|
1.2. МЕТОДОЛОГИЯ ФИЛОГЕНЕТИКИ,
ТАКСОНОМИИ И НОМЕНКЛАТУРЫ
1.2.1. ФИЛОГЕНЕТИКА 1.2.1.1. Анализ групп 1.2.1.2. Анализ признаков 1.2.1.2.1. Анализ различий
1.2.1.2.2. Анализ сходства
1.2.1.3. Компьютерный кладизм
|
А.П.Расницын. Теоретические основы эволюционной биологии // В.В.Жерихин, А.Г.Пономаренко, А.П.Расницын. Введение в палеоэнтомологию. М.: КМК. 2008. 371 с
1.2.2. ТАКСОНОМИЯ
1.2.2.1. Кладизм
1.2.2.2. Фенетика
1.2.2.3. Филетика |
1.2.2. ТАКСОНОМИЯ
Таксономическая классификация организует биологическое разнообразие таким образом, чтобы облегчить наше взаимодействие с ним. Существует много форм классификации, созданных для определенных целей и определенных категорий пользователей (например, система жизненных форм, или система вредителей по признакам причиняемого ими вреда). Они не представляют значительных теоретических проблем, и здесь речь пойдет только о той системе, которая используется в качестве междисциплинарного языка. Будем называть ее общей системой организмов.
Таксоны общей системы, чтобы выполнять свои функции, должны быть осмысленными объединениями с точки зрения максимально широкого круга пользователей, как птица представляет вполне определенное понятие не только для орнитолога, но и для охотника, повара и художника. Чтобы соответствовать такой цели, таксоны общей системы должны быть максимально однородными внутри себя и максимально различными между собой по максимально широкому кругу признаков. Это свойство позволяет системе выполнять еще две важнейшие функции. Во-первых, она свертывает информацию о живых организмах в доступной форме. Вспомним, что каждый из нас знает, скажем, о таракане, включая информацию о признаках старших таксонов. Во-вторых, она позволяет предсказывать свойства недостаточно изученных таксонов. Лишь ничтожная часть видов насекомых изучена сколько-нибудь детально, но мы не сомневаемся, что они состоят из эвкариотических клеток соответствующей структуры и функции, и с достаточной уверенностью описываем многие детали биологии даже вымерших видов. Например, находка в нижнемеловых отложениях Забайкалья неполного переднего крыла, определенного как крыло хлебного пилильщика (Cephidae), послужило основанием для предсказания, что уже в начале мела существовали покрытосемянные растения, на которых только и развиваются современные хлебные пилильщики (Расницын, 1969). И покрытосеменные действительно были найдены в той же Байсе и в тот самый год, когда предсказание было опубликовано (Вахрамеев, Котова, 1977).
В систематике последних десятилетий выделяются три основных направления (табл. 3): фенетика, кладизм и еще одно, часто неудачно именуемое «эволюционной систематикой» — в отличие от «филогенетической систематики», первоначального и столь же неудачного названия кладизма (действительно, чем отличаются термины «эволюционный» и «филогенетический» в данном контексте?). Для этого направления, которое пытается выявить и сформулировать принципы традиционной систематики, был предложен термин «филетика» (Пономаренко, Расницын, 1971).
Различия между тремя направлениями гораздо глубже, чем можно было бы предположить: им соответствуют совершенно различные представления о структуре и происхождении биологического разнообразия.
Одно из перечисленных направлений совместимо с синтетической, а более точно — популяционно-генетической теорией эволюции. Последняя, как мы видели, представляет эволюционный процесс как динамику аллелей в популяциях, контролируемую отбором и стохастическими процессами. Отбор формирует состав популяций и свойства организмов, и делает он это с точностью, ограниченной лишь стохастическими факторами и обменом генами, который унифицирует структуру популяций на уровне, отвечающем не локальным, а усредненным характеристикам отбора.
Организация живых существ предстает здесь набором признаков, свободно тасуемых отбором, а эволюционный процесс — равномерным в той мере, в какой равномерны определяемые средой вариации отбора. Эту равномерность нарушают лишь дивергенции и вымирания, каковые только и могут быть использованы для построения естественной системы организмов. То есть естественная система, отвечающая популяционно-генетической теории эволюции, может быть только строго генеалогической, с границами таксонов, проводимыми по точкам дивергенции (узлам кладограммы). Таксоны выделяются в соответствии с реконструированными фактами их истории, а не их нынешними свойствами, которые важны только как исторические свидетельства. Это точное описание кладистической системы: строго иерархической и с исключением парафилетических, т.е. предковых таксонов (табл. 1). Кладизм сводит систему группы к ее генеалогической истории, фенетика к доступным для анализа признакам сходства, а традиционная систематика (в зеркале филетики) пытается использовать и то, и другое, причем не произвольно, а упорядоченным образом (табл. 3).
