создателем теории устойчивости движения Александром Михайловичем Ляпуновым (доцентом, а позже – профессором Харьковского университета, ныне носящего имя В. Н. Каразина). Устойчивость определялась А. М. Ляпуновым с точки зрения воздействий возмущений на движение с определенными начальными условиями [8]. Применение такого подхода к описанию динамики биосистем сталкивается со значительными сложностями и не вполне соответствует сложившейся традиции. По нашему мнению, из математических подходов к рассматриваемому понятию для экологии полезнее всего представление об устойчивости по Ж. Л. Лагранжу, которое является частным случаем устойчивости по А. М. Ляпунову [5]. С этой точки зрения, устойчивость является способностью системы оставаться в ограниченной области своего фазового пространства.
Фазовым пространством в данном случае называется совокупность возможных состояний системы, описанных с помощью набора ее переменных состояния. Выбор переменных, задающих фазовое пространство, позволяет, например, рассматривать устойчивость сообщества и с точки зрения числа его видов, и с точки зрения постоянства его структуры. Суть феномена устойчивости остается при этом одной и той же, меняется только набор координат, в котором исследуется траектория изменений исследуемой системы.
Такое понятие устойчивости вполне соответствует практике его применения в экологических исследованиях. Например, хотя Ф. Клементс и не формулировал свои взгляды таким образом, устойчивость климаксного сообщества по Клементсу можно определить как способность сообщества сохранять с течением времени постоянным свой тип (на какой бы типологии не основывалось выделение этого типа).
К описанному подходу близка трактовка устойчивости геосистем, принадлежащая А. Д. Арманду [1]. Арманд выделяет устойчивость I, т.е. неизменность, инертность, а также устойчивость II, т.е. способность противостоять возмущениям. С устойчивостью II связано понятие бассейна устойчивости. Это «область в абстрактном многомерном пространстве, построенном на осях переменных состояния, из всех точек которой переходный процесс приводит систему в одно и то же равновесное или гомеостатическое состояние».
Арманд выделяет несколько типов состояний равновесия, при которых не происходит саморазвития рассматриваемой системы. Это равновесие устойчивое, неустойчивое, безразличное, циклическое, динамическое и подвижное. Первые три типа равновесия соответствуют тем, которые выделяет Р. Риклефс. В случае циклического равновесия система регулярно возвращается в одно и то же состояние. Сложнее всего понять два последних типа. «Динамическое равновесие — состояние системы, в котором противоположно направленные процессы (привнос и вынос вещества и энергии, организация и дезорганизация) взаимно скомпенсированы. Подвижное равновесие — траектория системы в пространстве состояний, по которой система следует за изменением входных переменных (факторов), сохраняя в каждый момент времени состояние равновесия устойчивого» [1]. Когда речь идет о биосистемах, всякое их равновесное состояние является динамическим. Альтернативой динамическому равновесию является устойчивость I, т.е. статичность. С нашей точки зрения, для того, чтобы описать разницу между устойчивым и подвижным равновесием, необходимо разделить влияющие на систему факторы на внешние и внутренние. Устойчивое равновесие — это результат уравновешивания внутренних факторов, а подвижное равновесие является следствием его смещения внешними факторами.
Итак, понятие устойчивости системы оказалось тесно связанным с типологией состояний биосистемы. Разделив пространство состояний системы на бассейны устойчивости (между которыми располагаются границы устойчивости [2]), можно получить динамическую типологию ее состояний. При решении этой задачи для ГПС Pelophylax esculentus complex будет полезной имитационная модель рассматриваемых систем.
