Маро А. Н., Шабанова А. В., Шабанов Д. А. Могут ли условия развития головастиков Bufo bufo определять темпы постметаморфического роста и созревания жаб? // Вопросы герпетологии. Материалы III съезда Герпетологического общества им. А. М. Никольского. – СПб: 2008. – С. 274–280.
ISBN 978-5-98092-021-0
© Герпетологическое общество им. А. М. Никольского, 2008
© Зоологический институт Российской академии наук, 2008
МОГУТ ЛИ УСЛОВИЯ РАЗВИТИЯ ГОЛОВАСТИКОВ Bufo bufo ОПРЕДЕЛЯТЬ ТЕМПЫ ПОСТМЕТАМОРФИЧЕСКОГО РОСТА И СОЗРЕВАНИЯ ЖАБ?
А. Н. Маро, А. В. Шабанова, Д. А. Шабанов
Харьковский национальный университет им. В. Н. Каразина
CAN CONDITIONS OF THE TADPOLE Bufo bufo DEVELOPMENT DETERMINE THE RATE OF THE POST-METAMORPHIC GROWTH AND MATURING OF TOADS?
A. N. Maro, A. V. Shabanova, D. A. Shabanov
V. N. Karazin Kharkiv National University
In the course of this work we have studied some population characteristics in 9 populations (breeding groups) of Bufo bufo from Kharkov oblast' (Ukraine). For detecting the age of toads we used the skeletochronology, the size structure was established by the morphometry. Essential differences were found between populations differing by their longevity. The «old» populations, that exist more than 12 years, possess the low size variability in both sexes. By results of capture- mark-recapture method we established that the essential part of toads took part in the breeding for over 7 years. The «young» populations, which originate from an artificial introduction into unoccupied habitats, possess more variable males, including the great number of smaller and younger ones, and larger females. Possibly these differences were caused by the variation of the tadpole density in breeding ponds.
Для представителей рода Bufo, в том числе для серой жабы (Bufo bufo (Linnaeus, 1758)) характерно формирование изолированных локальных популяций (групп размножения), тесно связанных с нерестовыми водоемами. Такие группы размножения характеризуются высокой степенью уникальности. Для изучения причин такой уникальности представляет интерес изучение последствий искусственного расселения серых жаб в ранее незанятые ими местообитания. Такое расселение было выполнено в 1994 году при участии одного из авторов данной работы.
В данной работе приведены результаты морфометрического изучения 9 групп размножения серых жаб (табл. 1). Все изученные особи пойманы на нересте. Точки сбора расположены в Змиевском (№№ 1—3 в табл. 1), Харьковском (№№ 4—8) и Чугуевском (№ 9) районах Харьковской области. Возраст жаб определяли по числу ростовых слоев в кости (Смирина, 1989). На замораживающем микротоме изготовляли поперечные срезы фаланг пальцев, которые окрашивали гематоксилином по Эрлиху. Количество линий склеивания в фалангах пальцев определили для 52 особей. Кроме того, в данной работе использованы результаты многолетнего мечения (путем ампутации пальцев) и повторного отлова серых жаб, выполнявшегося в Иськовом пруду (№ 3).
Жабы, взятые из Иськова пруда (№ 3), в 1994 г. были вселены в два водоема, №№ 4 и 6. В дальнейшем жабы расселялись и формировали новые группы размножения в соседних водоемах самостоятельно. Водоем № 4 является нижним в системе из соединенных прудов, поэтому здесь расселение жаб происходило против водотока. Через 10 лет после вселения жабы лишь начали осваивать третий из этих прудов (№ 5). Из водоема № 6 расселение жаб происходило вниз по течению по системе из семи прудов (водоем № 7 является шестым из них) и произошло в первые годы после интродукции. Затем жабы через водораздел заселили пруд в соседней балке (№ 8). Общая последовательность расселения такова: 3—>4—>5 и 3—>6—>7—> 8. Полевые наблюдения за перемещением жаб во время нереста позволили утверждать (Маро, Шабанов, 2005), что расселение жаб происходит при попадании особей, идущих на нерест, в иной водоем, расположенный у них на пути. Кроме того, головастики могут быть перенесены током воды.
Сравнение жаб из давно существующих и недавно возникших популяций показало существенные отличия в их размерной структуре (табл. 1, рис. 1). Например, при сравнении распределений по размеру жаб из водоемов № 3 («материнского») и № 8 по Колмогорову — Смирнову отличия оказываются значимыми (р < 0.001 при сравнениях как по размерам самок, так и по размерам самцов). При сравнении объединенных данных из давно существующих популяций (№№ 1—3), с одной стороны, и из популяций, образованных в результате искусственного расселения (№№ 4—8), с другой, отличия также оказываются значимыми (р < 0.005 при сравнении самок и р < 0.001 при сравнении самцов). Группы размножения в недавно заселенных местообитаниях характеризуются относительно более крупными размерами самок, а также участием в нересте совсем мелких самцов, принадлежащих к размерным классам, которые не принимают участия в размножении в давно существующих популяциях. Это дало основание выдвинуть предположение, что при заселении новых местообитаний у самцов серых жаб ускоряется созревание, а у самок — рост.