Таблица 3. Три таксономии
|
Тип |
Эволюционная основа |
Задающий процесс |
Рабочий критерий |
Сходство |
Кровное родство |
|
Кладизм |
синтетическая |
акт дивергенции |
синапоморфия |
игнорируется |
единственная основа системы |
|
Фенетика |
эпигенетическая |
дискретность биоразнообразия |
гиатус |
единственная основа системы |
игнорируется |
|
Филетика |
эпигенетическая |
дискретность биоразнообразия |
гиатус |
задающий критерий |
контролирующий критерий |
В отличие от синтетической, эпигенетическая теория эволюции, как мы тоже знаем, видит эволюционный процесс прежде всего как процесс эволюционного преобразования онтогенеза и в частности устоявшихся (отобранных в чреде поколений) эпигенетических процессов и их стабилизированных ансамблей — креодов. Напротив, мутации и аллели выступают здесь в роли переключателей между альтернативными траекториями развития, а не в качестве создателей структур, свойств и признаков.
Глубокая взаимозависимость между онтогенетическими процессами и, соответственно, между свойствами и признаками делает оптимизацию целого организма необычайно трудной задачей, продуктом напряженного компромисса. Однажды достигнутый успешный компромисс оказывается прочно стабилизированным (забуференным), поскольку удачная структура организма с большим трудом поддается изменению за пределы нормальной внутривидовой изменчивости (которая сама уже стабилизирована отбором предыдущих поколений). Поэтому эволюция (успешный выход за пределы стабилизированного эпигенотипа) затруднена, мало предсказуема и более или менее скачкообразна, а возникающее в результате биоразнообразие более или менее дискретно. Дискретность эксплуатируется как фенетикой, так и традиционной систематикой, которые строят свои системы путем выявления и прослеживания гиатусов — разрывов непрерывности биоразнообразия. Но обо всем по порядку.
1.2.2.1. Кладизм
Глубинное понимание кладизма разными его последователями отличается столь сильно (ср. Hennig, 1966а и Platnick, 1979), что предлагалось, например, использовать старое название «филогенетическая систематика» для версии Геннига, а кладизм оставить для так называемого трансформированного кладизма (de Queiroz, Donoghue, 1990a). Это предложение не получило большой поддержки, и я сохраняю здесь широкое понимание кладизма. Однако центральное утверждение кладизма, что «фенетическая классификация строится путем группирования по общему сходству, тогда как филогенетическая — по предполагаемым синапоморфиям» (Farris, 1979: 478), остается общепризнанным. Обосновывая его, В. Генниг (Hennig, 1966а) писал, что таксоны, обозначенные синапоморфиями, образуют систему, изоморфную филогении и потому наиболее эффективную. Отсюда и название системы — филогенетическая. Но есть и другая точка зрения, что, во-первых, само устройство (картина, pattern) природы таково, что система организмов может быть отображена ветвящейся диаграммой (иерархической классификацией). Во-вторых, эта картина (pattern) может быть отображена в результате изучения признаков и поиска иерархического (internested) набора синапоморфий. В-третьих, наши знания об эволюции, как и о системе, выводятся из того же иерархически организованного набора (паттерна) синапоморфий (Platnick, 1979: 538). Таким образом, паттерн признается первичным, а филогения — вторичной. Такой подход ставит вопрос: кто привносит паттерн в природу? каков его генерирующий процесс, если это не эволюция? Пока на этот вопрос не предлагается ответа, я не вижу возможности отдельного анализа паттерн-кладизма (трансформированного кладизма) и возвращусь к его классической версии.
Первый возникающий вопрос касается того, какой именно смысл вкладывается в центральное положение кладизма (синапоморфия как единственное обоснование таксона). Синапоморфия обычно определяется как состояние признака, приобретенное предковым видом группы и потому свойственное всем ее членам и только им. Это определение неточно, поскольку любое приобретенное состояние признака затем может быть утрачено, что считается невозможным для апоморфии (змеи утратили ноги, но не четырехногость как синапоморфию тетрапод). Апоморфия — не состояние признака, а факт, что такое состояние однажды было приобретено (факт истории группы).
Специального рассмотрения заслуживает вопрос о группах, лишенных апоморфий. Такие группы именуются парафилетическими или полифилетическими в зависимости от того, выделены ли они соответственно по симплезиоморфии или гомоплазии. Кладистическое решение состоит в разбиении таких групп на меньшие таким образом, чтобы все они стали монофилетическими (точнее, голофилетическими, см. ниже), т.е. были охарактеризованы апоморфиями. При этом, однако, возникает проблема парафилетических таксонов, которые не удается расчленить на голофилетические составляющие (метатаксоны по de Queiroz, Donoghue, 1990b). Таковы многие вымершие и некоторые современные группы, не все необходимые признаки которых могут быть изучены. В большинстве случаев эту проблему можно было бы посчитать временной и списать ее на неполноту знаний, которая будет преодолена. Однако это убежище неведения не спасает хотя бы потому, что параллельная эволюция, как мы видели, не оставляет нам надежды на расшифровку кладограммы во всех ее деталях.