Имитационное моделирование ГПС Pelophylax esculentus complex
Для моделирования динамики ГПС зеленых лягушек мы разработали совокупность требований пользователя к модели (URD, User Requirements Definition). Изучение динамики ГПС зеленых лягушек осуществлялось методом имитационного моделирования. Для верификации результатов моделирования мы разработали две диверсные имитационные модели. Совместно с А. А. Луциком мы создали мультиагентную имитационную модель ГПС [4]. Более удобной в работе в силу легкости внесения в нее изменений оказалась модель ГПС, основанная на рекуррентных разностных уравнениях, которая реализована в среде MicrosoftOfficeExcel[6]. Одна из версий этой модели (названная Batrachometrics-2010) находится в свободном интернет-доступе на сайте Центральной научной библиотеки Харьковского национального университета им. В. Н. Каразина по адресу https://dspace.univer.kharkov.ua/handle/123456789/2037. Важно подчеркнуть, что обе реализации модели, как мультиагентная, так и основанная на разностных уравнениях в среде MSExcel, генерируют качественно сходную динамику моделируемых систем.
В модели постулируется существование ГПС, состоящей из различных форм лягушек. В ГПС передаются геномы, отличающиеся по видовой принадлежности (P. lessonae и P. ridibundus), характеру передачи (клональные и рекомбинантные) и полу (женские и мужские). От генома (набора генотипов) и возраста особи зависят параметры ее жизнеспособности: максимальная продолжительность жизни, срок наступления половозрелости, выживаемость в отсутствие конкуренции, конкурентоспособность, привлекательность для партнеров, плодовитость и, в случае самцов, вероятность повторных амплексусов (соединений пар).
Шаг модели соответствует одному году. В течение каждого года работы модели имитируется снижение численности каждого возрастного класса каждого генотипа в течение зимовки. Емкость среды ограничивает суммарную численность или биомассу всех особей в составе ГПС. Если численность популяции превышает емкость среды, происходит ее конкурентное сокращение. Численность каждой группы особей снижается обратно пропорционально конкурентоспособности ее представителей. Вступающие в амплексус пары образуются в зависимости от привлекательности особей каждой группы. В размножении участвуют все представители пола, который при наблюдаемой в ГПС численности способен вступить в меньшее количество амплексусов, а также соответствующее количество представителей другого пола. Для всех возможных сочетаний генотипов родителей задаются генотипы возникающих потомков. После имитации выживания в зимовке, конкурентного сокращения, образования пар их размножения и образования потомков модель переходит на следующий шаг (в следующем году), и вся описанная последовательность расчетов повторяется.
Для работы модели необходимо задать определенные значения параметров жизнеспособности для всех рассматриваемых форм лягушек (подробнее — см. [6]). Далее в данной статье, кроме оговоренных случаев, используются значения, выставленные в Batrachometrics-2010 по умолчанию.
Фазовое пространство и типология ГПС Pelophylax esculentus complex
Тот факт, что зеленые лягушки образуют популяционные системы, отличающиеся по своему составу, известен уже несколько десятилетий. Самая распространенная типология популяционных систем лягушек предложена в 1975 г. при участии открывателя гибридной природы P. esculentus Л. Бергера [21]. В этой типологии буква L обозначает присутствие в популяционной системе P. lessonae, R — присутствиеP. ridibundus, а E — присутствие P. esculentus. Наличие среди гибридов триплоидных особей обозначается буквой t, присутствие исключительно самок — буквой f, и исключительно самцов — буквой m [7]. Например, можно утверждать, что в пойме реки Северский Донец в окрестностях биостанции Харьковского университета (с. Гайдары Змиевского района Харьковской области) обитает популяционная система REt-типа [17].
Описанный подход широко используется при изучении распространения различных форм зеленых лягушек, однако он не учитывает то обстоятельство, что особи P. esculentus могут производить гаметы с различными клональными геномами. Состав гамет определяет и состав будущего потомства, и особенности динамики рассматриваемой ГПС.
