Хотя представленная здесь статья сохраняет, по крайней мере для ее авторов, некоторый интерес, терминология в ней нуждается в корректировке. Представления о возможных путях изменения популяционных систем уточнились, некоторые понятия и даже названия видов изменились. В частности, от термина "мероклональное наследование" пришлось отказаться в пользу "гемиклонального наследования" (подробнее - тут). Pdf статьи в исходном виде тут. Здесь, в сетевой публикации "мероклональность" исправлена на "гемиклональность" с выделением правки цветом.
Шабанов Д. А., Зиненко А. И., Коршунов А. В., Кравченко М. А., Мазепа Г. А. Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2006. – Випуск 3 (№ 729).
УДК 597.851(477.54)
Изучение популяционных систем зеленых лягушек (Rana esculenta complex) в Харьковской области: история, современное состояние и перспективы
Д. А. Шабанов, А.И.Зиненко, А. В. Коршунов, М. А. Кравченко, Г. А. Мазепа
Харьковский национальный университет имени В.Н. Каразина (Харьков, Украина)
В статье дан обзор среднеевропейских зеленых лягушек (Rana esculenta complex). Описано характерное для их межвидовых гибридов мерогемиклональное наследование (передача одного из геномов без рекомбинации). Описаны основные методы исследования этого явления, а также история изучения зеленых лягушек на территории Харьковской области. В Харьковской области зарегистрированы разные типы популяционных систем зеленых лягушек, в том числе включающие ди- и полиплоидные гибриды. Высказано предположение, что состав популяционных систем зеленых лягушек отражает результат их локальной эволюции и частотно-зависимого отбора гибридных линий, производящих разные типы гамет. Предложена схема, описывающая возможные преобразования популяционных систем в ходе их эволюции. Сформулированы перспективные направления дальнейших исследований разнообразия зеленых лягушек в Харьковской области.
Ключевые слова: Rana esculenta complex, триплоиды, тетраплоиды, гибридогенез, мерогемиклональное наследование, популяционные системы, Харьковская область.
D.A. Shabanov, O.I. Zinenko, O.V. Korshunov, M.O. Kravchenco, G.A. Mazepa. The study of population systems of green frogs (Rana ecsulenta complex) in Kharkiv region: history, modern condition and prospects. The article presents a review of Central European green frogs, which includes: a specific way of interspecies hybrids reproduction, during which one genome inherited without recombination – merohemiclonal inheritance; description of the main research methods of this phenomenon and the history of studying of green frogs in Kharkiv region. Different types of green frogs’ population systems have been registered in Kharkiv region, which also include di- and polyploid hybridogenetic frogs. There is an assumption that the combination of frogs’ population systems is the result of their local evolution and frequency-dependent selection of hybridogenetic lines, producing different types of gametes. A scheme of possible ways of population systems transformation is suggested. The prospective ways of further studying of green frogs’ diversity are also represented.
Keywords: Rana esculenta complex, triploids, tetraploids, hybridogenesis, merohemiclonal inheritance, populations, Kharkiv region.
Д. А. Шабанов, О.І.Зиненко, О. В. Коршунов, М. О. Кравченко, Г. А. Мазепа. Вивчення популяційних систем зелених жаб (Rana esculenta complex)в Харківської області: історія, сучасний стан та перспективи. У статті наведено огляд середньоєвропейських зелених жаб (Rana esculenta complex). Описано мерогеміклональне спадкування (передача одного з геномів без рекомбінації), яке є характерним для їхніх міжвидових гібридів. Описано основні методи дослідження цього явища, а також історія вивчення зелених жаб на території Харківської області. У Харківській області зареєстровані різні типи популяційних систем зелених жаб, у тому числі такі, що включають ди- і поліплоїдні гібриди. Висловлено припущення, що склад популяційних систем зелених жаб відбиває результат їхньої локальної еволюції й частотно-залежного добору гібридних ліній, що виробляють різні типи гамет. Запропоновано схему, що описує можливі перетворення популяційних систем у ході їхньої еволюції. Сформульовано перспективні напрямки подальших досліджень різноманіття зелених жаб у Харківській області.