Еще более существенно, что обязательное разделение парафилетического таксона на голофилетические подгруппы неизбежно приведет нас к предковому виду, который является парафилетическим таксоном по определению и который невозможно разделить на голофилетические составляющие, так как это один вид. Некоторые исследователи и здесь прибегают к убежищу неведения, утверждая. что предковый характер вида никогда не может быть верифицирован (Nelson, 1973a). Однако, как показал Поппер, научные гипотезы в принципе не могут и не должны верифицироваться. С другой стороны, если каждая ветвь филогенетического древа имеет в основании предковый вид, то простой расчет показывает, что таковые должны составлять половину всех когда-либо существовавших видов. Действительно, в случае строгой дихотомии ветвления и при отсутствии филетического видообразования число предковых видов равно числу терминальных видов минус единица. Множественное ветвление (почкование, полихотомия) несколько уменьшает долю предковых видов, филетическая эволюция увеличивает их число, но в первом приближении приходится согласиться, что половина всех описанных ископаемых видов (и, вероятно, немалое число современных) являются непосредственными предками других видов.
Вид, предковый более чем для одного таксона, представляет непреодолимую теоретическую проблему для кладизма, если только не смягчить его центральное утверждение (таксон определяется исключительно через апоморфию). Этот необходимый шаг был сделан еще Геннигом, который считал, что апоморфия узаконивает таксон не по определению, а лишь как следствие из другого, более жесткого (базового) утверждения об изоморфии между системой и филогенезом. «Время существования вида определяется двумя событиями видообразования: тем, которому этот вид обязан своим возникновением как независимое репродуктивное сообщество, и тем, которое разделяет его на два или более таких сообществ» (Hennig, 1966: 66). Такой путь рассуждений делает дивергенцию единственным процессом, создающим таксоны, а апоморфия оказывается просто маркером дивергенции и, как следствие, таксона. Другим следствием будет исчезновение предкового вида в результате дивергенции — даже если один из продуктов дивергенции не обнаруживает ни малейших отличий от предкового вида (лишен апоморфий). Это утверждение выглядит несколько странным, но по крайней мере оно последовательно.
Таким образом, в кладистической парадигме некоторые (возможно, многие) виды не имеют отличий от своих предков. Тогда у нас нет даже теоретической возможности точно восстановить топологию кладограммы: возможно лишь некоторое приближение к ней. Данное обстоятельство делает преимущества подхода не столь очевидными и открывает другие возможности, например, инвертировать центральное кладистическое утверждение и считать кладистическую систему в конечном счете определяемой последовательностью апоморфий. Тогда события дивергенции можно рассматривать как орудие определения этой последовательности. Такой подход был использован Д. Бразерсом (Brothers, 1975) в его классическом исследовании филогении и систематики жалоносных перепончатокрылых. Сходную позицию, видимо, занимает Г.Дж. Нельсон, утверждая, что «происхождение без изменения ... не является достаточным объяснением таксона» (Nelson, 1989: 280-281).
Как дивергенция не всегда сопровождается апоморфным изменением всех ее продуктов, так и приобретение апоморфии в случае филетической эволюции оказывается не обязательно связанным с дивергенцией. Следовательно, установление точной последовательности апоморфий недостижимо даже в теории. Таким образом, два подхода по меньшей мере сравнимы по их значению для филогенетического исследования — не в том смысле, что они обязательно равноценны в этом отношении, а в том, что их достоинства и недостатки равно заслуживают специального изучения. Например, в версии Бразерса апоморфии вряд ли доступны не только точному секвенированию (выстраиванию в строгую последовательность), но и простому подсчету. В большинстве случаев апоморфии слишком сложны, чтобы можно было представить их возникновение за один шаг (в течение одного события видообразования). Но последовательности элементарных шагов приобретения сложной апоморфии должна соответствовать иерархическая система таксонов, которые в принципе неразличимы.
Другие, более серьезные проблемы кладизма будем обсуждать в сравнении с достоинствами и недостатками фенетики и филетики после их более подробного рассмотрения.
Итак, кладизм утверждает, что система организмов должна быть строго генеалогической, т.е. максимально точно отражать кровное родство организмов. Для этого таксоны делятся по узлам дивергенции с помощью синапоморфий, т.е. изменений, возникших у предка и выявленных у его потомков. Сходство интересно только как свидетельство генеалогии, а само по себе несущественно. Кладистический таксон не может породить другой таксон: в кладистическом мире таксонов-предков и таксонов-потомков не существует. Конечно, кроме видов, но поскольку идентифицировать предковый вид по мнению кладистов невозможно, предок — это не реальное существо или реальный вид, а просто коллекция примитивных признаков (плезиоморфий), обнаруженных у его потомков. В этом действительно есть резон, поскольку широкое распространение параллелизмов делает разговор о предковом виде в большинстве случаев беспредметным, но оно же делает иллюзорным и самое главное утверждение кладизма о единичном событии дивергенции (или, в другой его версии, о приобретении единичной апоморфии) как о единственном событии, дающем жизнь таксону.