Мы предлагаем использовать для характеристики ГПС состав передаваемых в ней геномов. В данной работе мы рассмотрим ГПС RE-типа, где из поколения в поколение передаются рекомбинантные геномы R, и могут передаваться клональные геномы (L) и (R). Построим двумерное фазовое пространство, осями которого являются доли женских клональных геномов (XL) и (XR) в общем количестве геномов (рис. 1). Началу координат в такой системе соответствует популяция R-типа, состоящая из особей P. ridibundus. На прямой, соединяющей точки на осях координат, которые соответствуют 100% (R) и 100% (L), находятся ГПС E-типа. Точки выше этой прямой не определены, т.к. сумма передающихся клональных геномов для них превосходила бы 100%. Остальные точки описанной плоскости соответствуют ГПС RE-типа.
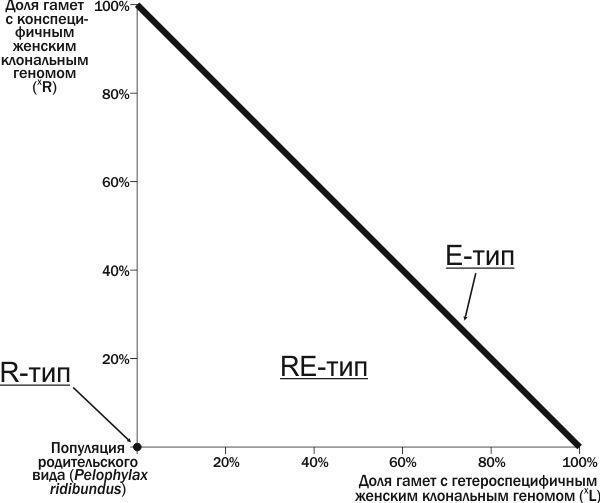
Рис. 1. Типы популяционных систем на фазовом пространстве ГПС, состоящей из особей P. ridibundus, а также гибридов, передающих женские клональные геномы обоих видов
Набор состояний ГПС, показанный на рис. 1, — это лишь часть возможного разнообразия ГПС зеленых лягушек. В ГПС с одним родительским видом могут передаваться и женские, и мужские клональные геномы. Помимо RE-типа ГПС можно рассматривать также LE-тип (набор фазовых состояний которого симметричен набору состояний RE-типа), а также LER-тип. Наконец, в ГПС могут присутствовать различные триплоидные гибриды.
Характеристика понятия устойчивости требует конкретизации возмущений, влияющих на рассматриваемую систему. Для состава ГПС возмущением является вызванное внешними обстоятельствами изменение численности представленных в ней форм особей. В рассматриваемом фазовом пространстве такими возмущениями являются добавления в ГПС особей P. esculentus, передающих женские клональные геномы: ♀♀(XL)XR, ♂♂(XL)YR, ♀♀XL(XR) и ♂♂YL(XR).
Трансформации ГПС RE-типа с женскими клональными геномами
Популяция R-типа может находиться в стационарном состоянии неограниченное время. При добавлении ♀♀XL(XR) или ♂♂YL(XR) система через несколько лет возвращается к прежнему состоянию. В отношении этого возмущения ГПС находится в состоянии устойчивого равновесия.
При попадании в популяцию P. ridibundus хотя бы одной особи, передающей геном (XL), в ней начинаются необратимые преобразования. Все потомство от скрещивания особей родительского вида и гибридов оказывается гибридным; вследствие этого доля гибридов от поколения к поколению возрастает. По мере накопления в ГПС гибридов, переносящих геном (XL), все большая доля скрещиваний происходит между ними и приводит к появлению нежизнеспособных особей (XL)(XL). Заканчивается описанный процесс гибелью ГПС (рис. 2).
