Авторское предуведомление к сетевой публикации
Благодаря любезности создателя «Макроэволюции» Александра Маркова я получил возможность опубликовать статью, написанную в 2002 году; теперь выкладываю ее и на "Батрахосе". Должен признаться, что эта статья встретила критическое отношение, и была отправлена на переделку редакцией "Журнала общей биологии", куда я ее предложил. Я весьма благодарен за критику, высказанную Алексеем Меркурьевичем Гиляровым. Коротко резюмируя ее направленность, могу сказать, что она касается недостаточной «развернутости», обоснованности ряда утверждений. К сожалению, моя попытка переделки этой статьи убедила меня, что для достаточного обоснования изложенной в ней идеи статью нужно превращать в книгу. В то же время я надеюсь, что и в нынешней форме этот текст окажется источником материала для обсуждения с единомышленниками и конструктивно настроенными «противниками» - сторонниками иных взглядов.
Из отзывов на рукопись этой статьи для меня был особо важен отклик Михаила Александровича Шишкина. Этот отклик также оказался острокритическим. С точки зрения Михаила Александровича, объяснительные подходы синтетической и эпигенетической теории принципиально различны, и их смешение, демонстрируемое в предлагаемой статье, не может привести ни к чему хорошему. На мой взгляд, такое совмещение необходимо. Я думаю, что одной из насущных задач построения новой эволюционной теории является построение хорошей классификации признаков по характеру управления их развитием. Когда речь идет о биохимических признаках бактерий (особенно признаков «есть поломка фермента»/«нет поломки фермента») методология синтетической теории эволюции оказывается вполне достаточной. Если нас интересует выработка новых адаптаций у представителей быстро эволюционирующих высокоорганизованных групп организмов, мы не сможем объяснить их на языке отбора аллелей, и должны будем обратиться к подходу эпигенетической теории.
Еще одно необходимое предуведомление касается использования термина «эпигенетический». Это понятие может использоваться в нескольких разных смыслах, даже если не касаться знаменитой дискуссии эпигенетиков и преформистов в истории биологии развития. Эти смыслы можно назвать молекулярным и онтогенетическим. Оба они в общих чертах отвечают определению К.Х. Уоддингтона, автора этого понятия в его современной трактовке. Согласно Уоддингтону, эпигенетикой называют раздел биологии о причинных взаимодействиях между генами и их продуктами, образующими фенотип. Разрыв между молекулярным и онтогенетическим подходами к эпигенетике отражает разрыв между уровнями исследования, доступными для современной науки. Мы можем изучать развитие или на уровне отдельных молекул, или на уровне онтогенеза как целого. Описание взаимовлияния всех факторов управления развития даже на уровне единственной клетки – неподъемная для современной науки задача. Впрочем, сам Уоддингтон использовал обсуждаемый термин скорее в онтогенетическом смысле, как, например, в своей известной метафоре эпигенетического ландшафта. В онтогенетическом смысле этой понятие используется и в этой статье (как и в работах М.А. Шишкина).
Считаю смыслом этой сетевой публикации обсуждение рассматриваемых в ней идей. Буду благодарен за конструктивную критику.
Д. А. Шабанов
Каким будет третий синтез в теории эволюции?
В первой половине XX века кризис классического дарвинизма (первого эволюционного синтеза) вызвал появление второго синтеза — синтетической теории эволюции (СТЭ). Современный кризис СТЭ должен привести к появлению третьего синтеза, который преодолеет ее редукционистскую ограниченность. Возможные элементы третьего синтеза разрабатываются сегодня относительно независимо друг от друга. Это экологическая теория В. А. Красилова, неосальтационизм, концепции видового мономорфизма, прерывистого равновесия, «эволюции эволюции» и т.д. Одним из следствий эпигенетической теории М. А. Шишкина, которая может стать ядром третьего синтеза, является новый подход к решению проблемы возникновения биологической целесообразности. Можно предположить, что в рамках третьего синтеза будет найдено объяснение механизма приобретения в ходе эволюции целостных комплексов признаков, многоуровневости эволюции и изменения ее темпов.
Накануне третьего эволюционного синтеза
Согласно периодизации развития эволюционного учения, предложенной Н. Н. Воронцовым (1999), дарвинизм стал первым широким синтезом эволюционных идей. В конце XIX— начале XX века наступил кризис первого синтеза. Некоторые из вызвавших этот кризис проблем были решены вторым синтезом — синтетической теорией эволюции (СТЭ). Нынешнее время — период кризиса СТЭ, предшествующего третьему эволюционному синтезу. Некоторые черты грядущего синтеза начали складываться уже несколько десятилетий назад (например, Шварц, 1967; Красилов, 1984; Шишкин, 1987; Назаров, 1991), однако этот процесс еще далеко не закончен.
Современное состояние эволюционной теории позволяет предугадать ряд особенностей третьего синтеза. Кризис эволюционизма стимулировал появление ряда новых теорий. Главная мысль данной работы — утверждение, что упомянутые в статье концепции, рассматривающие процесс эволюции с разных точек зрения, хорошо согласуются между собой. Их объединение, по мнению автора, должно привести к появлению третьего эволюционного синтеза. После характеристики некоторых черт кризиса СТЭ в этой статье рассматривается эпигенетическая теория эволюции, которая, по мнению автора, должна стать ядром нового синтеза.