1.2.2.2. Фенетика
В отличие от кладизма, фенетика и традиционная систематика (или, в рафинированном виде, филетика) исходят из целостности живых систем и, соответственно, из дискретности биоразнообразия и заняты поисками гиатусов. Поэтому фундаментальная концепция должна включать процесс, генерирующий такую дискретность. Первоначально дискретность легко объяснял креационизм, сейчас на эту роль реально претендует, по-видимому, только эпигенетическая теория эволюции (см. выше).
Из трех конкурирующих подходов фенетика наиболее прямолинейна в своих целях и средствах. Поскольку дискретность биоразнообразия описывается в терминах сходства, а не родства, то и задачу систематика по мнению фенетиков следует формулировать в тех же понятиях, а именно: система должна в явном виде строиться в терминах сходства. Избежать субъективности здесь можно с помощью исчисления сходства. Методы расчета коллективно именуются таксометрией или чаще, но менее удачно, нумерической таксономией. К сожалению, эти методы не были развиты до заявленного уровня — на мой взгляд не из-за того, что этот уровень недостижим, а потому что фенетика была вытеснена кладизмом до того, как она выполнила свою полезную работу. Действительно, методы таксометрии многочисленны и разнообразны, выбор метода, адекватного задаче, не прост и на нынешнем уровне едва ли возможен без изрядной доли субъективности. Впрочем, это проблема любого подхода, и кладизм здесь не исключение (см., например, Mickevich, 1978): выявить оптимальную область применения конкретного метода, его сильные и слабые стороны можно лишь ценой изрядных усилий. Прежде чем уступить поле боя кладизму, фенетика значительно, но недостаточно продвинулась в этом направлении.
Однако коренная проблема фенетики, на мой взгляд, иная. Сходство, с которым она работает, это не совсем то сходство, которое требует система организмов. Фенетики работают с изученными признаками, число которых должно быть разумно велико, но не более того. Система же, чтобы оптимально выполнять свои функции, должна опираться на сходства и различия по всем признакам, изученным и не изученным, включая и те, что, возможно, не будут изучены никогда. Конечно, такое сходство недоступно прямому изучению, но оно достаточно важно, чтобы попытаться оценить его косвенно. Именно эту цель и преследует филетика.
1.2.2.3. Филетика
Филетика, как и кладизм, исходит из того, что организация живых существ определяется не только и даже не столько условиями их существования, сколько их историей. Сходство истории (кровное родство) во многом предопределяет и сходство организации. Разница в том, что методология кладизма игнорирует все, кроме родства, фенетика игнорирует родство, а филетика пытается учесть и родство, и сходство.
Для того, чтобы филетический таксон одновременно отражал и сходство, и родственные отношения, он должен быть, во-первых, фенетически (по сходству) максимально однороден и максимально отличен от других таксонов, во-вторых, монофилетичен. Монофилия понимается в широком смысле по сравнению с кладизмом, т.е. включая голофилию (монофилию в кладистическом смысле, см. Ashlock, 1971) и парафилию. Филогенетическая квалификация таксонов определяется линиями происхождения, пересекающими границы таксона. Если его нижнюю границу пересекает единственная линия (один корень), таксон именуется монофилетическим (в широком смысле), если их больше — полифилетическим. Более точной, впрочем, будет другая формулировка: таксон следует считать монофилетическим, пока и поскольку нет надежных свидетельств его полифилии (презумпция монофилии таксона).
Кладисты переопределили понятие монофилии, имея в виду не только происхождение (корни) таксона, но и его дальнейшую судьбу. Если верхняя граница монофилетического (в нашем смысле) таксона не пересечена филетическими линиями, т.е. таксон не имеет явных потомков, мы его именуем голофилетическим, а кладисты — монофилетическим. Если же его верхняя граница пересечена (потомки есть), и для филетистов, и для кладистов это парафилетический таксон. Кладисты именуют голофилетический таксон монофилетическим, поскольку с их точки зрения парафилетический таксон точно так же не имеет права на существование, как и полифилетический. Они не нуждаются в термине, объединяющем голофилетический и парафилетический таксоны: для них такое объединение бессмысленно. А для филетики такой термин совершенно необходим, поскольку там обе эти формы таксона (голо- и парафилетические) признаны и равноправны. Именно таким, филетическим, было первоначальное понимание термина монофилия, оно же принято и здесь.