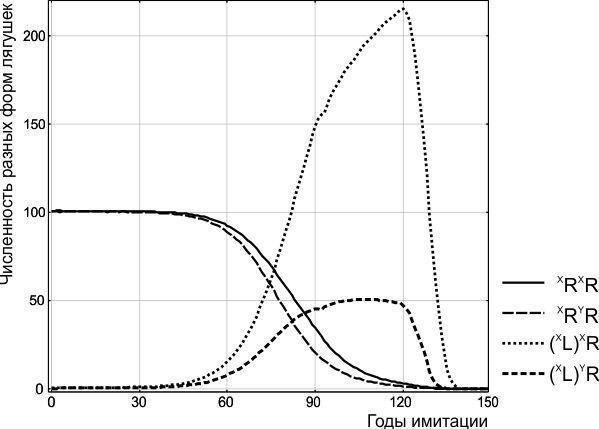
Рис. 2. Трансформации ГПС, полученной из популяции P. ridibundus, в которую на первом году имитации добавлена одна ♀ (XL)XR; около 140-го года ГПС погибает
На рис. 2 иллюстрируется переход системы из состояния неустойчивого к добавлению генома (XL) равновесия в состояние направленного изменения, характеризующееся возрастанием доли особей (XL)XRи (XL)YR. Результатом этого процесса является гибель всей системы.
По мере нарастания в ГПС доли геномов (XL) система теряет устойчивость к попаданию в нее геномов (XR), так как по принятым в модели Batrachometrics-2010 умолчаниям, особи (XL)(XR) являются жизнеспособными. Чтобы пояснить свойства системы на рассматриваемом участке фазового пространства, можно, вслед за М. Бигоном и соавторами [3], воспользоваться физической аналогией (рис. 3). Начальное состояние системы на рис. 3 является устойчивым равновесием по отношению к добавлению геномов (XR), и неустойчивым — по отношению к добавлению геномов (XL). В целом такое состояние следует характеризовать как неустойчивое равновесие. При попадании геномов (XL) система «скатывается» в сторону увеличения их доли и теряет устойчивость к попаданию в нее геномов (XR).
На 90-м году имитации внесение в показанную на рис. 2 ГПС как ♀ XL(XR), так и ♂YL(XR), не приводит ни к каким существенным изменениям. На 100-м и 110-м годах и то, и другое воздействие приводят к тому, что ГПС переходит к иному состоянию, при котором в ней передается значительное количество клональных геномов обоих видов (рис. 4).
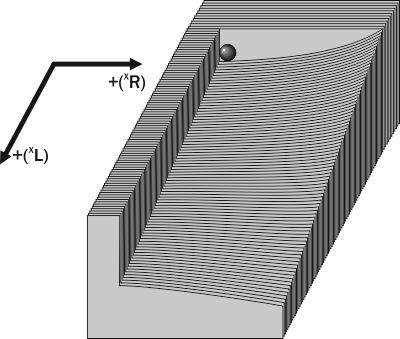
Рис. 3. Физическая аналогия свойств участка фазового пространства ГПС лягушек (см. рис. 1) вблизи начала координат (популяции P. ridibundus, показанной в виде шарика)
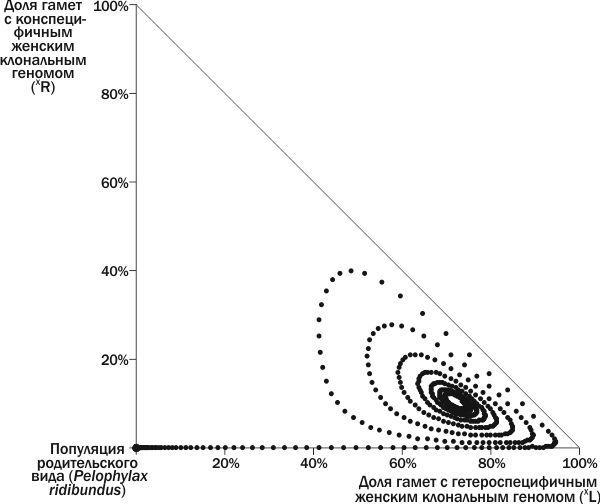
Рис. 4. Динамика ГПС в фазовом пространстве. В начале процесса к популяции P. ridibundus добавлена одна ♀ (XL)XR, на 100-м году имитации добавлена одна ♀ XL(XR). Продолжительность показанных на рисунке событий — 500 лет имитации
Как видно по рис. 4, ГПС, в которой передаются клональные геномы обоих видов, претерпевает затухающие циклические изменения своего состава. При анализе 500 лет имитации можно было бы допустить, что ГПС в конечном итоге переходит к состоянию циклического равновесия, при котором она движется по некоей замкнутой траектории. Приводя аналогии из динамики иных биосистем, можно указать, что состоянию циклического равновесия соответствуют циклические сукцессии биогеоценозов или незатухающие циклические колебания двух видов в системе «хищник-жертва», с запаздыванием реагирующих на изменения численностей друг друга [11, с. 231].