Некоторые авторитетные эволюционисты высказывали мнение, что накопление новых данных означает не замену СТЭ новым синтезом, а лишь видоизменение и расширение самой СТЭ. В качестве аналогии можно упомянуть взгляды И.И. Шмальгаузена, считавшего излишним разграничение дарвинизма и неодарвинизма (СТЭ). По мнению автора, в этом вопросе не следует смешивать науку в целом (в данном случае — эволюционную биологию) и ту или иную теорию (дарвинизм или СТЭ). Наука развивается, переходя от одних взглядов к другим. Теория — продукт определенного времени, характеризующийся относительно целостными взглядами. Так, для СТЭ характерно представление об эволюции, как об изменении частот аллелей в популяциях. Отказ от этого подхода не расширяет рамки СТЭ, а требует перехода к другой теории.
Кризис СТЭ
И первоначальный успех, и современный кризис СТЭ связаны с ее редукционизмом. Проявления его различны: это и сведение видообразования и макроэволюции к микроэволюции, и отрыв эволюции популяций от изменений биогеоценозов и биосферы, и принятие допущения о независимой эволюции отдельных генов. В частности, математическая теория отбора основана на предположении, что все аллели имеют постоянную приспособительную ценность. При этом не учитываются такие важные факторы, как влияние генотипа, неопределенность результатов онтогенеза (обусловленная взаимодействием внешних и внутренних факторов), а также экологическая неоднородность среды.
Называя проблемы, которые были решены благодаря синтезу теории эволюции и генетики, сторонники СТЭ часто ссылаются на критику дарвинизма Ф. Дженкином (Jenkin, 1867). Согласно этому автору, адаптивный признакне может сохранится в ряду поколений из-за «размывания» при скрещивании его носителей с лишенными данного признака особями. «После переоткрытия законов Менделя и доказательства, что факторы, определяющие развитие наследуемых признаков, передаются потомкам не дробясь, «кошмар Дженкина» был развеян» (Сойфер, 1975, с. 309). Однако очевидно, что Дженкин имел в виду отнюдь не моногенные признаки: «Предположим, белый человек потерпел кораблекрушение на острове, населенном неграми... Наш потерпевший кораблекрушение герой, вероятно, станет королем; он убьет великое множество черных в борьбе за существование; он будет иметь огромное количество жен и детей, в то время как многие его подданные будут жить и умрут холостяками. ... В первом поколении будет несколько дюжин смышленых молодых мулатов, в среднем превосходящих по интеллекту негров. Мы можем ожидать, что трон в течение нескольких поколений будет занимать более или менее желтый король; но сможет ли кто-нибудь поверить, что весь остров постепенно приобретет белую или даже желтую популяцию, или что островитяне приобретут энергию, храбрость, изобретательность, настойчивость, самоконтроль, выносливость, в силу которых наш герой убил так много их предков и породил так много детей, то есть те качества, которые фактически отбирает борьба за существование, если она может что-то отбирать?» (Jenkin, 1867, p. 289–290).
Для оценки рассуждения Дженкина не важно, что расового превосходства не существует, а названные признаки наследуются в основном культурно. Если адаптивность особи зависит от комплекса независимо наследуемых признаков, «рассыпание» такого комплекса должно препятствовать отбору. Предположение, что каждый из таких признаков будет отбираться независимо от других, опровергнуто в рамках самой СТЭ. Разрабатывая математическую теорию отбора, Дж. Холдейн (Haldane, 1957) показал, что отбор по нескольким независимым парам аллелей неэффективен из-за нарастания генетического груза и снижения репродуктивного потенциала популяции. Допущение о длительном отборе по множеству отдельных генов, каждый из которых вносит свой вклад на уровне фенотипа (особенно характерное для развившейся на основе СТЭ социобиологии) не только противоречит расчетам Холдейна, но и опровергается данными об относительной немногочисленности генов человека и других видов, полученным в ходе работ по расшифровке геномов.
Созданию СТЭ способствовало открытие С. С. Четвериковым генетического разнообразия носителей нормальных фенотипов из естественных популяций. Однако с точки зрения СТЭ этот феномен так и не нашел должного объяснения (Левонтин, 1978). Не менее сложно установить причины устойчивых во времени различий между соседними популяциями и группами размножения, обнаруженных у многих видов. Для объяснения этих явлений сторонники СТЭ прибегают к ссылкам на дрейф генов. Фактически это означает отказ от селекционизма, лежащего в основе самой СТЭ.
Изучение филогенеза многих таксонов показало ошибочность характерного для СТЭ представления о преимущественно дивергентном характере эволюции, тесно связанного с признанием ненаправленности мутаций и дрейфа генов. Параллелизмы и конвергенции являются нормой в становлении большинства групп (Татаринов, 1987).
Из сказанного вытекает, что теория, которая придет на смену СТЭ, должна будет объяснить механизм приобретения в ходе эволюции согласованных комплексов адаптивных признаков.
Некоторые другие черты кризиса второго синтеза рассмотрены далее в сравнении с альтернативными взглядами.
Эпигенетическая теория М. А. Шишкина
Центральную роль в третьем эволюционном синтезе (Гродницкий, 2001) может сыграть концепция М. А. Шишкина (1987, 1988), названная им «эпигенетической теорией эволюции Шмальгаузена-Уоддингтона». Хотя автор и исходит из идей И. И. Шмальгаузена и К. Х. Уоддингтона, он приходит к существенно иным выводам. Поэтому, вероятно, правильнее говорить об «эпигенетической теории М. А. Шишкина».
Ключевое понятие данной теории — эпигенетическая система (далее — ЭС), совокупность взаимодействий генетических и иных факторов, влияющих на онтогенез. В соответствии с этой теорией отдельные наследственные задатки не являются непосредственной причиной тех или иных особенностей фенотипа, а только влияют на систему управления онтогенезом. В результате естественного отбора отбираются и воспроизводятся в потомстве не отдельные аллели или признаки, а целостные фенотипы. Независимо от того, какие факторы (генетические и негенетические наследственные задатки, внешние воздействия) вызывают развитие адаптивного признака, его обладатели внесут относительно больший вклад в формирование следующего поколения.
