|
|
|||
|
БІОСИСТЕМИ. БІОСФЕРА
ЕКОСИСТЕМИ. ПОПУЛЯЦІЇ
ОРГАНІЗМИ У ДОВКІЛЛІ
ЛЮДСТВО ТА ЙОГО ДОЛЯ
|
|||
|
III-8. Сукцесії. Основні поняття |
|||

III-8. Сукцесії. Основні поняття
Чи доводилося вам опинитися на занехаяному полі або у покинутому саду? Ви, напевно, здогадуєтеся, що за їхнім зовнішнім виглядом можна встановити, коли за ними припинили догляд. Завдяки чому це стає можливим? Залишена сама собі екосистема (в даних прикладах — штучна сільськогосподарська екосистема — агросистема) починає змінюватися. Знаючи швидкість таких змін, можна оцінити, протягом якого часу воно відбувалося.
А тепер уявіть собі річку, що тече по широкій долині. Річище такої річки утворює петлі — меандр. Там, де річище робить поворот, поточна вода підмиває берег, в який вона впирається, і формує береговий обрив. А на протилежному краї такого берега розростається піщаний пляж. Біля самої води пісок голий, трохи далі — поріс травою, ще далі — трава утворює щільну дерновину, і, нарешті, на якійсь відстані від берега ростуть чагарники. Легко зрозуміти, що цей перехід від одного угруповання до іншого (екокліна) відбиває динаміку змін кожної окремої ділянки берега в часі. Там, де зараз росте чагарник, колись тільки намивався пляж, який утворював колись нову ділянку берега... Петлі річки ростуть і з часом з'єднуються один з одним. Річище прокладає собі коротшу дорогу, а його колишні ділянки утворюють стариці. Згодом стариці перетворюються в заплавні озера, наповнюються опадами й стають частинами заплавної луки. Наведені нами приклади показують здатність екосистем до змін у часі — сукцесії (від лат. successio — спадкоємність, успадкування).
Сукцесія — це послідовна зміна угруповань в одному оселищі. Це спрямований, контрольований угрупованням процес, що веде до певного стану. Сукцесія у біогеоценозах є більш тривалим процесом, ніж сезонні зміни, але й не настільки тривалим, як еволюція екосистем. Екокліна, таким чином, є сукцесією, що спостерігається у просторі.
Зазвичай в ході сукцесії можна виділити минущі стадії, які називаються серійними угрупованнями (або просто серіями), і остаточний стійкий стан — клімаксне угруповання (клімакс). Вчення про сукцесії розроблене у 20-і роки XX століття Фредеріком Клементсом, американським екологом, який розглядав сукцесію як якийсь аналог онтогенезу екосистеми.
Причиною сукцесій є не тільки зміна середовища існування якимись зовнішніми чинниками (наприклад, проточною водою, як в описаному випадку з меандром річки), але і вся сукупність взаємодій між компонентами угруповань, а іноді — і впливу з боку людини.
«Характерна динаміка угруповань, описана для кінця XIX — початку XX століть у вересових торфовищах Шотландії. Господар цих земель використовував їх для полювання на численних тут шотландських куріпок — граусів. У 1892–1893 рр. на верещатнику оселилося кілька пар чайок. Взяті господарем під охорону, вони на початку нашого століття вже утворили велику колонію, що налічувала 1,5–2 тис. пар. Птахи рясно удобрювали ґрунт, гігроскопічний матеріал їх гнізд накопичував вологу. В результаті почалося поступове заболочування, яке призвело до поступового зникнення вересу і заміни його жорсткими злаками. Угіддя стали використовуватися як сінокісні. Однак надалі злаки були витіснені ситником, пізніше — кінським щавлем та іншими бур'янами. Паралельно йшло зменшення чисельності граусу; на заболочених ділянках з'явилися качки — вид, менш цінний як мисливський трофей. Все це призвело до скасування охорони чайок; мало того, їхні гнізда стали навмисно розоряти. У 1917 році залишилося близько 30 пар, поступово відновлювався верещатник, зникли качки, знову з'явилися граус. Однак і до цього часу відновлення вихідного угруповання було неповним: на вересовій пустці збереглися злаки та деякі бур'яни — «слід» сукцесійної серії» (І.О. Шилов, 1998).