Однако в случае рассматриваемой модели ГПС равновесие не циклическое, и примерно к 1000-му году ГПС достигает точки устойчивого равновесия. Состав нерестового стада в этой точке таков: 10,5%♀♀ XRXR; 6,9%♂♂XRYR; 53,9%♀♀ (XL)XR; 16,7%♂♂(XL)YR; 8,6 % ♀♀ XL(XR); 3,4%♀♀ (XL)(XR). Этот состав ГПС очень далек от равенства полов: в нем всего лишь 23,6% самцов и 76,4% самок. Тем не менее, все ГПС, находящиеся внутри петлеобразной части траектории, показанной на рис. 4, приходят именно к этому состоянию.
А что происходит с ГПС, находящимися вне показанной на рис. 4 «петли»? Рассмотрим случай, когда геном (XR) добавляется лишь на 120-м году (рис. 5).
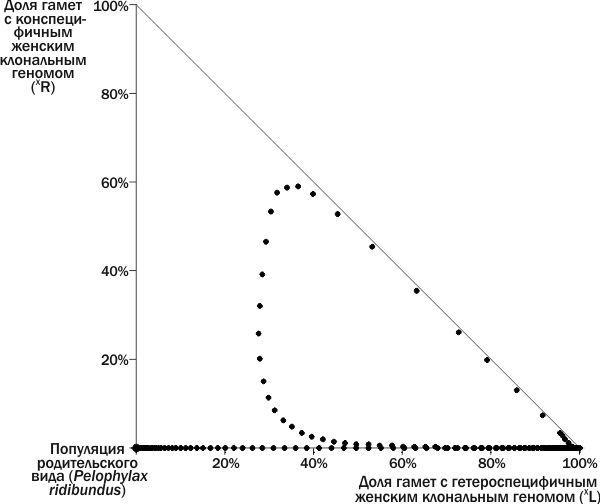
Рис. 5. Динамика ГПС в фазовом пространстве. В начале процесса к популяции P. ridibundus добавлена ♀ (XL)XR, а на 120-м году — ♀ XL(XR). Описав в фазовом пространстве петлю, ГПС на 215-м году имитации попадает в правый угол, т.е. точку, где все 100% передающихся геномов — это геномы (XL), и погибает
Разнообразие состояний ГПС зеленых лягушек с точки зрения их устойчивости
Исходя из изложенного, мы можем выделить на изученном фазовом пространстве состояний ГПС положения равновесия и их бассейны (рис. 6). Граница между бассейном устойчивого равновесия и бассейном гибели ГПС проходит между петлей, описанной ГПС на рис. 4 и более широкой петлей, описанной ГПС на рис. 5.Разместив на неизученной части фазового пространства точки, соответствующие начальным состояниям ГПС, мы можем узнать, к каким бассейнам они относятся. ГПС, лежащие на оси ординат, переходят в начало координат, к популяции P. ridibundus. Развитие всех остальных ГПС, как это ни удивительно, заканчивается их гибелью в точке, соответствующей 100% (XL).