Потомки наследуют от родителей не только гены, но и общую организацию ЭС. Если характер отбора остается прежним, преимущество получают те из них, которые воспроизводят в своем развитии отобранные в предыдущем поколении адаптивные фенотипы. Отбор таких фенотипов должен привести к перестройке ЭС, повышающей устойчивость приводящих к ним траекторий развития (креодов).
Пока популяционная норма остается адаптивной, отбор перестраивает ЭС таким образом, чтобы при максимально широком разнообразии генотипов обеспечивался нормальный ход онтогенеза. Следствием этого как раз и является обнаруженное С. С. Четвериковым генетическое разнообразие носителей нормального фенотипа из естественных популяций. Отбор против сложившейся фенотипической нормы дестабилизирует ведущие к ней эпигенетические креоды. В результате проявляется ряд относительно дискретных аберраций, которые соответствуют возможным для данной ЭС альтернативным креодам. Отбор в пользу одного из них увеличивает устойчивость его реализации в онтогенезе.
Таким образом, в соответствии с рассматриваемыми взглядами, новые признаки появляются вследствие преобразования видоспецифичной ЭС путем отбора. При этом стабилизируются варианты развития, первоначально возникающие как аберрации.
Сказанное подтверждается многочисленными фактами. Как известно, однозначного соответствия ген–признак не существует. Действие каждого гена зависит от других наследственных задатков и характеризуется той или иной экспрессивностью и пенетрантностью. Особенно неустойчиво проявление мутантных аллелей, реализация которых в онтогенезе не стабилизирована отбором: даже доминантные мутации не всегда не проявляются в гомозиготе. Р. Гольдшмидт (Goldschmidt, 1940) показал, что один и тот же признак может развиваться или не развиваться в силу весьма различных причин (различных влияний на ЭС): и мутаций различных генов, и воздействий среды.
Развитие эпигенетической теории началось с опытов Уоддингтона, в которых была продемонстрирована стабилизация лабильных признаков. В этих опытах отбор на способность формировать морфозы (аномалии развития) dumpy или bithorax в ответ на температурные или токсические воздействия приводил к тому, что эти морфозы начинали развиваться и при нормальных условиях (Waddington, 1957). С точки зрения концепции М. А. Шишкина, такой результат обусловлен перестройкой ЭС, стабилизирующей отбираемые пути развития (а не возникновением генов dumpy или bithorax).
В экспериментах Г. Х. Шапошникова (1978), в результате отбора морфозов тлей, вызванных сменой кормового растения, возникали новые, морфологически и экологически своеобразные формы экспериментальных животных, репродуктивно изолированные от исходных.
Эпигенетическая система как уровень интеграции
Возникает закономерный вопрос: следует ли рассматривать ЭС, как отдельный уровень регуляции развития, если взаимодействие генов рассматривается и в классической генетике? Не является ли понятие «эпигенетическая система» синонимом понятия «генотип»?
Очевидно, что выделение отдельного структурного или функционального уровня интеграции оправдано в том случае, когда на нем возникают некие новые (эмергентные) свойства, отсутствующие на нижележащих уровнях. Недооценка одного из существенных уровней интеграции (уровня n) приведет к типичной редукционистской ошибке: особенности уровня n+1 будут интерпретироваться как результат функционирования уровня n–1. При этом эмергентные свойства уровня n станут недоступными для изучения.
Онтогенез, один из сложнейших известных процессов, регулируется разнообразными наследственными задатками, средовыми влияниями и их взаимодействием. Эквифинальность индивидуального развития и зависящая от внешних условий возможность его адаптивного изменения задаются на уровне не отдельных генов, а ЭС в целом. Рассмотрение этого уровня является необходимым условием изучения важнейших свойств живых систем.
Где локализованы механизмы канализации развития? Этот вопрос не более корректен, чем, например, вопрос о том, в каком месте мозга находится данная мысль, или где в биосфере расположен регулятор содержания кислорода в атмосфере. Когда речь идет о сложных системах (ЭС, мозге, биосфере и др.), указать локализацию их эмергентных свойств попросту невозможно, так как они задаются совокупностью связей между всеми элементами. Впрочем, эпигенетические механизмы проявляются и на уровне отдельных наследственных задатков, мышление связано с функционированием клеток мозга, а биосферный гомеостаз осуществляется благодаря жизнедеятельности отдельных организмов.
Закономерно, что ядром первого эволюционного синтеза стало исследование изменчивости фенотипов и их взаимодействия со средой — наиболее «явного» уровня регуляции исторического развития (рис. 1). Изучение относительно простого механизма наследственной изменчивости и влияния отбора на частоты наследственных задатков привело ко второму синтезу. Представляется, что в ходе третьего синтеза будет понято значение самого сложного уровня регуляции развития — ЭС.
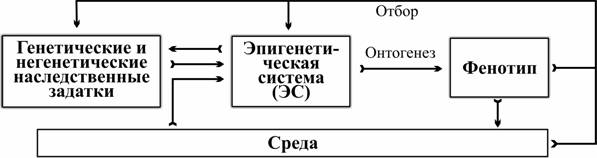
Рис. 1. Роль ЭС в управлении индивидуальным и историческим развитием
Преодоление «геноцентризма»
Трактуя описанные выше опыты по закреплению морфозов в результате отбора, К. Х. Уоддингтон назвал открытое им явление генетической (а не эпигенетической) ассимиляцией. Для объяснения результатов аналогичных опытов высказывалось предположение, что первоначальные феноадаптации замещаются геноадаптациями (Шмальгаузен, 1938; Гаузе, 1984). Оно основано на убеждении, что каждый признак обусловлен специфичными генами. Подобный «геноцентризм» широко распространен до сих пор. Например, утверждение, что «материалом для эволюции ... служат мутации и их комбинации», Н. Н. Воронцов (1999, с. 470) относит к числу положений, «о которых не спорят эволюционисты».