Скелетохронологические исследования подтвердили это предположение (рис. 2).
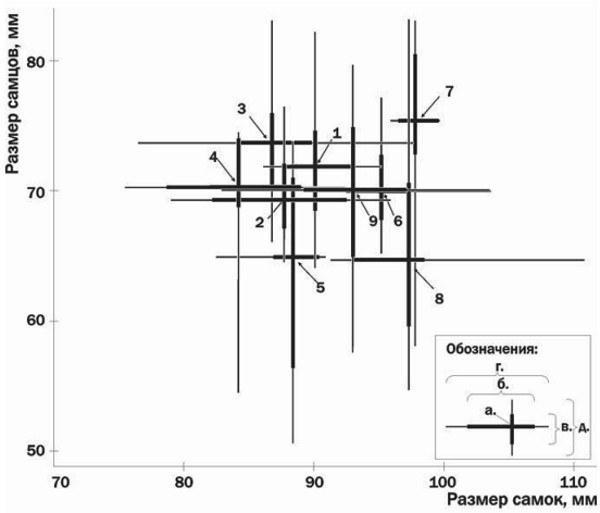
Рис.1. Размерная структура изученных выборок жаб. Характеристики каждой популяции показаны на рисунке крестом. Обозначения: а — средние значения длины тела самок и самцов; б и в (жирные линии) — диапазон изменчивости, включающий 50% особей для самок и самцов соответственно (верхний и нижний квартили); г и д (тонкие линии) — минимальные и максимальные значения размеров для самок и самцов соответственно
|
Таблица 1.Точки сбора, численность (N) и особенности размерной структуры изученных выборок серых жаб (L— средняя длина тела, s— стандартное отклонение длины тела).
|
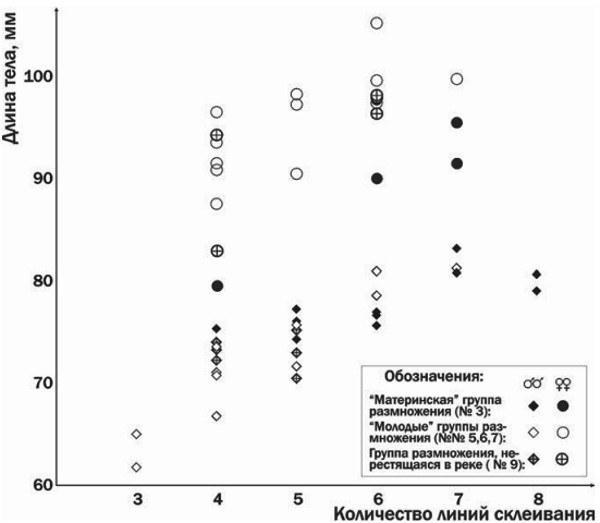
На рис. 3 видно, что самцы, участвовавшие в нересте в 2000 и 2001 году, составляли существенную часть группы размножения даже через 6 или 5 лет соответственно. Кроме того, представленные на рис. 3 результаты свидетельствуют о сокращении численности группы размножения Иськова пруда за время наблюдений. Возможно, это следствие того, что в 2000 году Иськов пруд был спущен во время нереста серых жаб (Ачкасова та ін., 2001). Следовательно, группу размножения серых жаб Иськова пруда характеризуют невысокие темпы роста в сочетании с многолетним (до 6—7 раз) участием в нересте.