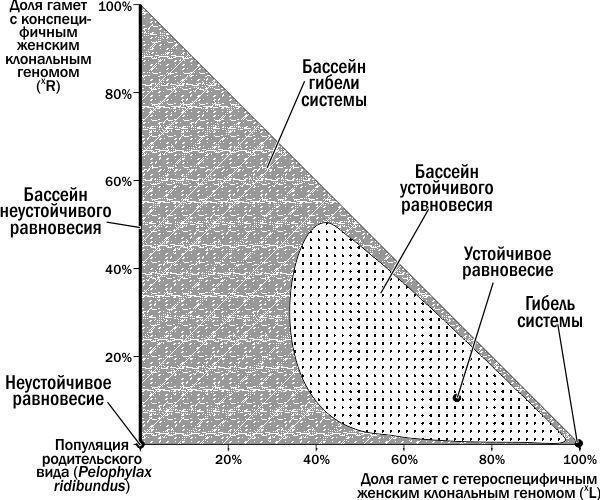
Рис. 6. Зоны (положения равновесия и их бассейны) изученного фазового пространства ГПС зеленых лягушек
На рис. 6 видно три бассейна устойчивости (один из них соответствует оси ординат) и три положения равновесия: неустойчивое, устойчивое и гибель системы. Как ни парадоксально, состояние неустойчивого равновесия тоже имеет свой бассейн. Это связано с тем, что популяция P. ridibundus, будучи неустойчива к одним возмущениям, демонстрирует устойчивость к другим.
Может ли ГПС находиться в иных состояниях устойчивости, кроме показанных на рис. 6? Выше было сказано, что А. Д. Арманд [1] выделяет в отдельные категории состояния безразличного и подвижного равновесия. Как было указано, рисунки 2, 4, 5 и 6 были получены при значениях параметров жизнеспособности, принятых в модели Batrachometrics-2010 по умолчанию. При изменении относительной жизнеспособности различных форм лягушек меняются и свойства фазового пространства. Так, при снижении относительной жизнеспособности гибридных лягушек, преимущество в их размножении компенсируется преимуществом особей родительского вида в жизнеспособности. При добавлении в популяции P. ridibundus ♀(XL)XR (т.е. в условиях, аналогичных тем, последствия которых показаны на рис. 2), ГПС переходит в состояние подвижного равновесия. Напомним, что с нашей точки зрения о подвижном равновесии можно говорить при действии на систему внешнего фактора, противостоящего внутренним процессам развития системы. В данном случае мы рассматриваем пониженную жизнеспособность гибридов как средовое, т.е. внешнее влияние на динамику ГПС.
Модель Batrachometrics-2010 является детерминистской. При одних и тех же начальных условиях она приходит к одному и тому же состоянию. Состояние безразличного равновесия достижимо при стохастичной, случайной динамике ГПС. Для моделирования таких процессов необходимо ввести в модель случайный компонент.
В следующей версии модели, Batrachometrics-2011, численность каждой группы лягушек раз в год округляется до целого числа, причем округление носит вероятностный характер (к примеру, величина 1,7 будет округлена до 1 с вероятностью 0,3, а до 2 — с вероятностью 0,7). Вследствие вероятностного округления при каждой очередной имитации с одними и теми же начальными условиями модель может приходить в различные состояния.
На рис. 7 показана динамика модели Batrachometrics-2011 в условиях пониженной жизнеспособности гибридных лягушек. С точки зрения отношения между представителями родительского вида и гибридов система переходит в состояние подвижного равновесия: доля рекомбинантных родительских геномов колеблется, несколько превышая 50%. Клональные геномы (XL) на рис. 7 представлены тремя идентичными по своей жизнеспособности и начальной численности гемиклонами. При различных имитациях с одними и теми же начальными условиями судьба этих гемиклонов оказывается разной. На рисунке показан случай, когда численность одного из них в силу случайных причин снижается до ноля, и он исчезает из ГПС.