Основна причина сукцесій — відсутність рівноваги між продукцією і диханням в екосистемі, тобто нерівновага екологічного балансу (див. пункт III-5). Це призводить до зміни запасу органіки в екосистемі та, зрештою, до зміни угруповання. Розташувавши різноманітні екосистеми на площині «продукція-дихання» (рис. III-8.1), ми можемо переконатися, що стійкими з них виявляються тільки ті, в яких продукція і дихання врівноважують одне одного.
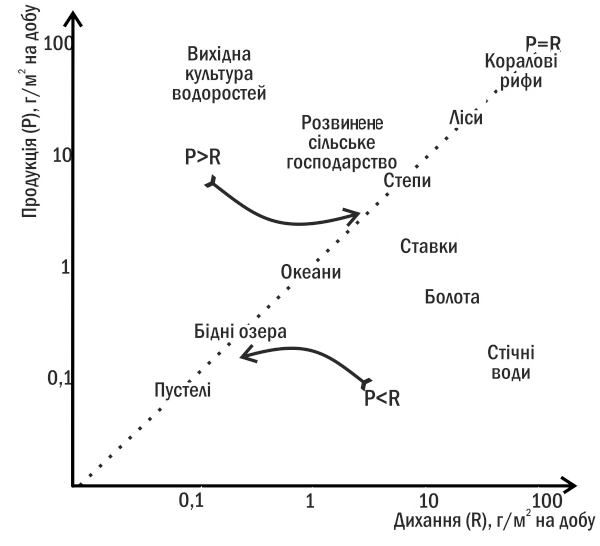
Рис. III-8.1. Ординація (розташування) різних типів екосистем на площині «продукція - дихання»
Якщо в якомусь угрупованні продукція перевищує дихання (наприклад, у вихідній культурі водоростей — розчині мінеральних солей, в який заселені фітопланктонні організми), у ньому починається автотрофна сукцесія. Надлишок органіки накопичується в такій екосистемі, змінюючи її властивості та створюючи середовище для розмноження гетеротрофів. Продукція в угрупованні знижується, дихання збільшується, і, врешті-решт, ці дві величини зрівнюються.
Гетеротрофна сукцесія (рис. III-8.2) починається зі стану, в якому дихання угруповання перевищує продукцію. Поступово гетеротрофи руйнують надлишок органіки, і в системі досягається рівновага між диханням і продукцією.
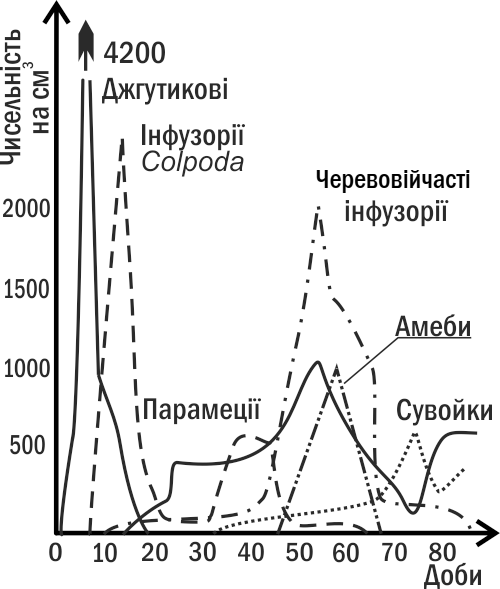
Рис. III-8.2. Приклад гетеротрофної сукцесії: динаміка чисельності різних найпростіших у сінному настої
Характерним випадком автотрофної сукцесії можна вважати заселення організмами території, на якій немає запасів органічної речовини. Розглянемо наступний приклад. На голій скельній породі поселяються лишайники, поступово роз'їдають камінь за допомогою виділених ними кислот. У тріщинах гірських порід з'являється ґрунт. На скелі починають рости мохи, а потім і трави. Формуючи дерновину з переплетених відмерлих стебел і кореневищ, трави утримують детрит і навіть збирають пил. Через дуже тривалий час на місці колишніх скель з'являються чагарники, а потім і ліс. Це була первинна сукцесія — сукцесія, яка йшла там, де не було запасів органічної речовини від колишніх угруповань. Інший приклад первинної сукцесії показано на рис. III-8.3.