Ключові слова: Rana esculenta complex, триплоїди, тетраплоїди, гібридогенез, мерогеміклональне спадкування, популяційні системи, Харківська область.
МероГемиклональное наследование у Rana esculenta
В состав комплекса среднеевропейских зеленых лягушек (Rana esculenta complex) входят три основные формы. Это прудовая лягушка (R. lessonae Camerano, 1882), озерная лягушка (R. ridibunda Pallas, 1771) и съедобная лягушка (R. esculenta Linnaeus, 1758). Все они встречаются в Харьковской области.
Зеленые лягушки относятся к жизненной форме береговых соскакивателей в воду, с которой связано возникновение основных особенностей отряда Anura. Благодаря своему широкому распространению, высокой численности и относительной доступности зеленые лягушки принадлежат к числу наиболее хорошо изученных и известных животных, а также распространенных экспериментальных объектов. Тем не менее, до второй половины XXвека таксономический статус входивших в состав этой группы форм неоднократно перематривался. Это связано и со значительной морфологической изменчивостью этих животных, и с характерным для них феноменом межвидовой гибридизации, который был открыт лишь в ходе экспериментов польского гидробиолога Л. Бергера (Berger, 1964 и последующие работы). Так, Бергер обнаружил, что при скрещивании R. lessonae и R. ridibunda потомство состоит из особей, которых следует относить к R. esculenta. При скрещивании таких гибридов с R. lessonae потомство также состояло исключительно из R. esculenta.
Объяснить эти результаты с помощью гипотезы клонального наследования смог австрийский биолог Хейнц Тюннер, использовавший для разделения различных форм лягушек электрофорез белковых маркеров (Tunner, 1974). Тюннеру и другим исследователям (например, Боркин и др., 1987) удалось доказать, что один из родительских геномов удаляется из клетки перед мейозом, а другой удваивается и передается в гаметы клонально, без рекомбинации. Схематичное объяснение описанных результатов Бергера, а также некоторых других феноменов, возникающих при скрещиваниях внутри Rana esculenta complex, приведено в табл. 1 (в таблице буквами обозначены геномы L – прудовой и R – озерной лягушек). Генотип LL соответствует прудовой лягушке, RR — озерной, а LR — их гибриду, съедобной лягушке.Один из геномов гибридных лягушек (выделенный полужирным шрифтом) передается клонально, без рекомбинации с другим геномом. Другой, неклональный геном используется в течение жизни гибридной особи, а перед мейозом элиминируется.
Таблица 1. Схемы различных типов воспроизводства в Rana esculenta complex. Геном прудовой лягушки обозначен как L, а озерной — как R; геном, элиминируемый в ходе гаметогенеза, перечеркнут
|
|
А. Образование гибрида при скрещивании родительских видов |
Б. Воспроизводство гибрида при его скрещивании с R. lessonae |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
В. Воспроизводство гибрида при его скрещивании с R. ridibunda |
Г. Гибридолиз — выщепление родительского вида при скрещивании гибридов |
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Как видно при сравнении вариантов Б и В в табл. 1, обитая с одним из родительских видов, гибридные лягушки производят гаметы другого вида (комплементарные гаметы), что позволяет им воспроизводить гибридов и в следующем поколении. Не во всех случаях это правило выполняется, но те формы гибридов, которые производят гаметы того же родительского вида, с которым они скрещиваются, будут исчезать из популяции в результате таких скрещиваний. Формы гибридов, производящих комплементарные родительскому виду гаметы, напротив, будут воспроизводиться успешно, и им будет благоприятствовать отбор.
При скрещивании друг с другом гибридных лягушек, производящих гаметы одного и того же родительского вида, может происходить выщепление этого вида в потомстве (см. табл. 1, Г). Такое явление получило название гибридолиза (от лат. hybrida —помесь и греч. lysis — разложение, распад). Особи, образующиеся в результате гибридолиза, обычно демонстрируют пониженную жизнеспособность, что может быть следствием изменений, накапливающихся в клональном геноме во время его передачи из поколения в поколение без рекомбинации.