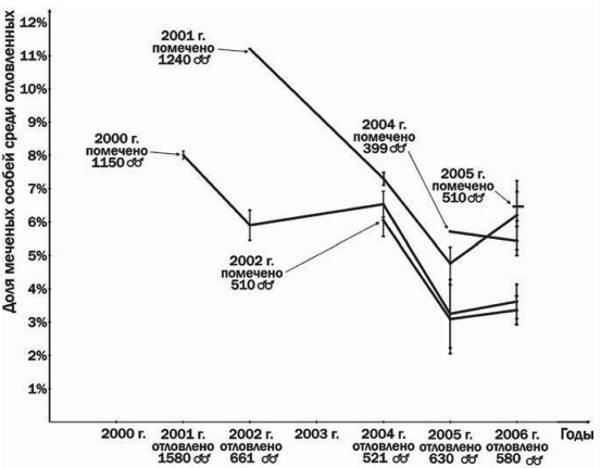
Рис.3. Результаты мечения и повторного отлова самцов серых жаб из Иськова пруда. Вертикальные отрезки отмечают максимальную и минимальную оценку возвратов, зависящую от интерпретации недостаточно хорошо различимых меток. Для каждого года указана численность отловленных и помеченных особей
Приведенные данные дают основание для предположения, что в материнской популяции самцы выходят на нерест более в позднем возрасте, но зато оказываются более конкурентоспособными. В конкурентной среде мелкие самцы будут иметь низкие шансы оплодотворить самку. Возможно, адаптивным является и более медленный рост самок в конкурентной среде, так как у амфибий и рыб до максимального возраста доживают не самые крупные и, как правило, более медленно и равномерно растущие особи (Мина, Клевезаль, 1976).
Значит, можно предположить, что развитие жаб в недавно освоенных местообитаниях способствует росту их численности, а в давно населенных — повышению их конкурентоспособности. Такое различие стратегий индивидуального развития можно сравнить с различием г- и К- стратегий по Р. Мак-Артуру и Е. Уилсону (Пианка, 1981). Следует отметить, что в данном случае речь идет не о сравнении стратегий различных видов, а о сравнении демографических стратегий популяций и онтогенезов отдельных особей одного и того же вида.
Особенности популяции серых жаб, нерестящихся в русле реки Север- ский Донец в месте с достаточно сильным течением (№ 9) заставляют предположить, что в проточной воде влияние высокой плотности головастиков на постметаморфический онтогенез жаб уменьшается. Описываемая группа размножения существует уже давно и характеризуется высокой численностью и плотностью нерестящихся жаб, но имеет характерную для «молодых» групп размножения размерную структуру: в ней сочетаются мелкие самцы и крупные самки (рис. 1). Возможно, одним из существенных факторов взаимодействия между головастиками, определяющими их будущий онтогенез, является выделение в воду тех или иных веществ, влияющих на их рост и развитие (Шварц, 1972 и другие работы).
На основании изложенного, мы выдвигаем следующую гипотезу. Скорость постметаморфического роста и возраст достижения половозрелости серых жаб определяется плотностью головастиков в водоемах, где они проходили развитие. Низкая плотность головастиков способствует развитию жаб, при котором происходит максимизация их плодовитости в течение ближайших лет. Самцы в таких популяциях принимают участие в размножении в относительно молодом возрасте и при некрупных размерах; самки отличаются высокой скоростью роста и, вероятно, относительно низкой продолжительностью жизни. Там, где плотность головастиков высока, характер постметаморфического развития жаб отражает максимизацию суммарной продуктивности в течение всей жизни в условиях конкурентной среды. В таких местообитаниях жабы медленнее развиваются и растут, но дольше живут и приходят на нерест готовыми к острой конкуренции. В частности, готовность самцов к половому отбору проявляется в достижении ими половозрелости при более крупных размерах.
Подтвердить или опровергнуть выдвинутые гипотезы могут дополнительные исследования, которые ведутся в настоящее время.
Авторы благодарят Э. М. Смирину, С. М. Ляпкова и В. Г. Ищенко за ценную критику, но ни в коем случае не возлагают на них ответственность за высказанные в статье предположения. Авторы также благодарны А. В. Коршунову, М. А. Кравченко и Г. А. Мазепе за помощь в полевых работах.
Ачкасова І. В., Дьяченко Л. А., Мякота Я. Ю., Шабанова Г. В., Шабанов Д. А., 2001. Розмірна структура популяції сірих ропух (Bufo bufo)// Біолоія та валеологія. 36. наук. праць. Харків: ХДПУ. Вип. 4. С. 41—46.
Маро А. Н., Шабанов Д. А., 2005. Механизмы расселения серой жабы (Bufo bufo (L. 1758); Amphibia, Anura) и особенности ее популяций в недавно заселенных местообитаниях // Матеріали Першої конференції Українського Герпетологічного Товариства. К.: Зоомузей ННПМ НАНУ. С. 107—110.
Мина М. В., Клевезаль Г. А.,1976. Рост животных. М.: Наука. 291 с.
Пианка Э., 1981. Эволюционная экология. М.: Мир. 400 с.
Смирина Э.М, 1983. Прижизненное определение возраста и ретроспективная оценка размеров тела серой жабы (Bufo bufo)// Зоол. журн. Т. 62. Вып. 3. С. 437—444.
Шварц С. С.,1972. Метаболическая регуляция роста и развития животных на популяционном и организменном уровнях // Известия АН. СССР. Сер. биол. № 6. С. 142—151