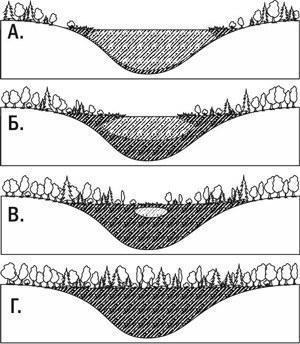
Рис. III-8.3. Приклад первинної сукцесії: замулювання озера
За якийсь час ліс, що виріс у ході первинної сукцесії, знищується пожежею. Утворюється згарище. На згарищі зберігаються залишки лісового ґрунту (рясно присипаного попелом) і навіть насіння багатьох видів рослин. Вже на наступний рік гар заросте травами (наприклад, у таких умовах дуже характерний іван-чай). За кілька років на місці згарища з'являться чагарники та молоді дерева, а вже за кілька десятиліть або через століття тут розвинеться ліс, що нагадуватиме той, що ріс тут до пожежі. Вторинна сукцесія проходить в оселищах, де збереглися залишки органічної речовини від попередніх етапів розвитку даної екосистеми (рис. III-8.4.)
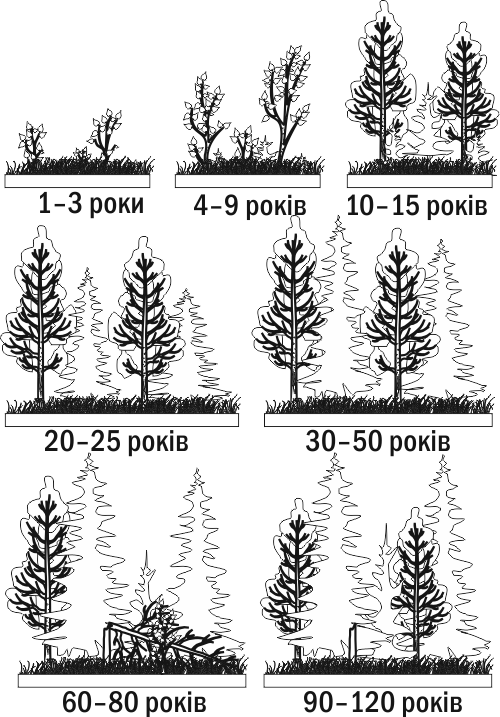
Рис. III-8.4. Приклад вторинної сукцесії: заростання згарища змішаним лісом
Гетеротрофна сукцесія вимагає для свого протікання запасу органіки. Більшість прикладів гетеротрофної сукцесії, які ми можемо спостерігати, розгортаються не в масштабах біогеоценозів, а в межах відносно невеликих екосистем. Наприклад, до їх числа належать тіла померлих тварин або стовбури повалених рослин. Сукцесії, що протікають в таких тимчасових екосистемах, називаються деструктивними — вони не ведуть до клімаксу, а просто знищують те тимчасове оселище в якому відбуваються. Іноді можна спостерігати екокліни й в гетеротрофних сукцесіях. Розкопайте опалу хвою і верхній шар ґрунту у хвойному лісі. Зверху хвоїнки майже не пошкоджені, а чим глибше — тим більше вони перетворені бактеріями, грибами та тваринами-детритофагами. Кожна опала голка проходить цей шлях руйнування, відповідний послідовності шарів хвойного опаду.
Клементс був прихильником концепції моноклімаксу. Нині перемогла концепція поліклімаксу, згідно з якою для кожного регіону існує певний кліматичний або зональний клімакс, і, крім того, в ньому можуть бути стійкі певні едафічні (тобто зумовлені особливостями ґрунтів) або локальні клімакси. Описано також циклічний клімакс, при якому екосистема проходить через сукупність певних станів, які змінюють один одного по колу. Іноді ці зміни визначаються катастрофічними подіями, що закономірно відбуваються на певному етапі розвитку екосистем. Так, ялинник після досягнення певної зрілості може закономірно знищуватися шкідниками; чапараль може регулярно вигоряти при пожежах.
Можна розглядати як загальні, так і спеціальні сукцесії, що стосуються змін видів у певних екологічних нішах. Так, у плодовому саду на молодих деревах харчуються багатоїдні комахи-фітофаги: хрущі, чорнотілки, п'ядуни, шовкопряди, листовійки. Коли дерева починають плодити, з'являються плодожерки, квіткоїди, довгоносики, що харчуються квітками та плодами. У міру старіння саду його населяють стовбурові шкідники (короїди, деревоточці, скляниці).