Показано, что близкие виды могут существенно отличаться организацией своего генетического материала, например, соотношением разных типов последовательностей (Корочкин, 1985). В то же время близость этих видов связана со сходством их ЭС. По-видимому, именно сходство ЭС разных видов является причиной существования гомологических рядов изменчивости, описанных Н. И. Вавиловым. Так как гомологические аберрации развития могут иметь различную генетическую основу, гомологическими являются не гены, а эпигенетически детерминированные креоды.
Как указывает А. С. Раутиан (1993), в силу принципа относительности содержания информации А.А. Ляпунова, информация в генотипе доступна только для определенного компетентного пользователя. Таким пользователем является не фенотип в целом, а именно ЭС, первоначально наследуемая от предыдущего поколения (например, при образовании яйцеклетки), а затем изменяющаяся в онтогенезе. Изменение фенотипов в ходе эволюции связано с не столько с изменением генов (и других наследственных задатков), сколько с изменением ЭС. Одна и та же последовательность нуклеотидов, прочтенная ЭС различных организмов, может иметь для них совершенно разный смысл! Рассмотренная в популярном кинофильме (и романе) возможность получить динозавра, пересадив его гены в яйцеклетку жабы, является необоснованной фантазией: таким путем можно получить разве что эмбрион жабы с нарушенным взаимодействием генотипа и ЭС. Как известно, в ходе антропогенеза последовательности ДНК человека претерпели лишь незначительные изменения. Таким образом, человек отличается от шимпанзе не столько тем, что имеет иные гены, сколько наличием реорганизованной ЭС.
«Геноцентризму» противоречит также все возрастающее число примеров негенетического наследования. По-видимому, динамическое наследование (передача специфической конформации белковых молекул), проявляющееся при прионных заболеваниях, является не аномалией, а результатом действия общего механизма (Инге-Вечтомов, 2000). Так, белки-шапероны принимают участие в синтезе многих полипептидных цепей, определяя их конформацию. Способностью к наследованию обладает даже цитоскелет, передающий особенности пространственной организации клеточных структур (Албертс и др., 1987, т. 3, с. 132). Культурное наследование у человека (и, в меньшей степени, у некоторых других видов) обеспечивает передачу приобретенных признаков и переход к более эффективным способам выработки адаптаций.
Как показано на рис. 1, ЭС не только зависит от наследственных задатков, но и влияет на них. Примером таких влияний на генетическую наследственность могут служить эпигены (Голубовский, Чураев, 1987), а на негенетическую — динамически наследуемые изменения конформации белков. Экспериментально установлена способность бактерий перестраивать свой генетический аппарат в течение жизни одной клетки, обеспечивая изменения генов, необходимые для питания на определенной среде (Голубовский, 2001). При выращивании льна на обогащенной биогенами почве происходила выработка наследственных изменений, приводящих к увеличению размеров растений (Marx, 1984). Приведенные факты могут показаться отвечающими взглядам Т. Д. Лысенко. Однако принципиальной особенностью этих результатов является то, что они получены в корректных экспериментах и лишены идеологической подоплеки.
Как известно, характерной особенностью смены научной парадигмы является переосмысление известных фактов. Эпигенетическая теория дает основания для ревизии таких понятий, как ген, мутация, модификация, движущий отбор и др (Шишкин, 1987). Неправомерным оказывается отождествление менделевских генов с локализованными в хромосомах цистронами. На самом деле, менделевский ген является переключателем между двумя эпигенетически стабилизированными траекториями индивидуального развития, а не причиной их возникновения. Вопреки общепринятым взглядам, можно выделить всего два основных типа отбора: стабилизирующий и дестабилизирующий. Признаки следует делить не на наследуемые (генотипические) и ненаследуемые (фенотипические), а на устойчивые и лабильные в своем проявлении.
Проблема целесообразности решена?
Проблема биологической целесообразности стала ключевой проблемой для науки о природе еще во времена Аристотеля. Представление, что целесообразные организмы возникли случайно, появилось в античности и в различной форме было выражено Эмпедоклом, Эпикуром и Лукрецием. Дарвинизм добавил к этому представлению гипотезу о длительных последовательностях малых ненаправленных изменений, случайно повышающих приспособленность организмов. Современная нелинейная термодинамика изучает самоорганизацию как следствие случайных возмущений. Тем не менее, представление о возникновении целесообразности на основе случайных уклонений нельзя считать окончательно доказанным. Ему противоречит тот факт, что мутации обычно неблагоприятны и не интегрированы в генотип. Для ученых, убежденных в совершенстве природы, идея случайного возникновения целесообразности неприемлема. Вероятно, именно поэтому Л. С. Берг постулировал имманентную, недоступную научному анализу целесообразность живого (Берг, 1977).
Эпигенетическая теория открывает возможность нового решения проблемы целесообразности, выходящего за рамки предложенной Бергом классификации (там же, с. 99–101). Целесообразность мутаций, постулируемая в СТЭ, — явление на грани чуда. Напротив, вызванные дестабилизацией нормального развития аберрации потенциально целесообразны, так как в их формировании участвует ЭС, сформированная предшествовавшей эволюцией данного вида и жизни в целом. Мутация гена не связана с соответствующими изменениями других элементов генотипа; эпигенетическая аберрация может затрагивать многие признаки при сохранении корреляций между ними. Мутации возникают случайно, а аберрации — когда прежняя популяционная норма перестает быть адаптивной. Наконец, как отмечает Д. Л. Гродницкий (2001), закономерность появления аберрантных фенотипов обусловливает достаточную вероятность встречи их носителей.