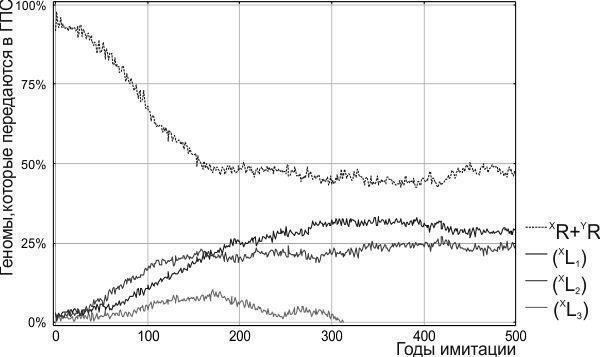
Рис. 7. Динамика модели Batrachometrics-2011 (с вероятностным округлением численностей групп особей). Пониженная жизнеспособность гибридов приводит к подвижному равновесию между особями родительского вида и гибридами. Три гетероспецифичных клональных гемиклона находятся в состоянии безразличного равновесия; в начале 4-го столетия имитации один из них исчезает из ГПС вследствие случайных причин
Типы устойчивости биосистем
Подводя итог изучению динамики ГПС зеленых лягушек с помощью имитационных моделей, можно предложить классификацию типов устойчивости биосистем. Эта классификация в основном соответствует взглядам А. Д. Арманда [1]. Типы устойчивости можно разделить на две группы (рис. 8). К первой относятся те состояния, которые могут быть «центрами» (точнее — аттракторами) бассейнов устойчивости. Это устойчивое равновесие, циклическое равновесие (примеры которого в данной работе детально не рассматривались), подвижное равновесие и состояние гибели системы. Может показаться странным, что гибель системы («негативное» состояние) оказывается стоящей в одном ряду с устойчивым равновесием (которое воспринимается как «позитивное» состояние). Роднит эти состояния то, что попав в них, система остается в них навсегда (по крайней мере, пока ее не выведет оттуда внешнее воздействие).
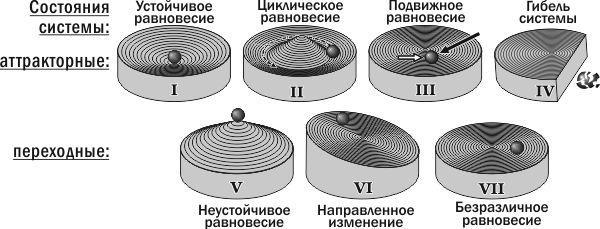
Рис. 8. Типы устойчивости биосистем (физические аналогии)
Переходные состояния образуют бассейны аттракторных состояний. Это неустойчивое равновесие, направленное изменение и безразличное равновесие. Границы между бассейнами (границы устойчивости) представлены состояниями неустойчивого равновесия.
Как показывают рассмотренные в данной статье примеры, описанные типы устойчивости не абсолютны. Их выделение зависит от того, какие параметры состояния системы отслеживаются (в какой системе координат строится фазовое пространство ее состояний). Так, достигнутое ГПС на рис. 7 состояние можно трактовать двояко. С точки зрения отношения между рекомбинантными и клональными геномами оно является состоянием подвижного равновесия, а с точки зрения отношения между различными гемиклонами его следует считать безразличным равновесием.
Выводы
Гемиклональные популяционные системы (ГПС) гибридогенных комплексов являются группой биосистем, которая была открыта совсем недавно, и их изучение только начинается. Это изучение не только предоставляет ценный материал для познания рассматриваемых групп организмов, но и расширяет понимание общих свойств биосистем.
В данной работе устойчивость биосистемы рассматривается как способность оставаться в определенной части ее фазового пространства. Эта часть фазового пространства соответствует определенному типу рассматриваемых систем. Все пространство состояний биосистемы может быть разделено на бассейны устойчивости, соответствующие определенным аттракторным состояниям. Предлагаемый подход примерен для части фазового пространства ГПС зеленых лягушек RE-типа, которая изучалась с помощью имитационного моделирования. Построенные на разной вычислительной основе имитационные модели продемонстрировали качественно сходную динамику изучаемых систем. Показано влияние на расположение и конфигурацию бассейнов устойчивости тех условий, в которых развивается рассматриваемая ГПС.