Существует еще более широкое понимание монофилии — как происхождения естественного таксона несколькими корнями от общего предка в недрах предкового таксона (плюрифилия, пахифилия, широкая монофилия). Именно так сейчас представляют происхождение млекопитающих, рептилий и многих других крупных групп: отсюда и термины маммализация, артроподизация, ангиоспермизация и т. п. Широкая монофилия не только подпадает под определение полифилии в кладистической формулировке, но и вообще исчерпывает реальное содержание этого термина. Согласно определению, за пределами широкой монофилии остается только множественное происхождение таксона, естественность которого отрицается (мы еще будем иметь дело с ними в разделе 1.2.3.3). Но для искусственного таксона множественность его происхождения тривиальна, и вряд ли кому-то придет в голову это обсуждать специально. Содержать отдельный термин для такого случая мне кажется излишним, так что я буду называть полифилией все случаи множественного происхождения таксона, как это и сформулировано выше.
Таким образом, оказывается, что многие (если не все) хорошо изученные таксоны являются полифилетическими в указанном смысле. В то же время трудно усомниться в естественности рептилий, млекопитающих, членистоногих, покрытосеменных и многих других таксонов, для которых более или менее убедительно показано происхождение более чем одним корнем от предкового таксона. Таким образом, полифилия может оказаться нормой в таксономии. И тем не менее я считаю, что методологически правильнее исходить из презумпции монофилии. Тому я вижу две причины: во-первых, на нынешнем весьма низком уровне изученности большинства таксонов обнаружение множественности происхождения конкретной группы скорее будет фальсифицировать гипотезу ее естественности, чем презумпцию монофилии. Если же группа действительно естественна, то дополнительное исследование, спровоцированное обнаружением полифилии (см. ниже), только подтвердит ее естественность и в конце концов заставит признать презумпцию монофилии несправедливой в данном конкретном случае.
Существует и другая причина, почему отказ от презумпции монофилии (в принятом здесь понимании, исключающем широкую монофилию) представляется методологически опасным. Это означало бы отказ от использования филогенеза для контроля естественности таксона (см. ниже) и, следовательно, переход на позиции фенетики. Такое решение кажется по меньшей мере преждевременным.
Вернемся к филетической процедуре. Филетический таксон был определен как монофилетический континуум (Пономаренко, Расницын, 1971). Определение монофилии дано выше, континуум же понимается как непрерывная цепь, ветвящаяся или нет, построенная из подчиненных таксонов (монофилетических континуумов меньшего объема) таким образом, что каждый из них фенетически ближе к любому из его непосредственных соседей по цепи, чем к членам любых других таксонов (рис. 6). Таким образом, континуум отделен гиатусом (перерывом непрерывности сходства) от всех других континуумов.
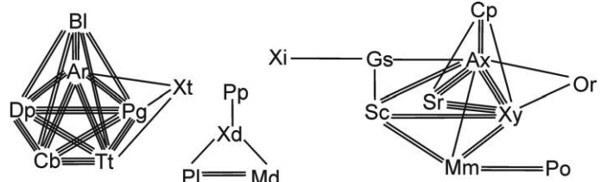
Рис. 6. Фенетическая структура низших перепончатокрылых насекомых на уровне семейства, демонстрирующая континуумы нескольких иерархических уровней (из Расницына, 19726). Линиями изображены уровни сходства: одинарной линией — низший уровень из изображенных, тройной — наивысший. Сокращения: Аг — Аrgidае, Ах — Аnахуеlidае, В1 — Blasticotomidae, Сb — Cimbicidае, Ср — Серhidае, Dр — Diprionidае, Gs — Gidasiricidае, Мd — Меgаlоdontesidае, Mm — Myrmiciisае, Оr — Orussidае, Рg — Pergidae, Рl — Раmphiliidае, Ро — Раrоryssidaе, Рр — Раrараmphiliidае (ныне в Sepulcidае), Sc — Sepulcidае, Sr — Siricidае, Tt — Tenthredinidае, Хd — Хyelydidае, XI — Хуеlidае, Xt — Xyelotomidae, Ху — Хiphydriidае.
Строить систему, основанную на двух разных критериях (в данном случае это родство и сходство), рискованно тем, что допускает произвол в их применении — если только сферы компетентности двух критериев четко не разведены. Кладизм и фенетика решают проблему предельно просто — отказываясь использовать тот или иной из двух критериев: кладизм игнорирует сходство, фенетика — родство. Филетика действует тоньше: она использует оба критерия, но по-разному. Сходство используется как критерий, задающий систему, родство — как критерий, контролирующий качество системы. Таксон выделяется прослеживанием гиатусов (разрывов в сходстве), а затем проверяется на единство происхождения.