Если какая-то из возникающих аберраций оказывается адаптивной, в популяции происходит процесс, моделируемый описанными опытами Уоддингтона: отбор на устойчивость ее воспроизведения в ряду поколений. Конечный этап при этом — стабилизация новой нормы.
Итак, новый признак проходит такие этапы своего становления: более или менее адаптивная «модификация», развивающаяся при соответствующем воздействии среды; один из альтернативных вариантов развития, все более устойчиво реализующийся в онтогенезе; норма, для развития которой не требуется специфических внешних воздействий.
Чтобы объяснить появление (зачастую параллельное у разных групп организмов) комплексов целесообразно взаимосвязанных признаков следует рассматривать влияние регуляторного механизма, лежащего выше уровня отдельных наследственных задатков — ЭС. Именно таков путь преодоления «кошмара Дженкина».
П. Тейяр де Шарден (1987) утверждал, что жизнь движется вперед «на ощупь». Эти слова обычно понимали как констатацию неизбежности накопления случайных ошибок. Вероятно, аналогия Тейяра значительно глубже. Хотя в эволюционном процессе нет заранее определенного конечного состояния, в нем нет и случайного метания из стороны в сторону. Направление каждого шага эволюции обусловлено реакцией целостного организма на изменение среды, в которой он развивается.
Экологическая теория эволюции
Характерный для СТЭ редукционизм проявляется не только в представлении системы управления развитием в виде мозаики генов, но и в трактовке популяции как основного театра эволюционных событий.
Каждый биоценоз обладает определенной способностью к поддержанию гомеостаза. Изменения внешних (например, климатических) факторов могут компенсироваться внутренними регуляторными механизмами данного биоценоза. Именно они поддерживают характерную структуру экологических ниш в определенном диапазоне значений внешних условий. При этом на популяции действует стабилизирующий нормальное развитие отбор, вследствие чего они не изменяются или эволюционируют согласованно и медленно. Эволюцию в пределах стабильного биоценоза В. А. Красилов (1969) назвал когерентной эволюцией. В случае распада биоценоза наблюдается быстрая некогерентная эволюция отдельных его элементов. Популяции наиболее пластичных и наименее интегрированных в разрушенное сообщество видов способны существовать и в изменившихся условиях. Прекращение стабилизирующего отбора и разнообразие условий развития приводят к дестабилизации онтогенеза и проявлению резерва изменчивости, характерного для ЭС данных видов. В неустойчивых сообществах преимущество получают r-стратеги. Результатом r-отбора становится ускорение индивидуального развития, которое может приводить к частичной потере специализации, достигавшейся на последних этапах онтогенеза. Мозаичность участков территории, находящихся на различных стадиях сукцессии и формирования новых биоценозов, способствует квантовой эволюции.
Так, в антропически измененных биоценозах у ряда видов зарегистрировано быстрое возникновение адаптивных признаков (например, новых форм гнездового поведения птиц или упрощения жизненного цикла проходных рыб).
В эпигенетической теории не рассматриваются причины того, что отбор начинает поддерживать одну из аберраций, а не прежнюю норму. Экологическая теория объясняет скачкообразное изменение характера отбора. Обе концепции акцентируют внимание на целостных свойствах изучаемых систем и удачно дополняют друг друга.
Концепция видового мономорфизма
Интересно сопоставить выводы эпигенетической теории эволюции с данными о существовании мономорфного комплекса генетических и физиологических признаков у видов. «Одно из важнейших свойств эукариотического генома — двойственность его структурно-функциональной организации, находящая прямое отражение в сосуществовании двух реальных явлений: полиморфизма и мономорфизма» (Алтухов, 1989, с. 219). Внутривидовой мономорфизм характерен для генетических систем с множеством структурных компонентов и высокой избыточностью (там же, с. 203), то есть для регуляторной части генома.
Аналогично, Х. Карсон различает две системы генетической изменчивости: «открытую», представленную локусами, обеспечивающими внутривидовой полиморфизм, и «закрытую», связанную с коадаптированными блоками генов (Carson, 1982). Видообразование, с этой точки зрения, является следствием реорганизации «закрытой» части генома.
Д. К. Беляев (1974) установил, что дестабилизирующий отбор по одним признакам приводит к проявлению широкой изменчивости других, ранее стабильных, признаков. Возможно, это явление связано с реорганизацией «закрытой» части генома и ЭС.
Изучение изменчивости генома приводит ко вполне согласующемуся с эпигенетической теорией выводу: «на надвидовом уровне эволюция — это не только или, точнее, не столько процесс появления новых генов с новыми функциями, сколько быстрая реорганизация генетического материала с последующим развитием новых систем взаимодействия (и регуляции) генов на посттранскрипционном и посттрансляционном уровнях» (Алтухов, 1989, с. 216).
С. С. Шварц показал, что «приспособления специализированных видов и приспособления отдельных популяций широко распространенных видов идут принципиально различными путями» (1980, с. 45). Адаптации внутривидовых форм к изменяющимся условиям среды выражаются в морфофункциональных сдвигах (изменении размеров сердца или почек, кислородопоглотительной способности крови и т.п.). Видовые адаптации происходят на тканевом и биохимическом уровнях, что делает ненужными морфофункциональные сдвиги. Целостность вида, помимо генетических механизмов, поддерживается видоспецифичной реакцией на изменения условий среды (там же, с. 132), которую можно рассматривать и как отражение общности ЭС.