В данной работе продемонстрировано шесть типов состояний устойчивости биосистемы; седьмой тип известен из литературных источников. В целом эти типы образуют две группы: аттракторные состояния, «центры» бассейнов устойчивости, и переходные состояния, которые со временем переводят рассматриваемую систему к тем или иным аттракторным состояниям.
Библиографические ссылки
1. Арманд А. Д. Механизмы устойчивости геосистем. - М.: Наука, 1992. - 208 с.
2. Арнольд В. И. Теория катастроф. – М.: Наука, 1990. – 128 с.
3. Бигон М. Экология. Особи, популяции и сообщества: в 2-х т. Т. 2./ Бигон М., Харпер Дж., Таунсенд К. // М.:Мир, 1989. –477 с.
4. Владимирова М. В. Исследование целостных свойств биосистем на примере имитационного моделирования гибридогенных популяционных систем зеленых лягушек / Владимирова М. В., Жолткевич Г. Н., Луцик А. А., Шабанов Д. А.// Вісник Харківського нац. ун-ту. Серія «Математичне моделювання. Інформаційні технології. Автоматизовані системи управління» — 2007, № 780, C. 61–70.
5. Калитин Б. С. Качественная теория устойчивости движения динамических систем. – Минск: БГУ, 2002. – 198 с.
6. Кравченко М. А. Моделирование трансформаций гемиклональных популяционных систем зеленых лягушек (Pelophylax esculentus complex; Amphibia, Ranidae) с помощью рекуррентных разностных уравнений/ Кравченко М. А., Шабанов Д. А. // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. — 2010. – Вип.12 (№920). – С. 70–82.
7. Лада Г. А. Среднеевропейские зеленые лягушки (гибридогенный комплекс Rana esculenta): введение в проблему // Флора и фауна Черноземья. — Тамбов, 1995. — С. 88—109.
8. Ляпунов А. М.Общая задача об устойчивости движения. М.-Л.: Гостехиздат, 1950. – 472 с.
9. Математический энциклопедический словарь. – М.: Советская энциклопедия, 1988. – 847 с.
10. Одум Ю. Экология: в 2-х т. Т. 1. – М.: Мир, 1986. – 328 с.
11. Пианка Э. Эволюционная экология. – М.: Мир, 1981. – 400 с.
12. Риклефс Р. Основы общей экологии. – М.: Мир, 1979. – 424 с.
13. Стороженко В. Г. Устойчивые лесные сообщества. Теория и эксперимент. – Тула: Гриф и К., 2007 – 192 с.
14. Сукачев В. Н. Избранные труды. – Л. Наука, 1972. – Т. 1, 343 с.
15. Уиттекер Р. Сообщества и экосистемы. – М.: Прогресс, 1980. – 327 с.
16. Шабанов Д. А. Які ж зелені жаби населяють Харківську область? Термінологічний і номенклатурний аспекти проблеми / Шабанов Д. А., Коршунов О. В., Кравченко М. О. // Біологія та валеологія. — Харків: ХДПУ, 2009. — Вип. 11. — С. 164–125.
17. Шабанов Д. А. Зеленые лягушки: жизнь без правил или особый способ эволюции? / Шабанов Д. А., Литвинчук С. Н. // Природа. — 2010. — № 3. — С. 29–36.
18. Begon M. Ecology. From individuals to ecosystems. / Begon M., Townsend C. R., Harper J. L. // Blackwell Publishing, 2006. — 738 р.
19. Clements F. E. Plant succession. An analysis of the development of vegetation. – Washington: Carnegie Inst., 1916 – 242 p.
20. PlötnerJ. Die westpaläarktichen Wasserfrösche. Bielefeld: Laurenti-Verlag, 2005. – 161 S.
21. Uzzell T. M. Electrophoretic phenotypes ofRana ridibunda, Rana lessonae and their hybridogenic associate Rana esculenta/ Uzzell T. M., Berger L. // Proc. Acad. nat. Sci. Phila. — 1975. — Vol. 127. — P. 13-24.