Итак, филетика требует, чтобы члены ее таксона были достаточно сходны, чтобы удовлетворять критерию континуума, и при этом не обнаруживали бы признаков полифилии (происхождения от более чем одной линии предков). Если континуум оказывается полифилетичным, критерии вступают в конфликт. В этом случае систематик едва ли станет отбрасывать все, что было сделано, и начинать работу сначала. Его естественной реакцией будет попытка фальсификации этого результата путем дополнительного изучения соответствующей группы. Попытка обычно оказывается успешной в том или ином смысле. Например, более широкий анализ может обнаружить, что сходство полифилетических групп поверхностное. Отряд сетчатокрылых насекомых (Neuroptera) долгое время включал группы, близкое родство которых впоследствии не подтвердилось. Так, еще Шарп (1910) объединял под этим именем, помимо сетчатокрылообразных в современном понимании, также эмбий (Embioptera), термитов (Isoptera), сеноедов (Psocoptera), пухоедов (Mallophaga), стрекоз (Odonata), веснянок (Plecoptera), поденок (Ephemeroptera), скорпионниц (Mecoptera), и ручейников (Trichoptera). Похожей была судьба млекопитающих отряда толстокожих (Pachydermata): таксон использовался в XVIII и XIX веках для объединения слонов, тапиров, носорогов, бегемотов и свиней, но затем был отвергнут в пользу выделения более естественных (монофилетических) отрядов.
В других случаях удалось фальсифицировать гипотезу полифилии, как в случае с зайцеобразными. Они были выделены в свое время в особый отряд Lagomorpha, будто бы лишь конвергентно сходный с грызунами (Gidley, 1912). Хотя статус отряда за зайцеобразными сохранился, по современным представлениям они образуют с грызунами монофилетическую группу Glires (Wilson, 1989).
Вполне возможен случай, когда обе гипотезы (сходства и полифилии) окажутся равно правдоподобными. Скорее всего, это будет означать, что наша попытка была недостаточной, и нужно попытаться заново проанализировать ситуацию и устранить конфликт. Но нельзя исключить, что мы действительно добрались до реальной множественности корней естественного таксона, о которой мы уже неоднократно говорили. Как отличить одну ситуацию от другой, у меня рецепта нет, но на всякий случай лучше еще раз попытаться проанализировать систему и филогенез группы на более широком материале.
Существуют и совершенно очевидные случаи, когда традиционная систематика сохраняет несомненно полифилетические таксоны, которые не могут быть без значительных потерь заменены монофилетическими. Я имею в виду различные паратаксоны в таких областях, как палеонтология, паразитология, микология, а также многие таксоны прокариот (подробнее см. раздел 1.2.3). Существование паратаксонов еще раз подтверждает, что традиционная систематика в конечном счете ориентируется на сходство, а родство использует лишь как способ косвенным образом оценить тотальное сходство (в том числе по еще не изученным признакам), которое прямой оценке недоступно. Следовательно, родство здесь используется приблизительно так же, как в генниговой версии кладизма апоморфия используется для установления дивергенций, а в бразерсовой версии дивергенции служат для определения последовательности апоморфий.
Таким образом, все рассмотренные подходы одинаково последовательны (монистичны, не эклектичны): геннигов кладизм в конечном счете ориентирован на последовательность актов дивергенции, бразерсов — на последовательность апоморфий, фенетика — на наличное (доступное непосредственному изучению) сходство, филетика — на тотальное сходство (недоступное прямому наблюдению, но косвенно оцениваемое с помощью родства).
Теперь мы можем вернуться к вопросу о сравнительных достоинствах и недостатках кладизма и филетики. На мой взгляд, важнейшим достоинством филетики является ее конечная опора на сходство, т.е. на признаки как таковые. Она может удовлетвориться сходством в наличных признаках, если сколько-нибудь надежные свидетельства родства отсутствуют, и будет при этом столь же эмпирична, как фенетика. Фактически это и будет фенетика. Но она же будет использовать все (и только!) достаточно надежные филогенетические данные, чтобы повысить надежность создаваемой ею системы, не делая ее при этом слишком гипотетичной. В противоположность филетике, кладистическая система и непосредственно, и в конечном счете опирается на родство, т.е. на интерпретацию признаков (на предположения относительно эволюционной последовательности состояний признаков). В результате кладистическая система значительно более гипотетична, чем филетическая.
Другая сторона сказанного заключается в том, что филетика «всеядна», а кладизм «привередлив». Кладизм предъявляет гораздо более жесткие требования к используемым им признакам, чем филетика (и фенетика), потому что признаки, не поддающиеся надежной поляризации, бесполезны. Это, правда, касается только геннигова кладизма (ручного, или «кладизма пера и бумаги»). В компьютерном кладизме можно использовать любые признаки, но если программа не сможет уверенно реконструировать последовательность изменений некоторого признака (а она вряд ли сможет это сделать много лучше, чем хороший систематик), то этот признак будет лишним в расчетах.
Более того: признаки, хорошо работающие в какой-то части исследуемой совокупности, но обнаруживающие многочисленные параллельные изменения (гомоплазии) в других ее частях, как мы видели, девальвируются и учитываются лишь в малой степени — даже там, где в действительности они филогенетически информативны. Менее изученные таксоны, например, вымершие, охарактеризованы меньшим числом признаков и поэтому часто не находят точного места в компьютерных кладограммах, даже если сохраняют филогенетически важные признаки. В результате кладисты вынуждены объявлять паратаксонами («плезионами») многие группы, рассматриваемые филетикой как нормальные (ортотаксоны), но известные недостаточно для того, чтобы точно охарактеризовать их родственные отношения с лучше изученными таксонами. Естественно, это снижает таксономическую эффективность кладизма.