Некоторые признаки, демонстрируя различия между разными таксонами, оказываются мономорфными внутри большинства из них (и этим напоминают видовые признаки, связанные с «закрытой» частью генома и ЭС). Так, все млекопитающие имеют одинаковый размер эритроцитов, иной, чем у птиц, рептилий, рыб и бесхвостых амфибий. Единственная группа позвоночных, внутри которой зарегистрировано существенное разнообразие размеров эритроцитов — хвостатые амфибии (Шмидт-Ниельсен, 1987, с. 128).
Вероятно, в рамках третьего синтеза придется отказаться от характерного для СТЭ градуализма (вытекающего из представления о том, что приспособленность возникает в результате случайно полезных мутаций). Можно предположить, что в нем будут разграничены периоды стабилизации и дестабилизации системы управления развитием. Представляется обоснованным предположение, что мономорфная часть видового генома является основой видоспецифичной ЭС. Занятие новой адаптивной зоны, возникновение таксонов того или иного уровня, скорее всего, будет происходить в результате перестройки основных характеристик ЭС.
В этой связи представляет интерес эпигенетическая концепция вида предложенная К. Е. Михайловым в работе, опубликованной в Internet (Михайлов, url). Согласно этой концепции, основная характеристика вида — стабилизированный онтогенез. При таком подходе популяции и особи, находящиеся на переходном этапе видообразования в период дестабилизации онтогенеза, следует рассматривать как девиантные формы, близкие к одному из видов, но не относящиеся к нему. Развивая взгляды Михайлова, можно включить наличие стабилизированных эпигенетических креодов и в характеристику надвидовых таксонов. Неожиданным следствием этого подхода может стать обоснование выделения переходных групп, не относящихся к таксонам некоторых более высоких рангов, то есть форм incertae sedis(например, семейств, не относящихся ни к одному из отрядов своего класса).
Неосальтационизм и прерывистое равновесие
Одним из возможных путей реорганизации генома являются хромосомные перестройки (Алтухов, 1989; Воронцов, 1999). Вероятна тесная связь между реорганизацией генома и гибридизацией (Боркин, Даревский, 1980).
В последние десятилетия получены многочисленные подтверждения значения скачкообразных событий для эволюции, заставившие вспомнить о сальтационизме, казалось бы, опровергнутом вторым эволюционным синтезом. Изучение кариотипов близких видов показало важную роль хромосомных перестроек и полиплоидии в видообразовании не только у растений, но и у животных (в том числе в весьма прогрессивных группах, например у жаб и грызунов). Это позволяет предполагать, что во многих случаях возникновение частичной либо полной изоляции предшествует экологической или морфофизиологической дифференциации (Воронцов, 1999). В таком случае конкурентное исключение кариологически изолированных форм должно способствовать их дивергенции.
Можно привести примеры достаточно изученных сальтационных событий. Штамм Clostridium botulinum, вызывающий ботулизм, при заражении определенным фагом превращается в Clostridium novyi, возбудителя газовой гангрены (Eklund et al., 1974). При возникновении нескольких родов млекопитающих происходило закрепление аберраций hairless, мопсовидности, папилломатоза (Воронцов, 1999, с. 507–507). Один из ферментов (Cu-Zn-супероксидисмутаза) биолюминесцирующей бактерии Photobacter leiognathi сходен не с аналогичными ферментами прокариот, а с ферментами сребробрюшковых рыб Leiognathidae, симбионтами которых являются эти бактерии. Это свидетельствует о «горизонтальном переносе» наследственной информации (там же, с. 518). Изменение гормонального баланса добавлением тироксина вызывает у илистого прыгуна (Periophthalmus) перестройку онтогенеза, включающую развитие «конечностей» вместо грудных плавников (Harms, 1934). Согласно данным молекулярной систематики основные группы организмов возникли в результате последовательных эндосимбиозов (Кусакин, Дроздов, 1994) — скачкообразных эволюционных событий. Наконец, некоторые эволюционные события (например, происхождение вторичноротых животных от первичноротых, соединение хоанами обонятельного канала с ротовой полостью у рыб) не могли происходить градуально.
Приведенные и другие факты показывают недопустимость характерного для СТЭ сведения макроэволюции и видообразования к микроэволюции. Неосальтационизм, как и эпигенетическая, и экологическая теории, объясняет неравномерность темпов эволюции, на которой зиждется концепция прерывистого равновесия. Возможность освоения новой адаптивной зоны может появиться в результате тех или иных сальтационных событий. К ним относятся существенное изменение хода развития в результате макромутации, приобретение адаптивного признака при горизонтальном переносе или целостного комплекса признаков при гибридизации (Боркин, Даревский, 1980), интерградации или симбиогенезе. Дальнейшая эволюция системы управления развитием будет происходить под влиянием отбора, стабилизирующего некоторые из новых вариантов развития.
Возрастание эффективности эволюционного процесса
С точки зрения СТЭ скорость эволюции должна уменьшаться по мере усложнения генетической системы организмов и замедления смены их поколений. Этому противоречит тот факт, что темпы эволюции ускоряются с возрастанием морфофизиологической сложности организмов. Удивительным моментом в истории Земли является так называемый «кембрийский взрыв»: возникновение в относительно короткий срок подавляющего большинства когда-либо существовавших типов животных, многие из которых вскоре вымерли. Едва ли причины этого события коренятся в малых ненаправленных популяционно-генетических изменениях. Более вероятно, что оно вызвано появлением механизмов, существенно повышающих эффективность макроэволюции. Например, это могло быть приобретение организмами билатеральной симметрии тела, связанное не столько с приобретением соответствующих генов, сколько с развитием эпигенетически зарегулированных вариантов развития клеток, тканей и органов, отвечающих за дифференциацию частей тела. Стабилизации отбором наиболее удачных планов строения тела должно было предшествовать широкое разнообразие их вариантов. В настоящее время за план строения отвечает хорошо зарегулированный эпигенетический комплекс, и значительных новаций у современных организмов не наблюдается.