Филетическая система, учитывающая как сходство, так и родство, более информативна, чем другие. Дж.С. Фаррис (Farris, 1979) в качестве возражения напоминает, что кладистическая система в ее наиболее полной форме указывает происхождение всех признаков и поэтому более информативна, чем фенетическая (и, следовательно, филетическая). Этот вывод не бесспорен, поскольку исчерпывающая полнота системы недостижима. Действительно, чем больше информации мы набираем о поляризации известных признаков, тем больше обнаруживается новых признаков, характер эволюции которых еще не известен и которые поэтому не могут быть использованы в «ручном» кладистическом анализе. В результате приходится работать с неполным набором признаков по сравнению с филетикой, и в этом смысле система несет меньший объем информации. Компьютерный кладизм может использовать все доступные признаки, но поляризацию трансформационных серий он производит автоматически, исходя просто из распределения состояний признаков (по методу внешней группы), т.е. из весьма ограниченной информации. Все прочие знания об истории признаков, широко используемая филетикой и ручным кладизмом, здесь игнорируется, так что общие потери информации велики.
Фактически любой из сравниваемых таксономических методов использует только часть доступной информации, причем эти части не слишком сильно перекрываются. Компьютерный кладизм использует обширное поле данных, но весьма избирательно и поверхностно. Фенетика действует сходным образом, но не практикует избирательности. Ручной кладизм использует ограниченный «элитный» материал, отобранный из неограниченно широкой совокупности. Филетика также «просеивает» неограниченный объем исходных данных, но она более свободна, чем «ручной» кладизм, в его сортировке, и чем фенетика и компьютерный кладизм, в его интерпретации. Поэтому она способна к извлечению более обширной и разнообразной информации, чем другие подходы.
Существует и другой аспект проблемы. Из сказанного выше видно, что информация заключена не в системе самой по себе, а в ее описании или, скорее, в том, что лежит в ее основании. Система как таковая содержит только весьма ограниченную информацию о топологии соответствующей дендрограммы и о длине ее междоузлий, интерпретированной в таксономических рангах. Так, симметричная дихотомическая дендрограмма о четырех конечных таксонах и без дополнительной информации об их рангах содержит три бита информации и не больше. Детализация ранжирования таксонов обогащает информационное содержание системы, и в этом плане наиболее богаты оригинальные системы Геннига с их невероятным числом нумерованных рангов, исчерпывающе отражающих предполагаемую последовательность актов дивергенции. Однако этот подход был признан непрактичным, и он фактически оставлен в пользу традиционной линнеевой иерархии рангов (Wiley, 1979). Такая форма ранжирования содержит ограниченную кладистическую информацию и использует ее произвольным образом (привязка ранга к одной из многих последовательных дивергенций произвольна), делая тем самым послегенниговый кладизм эклектичным. Искусственные правила типа нельсонового phyletic sequencing (Nelson, 1973b; Cracraft, 1974) мало помогают. Мы можем договориться, что гребенчатую кладограмму следует преобразовать в группу таксонов одного ранга, перечисляемых в порядке их дивергенции, начиная с отошедшего первым, но в последней паре порядок неизбежно будет произвольным. И мы не сможем отличить такой список от другого, построенного произвольно или основанного на более сложной кладограмме, не строго гребенчатой или/и включающей политомию.
Утверждается, что фенетика и филетика в отличие от кладизма используют таксоны, охарактеризованные отсутствием признака (Platnick, 1979). Имеется в виду парафилетический таксон, характеризующиеся отсутствием апоморфий. Выше, однако, было показано, что апоморфия не есть признак или состояние признака. Апоморфия — это гипотеза относительно истории признака, так что ее отсутствие и отсутствие признака — вещи разные. Ящерицы лишены апоморфий, отделяющих их от змей, но у них есть признак — ноги.
Заслуживает обсуждения также проблема симбиотических и гибридных таксонов, важная в силу хотя бы того, что они слишком обычны, чтобы их игнорировать как нечто экзотическое. Первую категорию представляет не менее чем сам таксон эвкариот, а гибриды очень характерны, например, для многих таксонов растений. Согласно обзору Н.Н. Цвелева (1993), в семействе злаков (Poaceae) триба Triticeae включает 500 видов, из которых 300 кариотипически определенно представляют собой межродовые гибриды, которыми, в частности, составлены самые большие роды. Утверждается, что то же самое свойственно и многим другим таксонам высших растений. Симбиотический и гибридный таксоны полифилетичны (или составляют часть полифилетического таксона) по определению, поскольку их граница (или граница старшего таксона) пересечена более чем одной предковой линией, что категорически нарушает принципы кладизма. Тем не менее, такие таксоны принципиально не могут быть расчленены на голофилетические составляющие и поэтому должны быть исключены из компетенции кладизма, с чем вряд ли кто-либо согласится.