Как указывал И. И. Шмальгаузен, следствием межгруппового отбора является вытеснение форм, отстающих в темпах и качестве приспособления к меняющейся среде. Такой процесс должен приводить к совершенствованию самого механизма эволюции (Шмальгаузен, 1968; Завадский, Колчинский, 1977). Ускорение эволюции у высокоорганизованных форм (Расницын, 1978) — результат действия такого межгруппового отбора.
По всей видимости, по мере усложнения организмов все большую роль в формировании их признаков играла эпигенетическая компонента. Биохимические признаки прокариот определяются в основном на генном уровне (это широко используется в генной инженерии). Что касается морфологических признаков высокоразвитых организмов, то они зависят от сложной системы эпигенетических взаимодействий. Существование механизмов, обеспечивающих целостность вида и проявляющихся в видовом мономорфизме, также может быть следствием «эволюции эволюции».
Повышению эффективности эволюционирования многих видов способствует подразделенность их популяционной структуры (Коновалов, 1974; Алтухов, 1989). Так, даже на относительно однородной территории зеленые (Bufo viridis) и серые (B. bufo) жабы формируют локальные группы, которые по многим признакам отличаются сильнее, чем подвиды или близкие виды (Шабанов, 2001, 2002). Значительное количество уровней разнообразия, на большинстве из которых может идти групповой отбор (Шабанов, Шабанова, 2001), повышает его эффективность.
Таким образом, есть основания предполагать, что магистральным путем эволюции является совершенствование эпигенетических механизмов регуляции индивидуального и исторического развития. Следовательно, эффективность перестройки ЭС под действием отбора может быть мерой эволюционного прогресса тех или иных групп. Важнейшие вехи этого пути — развитие регуляции экспрессии бактериальных цистронов; возникновение полового размножения; образование многотканевых организмов; появление механизма культурного наследования.
Некоторые итоги
На основании изложенного можно сделать ряд выводов.
1. Наше время — период кризиса второго эволюционного синтеза (СТЭ) и формирования третьего синтеза. Такой синтез призван описать пути возникновения адаптивных комплексов признаков, действие направляющих эволюцию и регулирующих ее темпы факторов, историческое совершенствование механизмов эволюции.
2. Отличительной особенностью третьего синтеза должен стать системный подход к эволюции, ее изучение на различных уровнях с определением интегрирующих механизмов для каждого из них.
3. Третий синтез должен объединить достижения различных относительно независимых концепций (классического дарвинизма, биологии индивидуального развития, современной генетики, эпигенетической теории М. А. Шишкина, экологической теории В. А. Красилова, неосальтационизма, концепций видового мономорфизма, «эволюции эволюции» и прерывистого равновесия и других).
4. Представление о скачкообразном характере видообразования согласуется с данными о реорганизации генома при возникновении вида и видовом мономорфизме. Экологическая и эпигенетическая теории, а также неосальтационизм объясняют причины неравномерности темпов эволюции, а концепция прерывистого равновесия сосредотачивается на его результатах.
5. Эпигенетическая теория М. А. Шишкина акцентирует внимание на ключевом уровне регуляции онтогенеза. Одним из ее следствий является новый подход к решению проблемы возникновения биологической целесообразности.
Какие данные могут служить подтверждением изложенных здесь взглядов? Поддается проверке предположение о том, что при развитии в контролируемых условиях онтогенез особей, происходящих из нарушенных (в том числе и антропическими воздействиями) экосистем, должен был менее стабильным, чем тех, которые происходят из эталонных местообитаний.
Для подтверждения эпигенетической теории представляется полезной проверка предположения о том, что аллели менделевского гена являются не последовательностями нуклеотидов, а заданными всей эпигенетической системой переключателями между стабилизированными вариантами нормы. Для этого желательно провести их экспериментальное получение и разрушение. Предположим, при отборе в дестабилизированной популяции сохраняются носители как одной, так и другой из двух альтернативных аберраций. Появятся ли в результате такого отбора менделирующие эпигенетические переключатели онтогенеза? Будет ли этот процесс эффективнее при условиях культивации, существенно отличающихся от нормальных (например, при выращивании дрозофил на нестандартной среде)? Приведет ли отбор на устойчивое воспроизведение гетерозиготного фенотипа (возникающего при скрещивании линий, у которых стабилизированы разные варианты нормы) к прекращению его менделевского расщепления? Один из вариантов такого отбора — сохранение обладателей гетерозиготного фенотипа из тех выводков, где их доля была максимальной.
Проверить некоторые положения данной статьи позволит изучение характера изменчивости мономорфной части генома у экспериментальных популяций, подвергнутых острому отбору на смену адаптивной нормы.
Главная цель, которую ставил перед собой автор — стимулировать обсуждение и экспериментальное изучение затронутых проблем.
Список цитированной литературы
Албертс Б., Брей Д., Льюис Дж., Рэфф М., Робертс К., Уотсон Дж. Молекулярная биология клетки: В 5-ти т. Т. 3. М.: Мир, 1987. 296 с.
Алтухов Ю. П. Генетические процессы в популяциях. М.: Наука, 1989. 328 с.
Беляев Д. К. О некоторых вопросах стабилизирующего и дестабилизирующего отбора // История и теория эволюционного учения. Л.: 1974. С. 76–84.
Берг Л. С. Труды по теории эволюции. Л.: Наука, 1977. 387 с.
Боркин Л. Я., Даревский И. С. Сетчатое (гибридогенное) видообразование у позвоночных // Журн. общ. биол. 1980. Т. 41. № 4. С. 485–506.
Воронцов Н. Н. Развитие эволюционных идей в биологии. М.: Изд. отд. УНЦ ДО МГУ, Прогресс-Традиция, АБФ, 1999. 600 с.
Гаузе Г. Ф. Экология и некоторые проблемы происхождения видов // Экология и эволюционная теория. Л.: Наука, 1984. С. 5–105.
Голубовский М. Д. Неканонические наследственные изменения // Природа. 2001. № 8. С. 3–9.
Голубовский М. Д., Чураев Р. Н. Динамическая наследственность и эпигены // Природа. 1997. № 4. С.16–25.
Грант В. Эволюционный процесс. Критический обзор эволюционной теории. М.: Мир, 1991. 488 с.
Гродницкий Д. Л. Эпигенетическая теория эволюции как возможная основа нового эволюционного синтеза // Журн. общ. биол. 2001. Т. 62. № 2. С. 99–109.
Завадский К. М., Колчинский Э. И. Эволюция эволюции (историко-критические очерки проблемы). Л., «Наука», 1977. 236 с.
Инге-Вечтомов С.Г. Прионы дрожжей и центральная догма молекулярной биологии // Вестн. РАН. 2000, Т. 70. №4. С.299–306.
Коновалов С. М. Субизолят как относительно жесткая система. Структура субизолята // Журн. общ. биол. 1974. Т. 35. № 6. С. 819–838.
Корочкин Л. И. Параллелизмы в молекулярной организации генома и проблемы эволюции // Молекулярные механизмы генетических процессов. М.: Наука, 1985. С. 132–146.
Красилов В.А. Филогения и систематика. Проблемы филогении и систематики. Владивосток, 1969. С. 12–30.
Красилов В. А. Теория эволюции: необходимость нового синтеза // Эволюционные исследования. Макроэволюция. Владивосток: ДВНЦ АН СССР, 1984. С. 4–17.
Кусакин О. Г., Дроздов А. Л. Филема органического мира. Ч. 1. Пролегомены к построению филемы. СПб.: Наука, 1994.
Левонтин Р. Генетические основы эволюции. М.: Мир, 1978. 352 с.
Михайлов К. Е. Типологическое осмысление «биологического вида» и пути стабилизации околовидовой таксономии у птиц
Назаров В. И. Учение о макроэволюции. На путях к новому синтезу. М.: Наука, 1991. 288 с.
Расницын А. П. Темпы эволюции и эволюционная теория (гипотеза адаптивного компромисса) // Эволюция и биоценотические кризисы. М.: Наука, 1987. С. 46–64.
Раутиан А. С. О природе генотипа и наследственности // Журн. общ. биол. 1993. Т. 54. № 2. С. 131–148.
Сойфер В. Н. Генетика. Зарождение хромосомной теории наследственности // История биологии с начала XX века до наших дней. М.: Наука, 1975. С. 295–313.
Татаринов Л. П. Параллелизм и направленность эволюции // Эволюция и биоценотические кризисы. М.: Наука, 1987. С. 124–144.
Тейяр де Шарден П. Феномен человека. М.: Наука, 1987. 240 с.
Шабанов Д.А. Мозаичное разнообразие зеленых жаб (Bufo viridis) в пределах однородного физико-географического региона // Вопросы герпетологии. М.: МГУ, 2001. С. 334–336.
Шабанов Д. А. Популяционное разнообразие видов рода Bufo в Харьковской области как предмет изучения и охраны // Вісн. Харківск. ун-ту. 2002. № 551. Ч. 2. С. 232–236.
Шабанов Д.А., Шабанова А.В. Уровни разнообразия зеленых жаб (Bufo viridis) // Фальцфейнівські читання. Херсон: Терра, 2001. С. 212–213.
Шапошников Г. Х. Динамика клонов, популяций и видов и эволюция // Журн. общ. биол. 1978. Т. 39. № 1. С. 15–33.
Шварц С. С. Современные проблемы эволюционной теории // Вопр. филос. 1967. № 10. С. 143–153.
Шварц С. С. Экологические закономерности эволюции. М.: Наука, 1980. 280 с.
Шишкин М. А. Индивидуальное развитие и эволюционная теория // Эволюция и биоценотические кризисы. М.: Наука, 1987. С. 76–124.
Шишкин М. А. Эволюция как эпигенетический процесс // Современная палеонтология. Методы, направления, проблемы, практическое приложение. М.: Недра, 1988. Т. 2. С. 142–169.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. М., 1938. 231 с.
Шмальгаузен И. И. Кибернетические проблемы биологии. Новосибирск, 1968. 224 с.
Шмидт-Ниельсен К. Размеры животных: почему они так важны? М.: Мир, 1987. 259 с.
Carson H. L. Speciation as a major reorganization of polygenic balances// Mechanisms of speciation. NewYork: Liss., 1982. P. 411–433.
Eklund M. W., Poysky F. T., Meyers J. A., Perloy G. A. Interspecies conversion of Clostridium botulinum type C to Clostridium novyi type A by bacteriophage // Science. 1974. Vol. 186. 456–458. (цит. по Грант, 1991, с. 189)
Goldschmidt R. The material basis of evolution. New Haven, London, Oxford, 1940. 436 p.
Haldane J. В. S. The cost of natural selection // J. Genet. 1957. Vol. 55. P. 511–524.
Harms, J. W. Wandlungen des Artgefüges. Heine, Tübingen. 1934. 212 p. (цит. по Goldschmidt, 1940)
Jenkin F. The origin of species Art. I.// North Brit. Rev. 1867, June. Vol. 46. P. 277–318.
Marx J. L. Instability in plants and the ghost of Lamarck // Science. 1984. Vol. 224. P. 1415–1416.
Waddington C. H. The Strategy of the Genes: a Discussion on Some Aspects of Theoretical Biology. London: Allen and Unwin, 1957. 262 p.