Существуют и другие соображения, имеющие отношение к сравнению конкурирующих таксономических концепций. Однако и сказанного достаточно для заключения, что каждой из них свойственны свои достоинства и недостатки, которые, впрочем, не всегда уравновешивают друг друга.
Кладизм сводит систему группы к ее генеалогической истории, фенетика к доступным для анализа признакам сходства, а традиционная систематика (в зеркале филетики) пытается использовать и то, и другое, причем не произвольно, а упорядоченным образом. При этом, если сколько-нибудь надежные свидетельства родства отсутствуют, традиционная систематика удовлетворится просто сходством в наличных признаках, и тогда это будет фенетика. Но когда возможно, она же будет использовать все доступные филогенетические данные, чтобы повысить надежность создаваемой ею системы.
Фенетика сугубо эмпирична, а кладизм, напротив, высоко гипотетичен, так как и непосредственно, и в конечном счете опирается на родство, т. е. на интерпретацию сходства. Филетика же свободна занимать любую позицию в этом спектре в зависимости от свойств материала, доступного в каждом отдельном случае. Филетика «всеядна» и может работать с любым доступным материалом, используя при необходимости любые фенетические и кладистические методы. Напротив, кладизм «привередлив» и предъявляет гораздо более жесткие требования к используемым им признакам. Поэтому так часто кладистическая система оказывается либо неустойчивой, когда минимальные изменения параметров расчета существенно меняют форму кладограммы, либо не интересной («не разрешенной», с не выясненными отношениями многих таксонов). По этой же причине кладисты вынуждены объявлять паратаксонами («плезионами») многие группы, которые для филетики вполне нормальны.
Тем не менее, кладизм — доминирующая система взглядов на Западе, и у нас дело идет к тому же. Так проще. Четкая методика, меньше места для интуитивных оценочных функций, вроде оценки уровня сходств и различий, нет отдельных задач построения системы и филогенеза. Особенно велики подобные преимущества у компьютерного (экономного) кладизма. Здесь сохраняется единственный этап, на котором еще нужно думать: это выделение группы для анализа, поиск и отбор признаков для расчета кладограммы. Впрочем, кладисты не утруждают себя анализом методологии этой важнейшей части работы. А дальше все просто: составляем матрицу, вставляем ее в сложную, но детально описанную и потому вполне доступную для пользователя компьютерную программу, запускаем мощный компьютер и ждем окончания расчетов. Если кладограмма не понравилась, можно изменить параметры расчета, и пусть компьютер посчитает еще. В конечном счете он выдаст приемлемый результат, хотя бы минимально согласующийся со здравым смыслом (т.е. с априорными ожиданиями).
Фактически кладизм является ярким примером дедуктивного, а не гипотетико-дедуктивного построения. Действительно, кладизм использует в качестве постулата весьма правдоподобное утверждение — что общее филогенетическое прошлое обуславливает сходство родственных групп. Отсюда делается вывод, не подвергаемый дальнейшей проверке, что система организмов должна быть строго изоморфна их генеалогии, эксплицированной в кладограмме. При этом не принимаются всерьез никакие доказательства, что по крайней мере некоторые из важнейших постулатов кладизма лишены биологического обоснования, а результаты их последовательного применения нередко противоречат здравому смыслу и результатам, полученным другими методами, включая палеонтологический и молекулярный (см. выше). Нет никаких биологических причин для того, чтобы дивергенция проходила только в форме строгой дихотомии с обязательным исчезновением предкового вида в момент дивергенции — наоборот, существуют очень надежные наблюдения над ископаемыми (например, Невесская и др., 1986, Nevesskaya еt аl., 2001), говорящие о том, что предковый вид в течение миллионов лет может давать начало все новым и новым дочерним видам. Равным образом, генеалогическая близость может эффективно отражать отношения по другим свойствам организмов только при существенной равномерности эволюционного процесса, но как уже было показано, палеонтология в избытке предъявляет убедительнейшие свидетельства крайней неравномерности эволюционного процесса.
Получается, что все-таки лучше заниматься традиционной систематикой, но при этом четко понимать ее основания, критерии и цели, сформулированные филетикой, и использовать методы кладизма и фенетики только там, где они могут реально помочь.
|
|
А.П.Расницын. Теоретические основы эволюционной биологии // В.В.Жерихин, А.Г.Пономаренко, А.П.Расницын. Введение в палеоэнтомологию. М.: КМК. 2008. 371 с
1.2.1.3. Компьютерный кладизм
1.2.2. ТАКСОНОМИЯ
1.2.2.1. Кладизм |
1.2.2.2. Фенетика 1.2.2.3. Филетика |