Для обозначения способа воспроизводства, зарегистрированного у зеленых лягушек, предложен целый ряд терминов. Впервые аналогичный феномен был описан у мексиканских карпозубых рыб рода Poeciliopsis (Schultz, 1969) под названием «гибридогенез» (от hybrida и греч. genesis — возникновение). Это название не очень удачно, так как суть самого феномена состоит не столько в возникновении гибридов, сколько в их воспроизводстве за счет одного из родительских видов, некоем «гаметном (геномном) паразитизме» (Günther, 1979). Поскольку геном одной из родительских форм используется лишь временно, Л.Я. Боркин и И.И. Даревский (1980) предложили называть его «кредитогенезом» (от лат. creditum — ссуда). Различными авторами используются также термины «клептогенез» (от лат. klepto — ворую; Dubois, Günther, 1982)), а также «гемиклональное» (от лат. hemi- —полу- и греч. clon —отпрыск, ветвь) и «мероклональное» (от греч. meros—часть, доля) наследование. С нашей точки зрения, оптимальным является термин мероклональное наследование, как носящий самый общий характер. Ему не противоречат ни наследование одного или двух геномов у триплоидных гибридов, ни зарегистрированные случаи рекомбинации между геномами (см. ниже). Некоторые из этих терминов совместимы друг с другом: например, можно говорить о мерогемиклональном наследовании при кредитогенезе.
В связи с изложенным возникает естественный вопрос: следует ли использовать для обозначения гибридных лягушек видовое название R. esculenta? В пользу придания съедобным лягушкам самостоятельного статуса свидетельствует то, что гибриды не возникают раз за разом при скрещивании родительских видов, а устойчиво воспроизводятся в ряду поколений, передавая клональный геном и приобретая новые особенности в ходе эволюции. Гибриды имеют свой ареал, выходящий за пределы пересечения ареалов родительских видов. Кроме того, известно, что гибридизация родительских видов в природе случается довольно часто, однако продукты такой гибридизации обычно обладают пониженной жизнеспособностью и часто стерильны, поэтому проигрывают гибридам, воспроизводящимся путем гибридогенеза (Plötner, 2005). Существенным обстоятельством является то, что гибриды могут образовывать чистые популяции, где существуют без скрещивания с родительскими видами. К слову, R. esculenta была описана Карлом Линнеем существенно раньше, чем более поздние исследователи описали ее родительские виды R. lessonae и R. ridibunda. В данном случае, как и во многих других, богатство проявлений жизни не укладывается в прокрустово ложе однозначных правил и унифицированных категорий. Съедобная лягушка не является ни обычным гибридом, ни настоящим видом. Чтобы придать определенный таксономический статус таким гибридным формам, была предложена таксономическая категория клептон (Dubois, Günther, 1982). В таком случае название распространенных в Харьковской области гибридных лягушек следует записывать как Rana klepton esculenta (сокращенно R. kl. esculenta). Хотя в данной статье гибридные формы обозначаются просто как R. esculenta, следует учитывать, что это название применяется в отношении особой формы, а не обычного вида.
Разнообразие популяционных систем зеленых лягушек
Несмотря на экологическую специфичность R. lessonae, R. ridibunda и R. esculenta, все они способны населять одни и те же местообитания, будучи привязаны к берегам разнообразных пресных водоемов. Во время нереста, в репродуктивный сезон они могут образовывать общие группы размножения, включающие представителей разных форм. В них происходит не только воспроизводство названных форм, но и их гибридизация. Поскольку термин «популяция» не относится к группам, включающим представителей разных видов, совокупности зеленых лягушек, населяющих определенные местообитания и совместно участвующих в размножении, принято называть «популяционными системами».
После открытия Л. Бергера был описан целый ряд типов популяционных систем зеленых лягушек, отличающихся как по своему составу, так и по характеру гамет, продуцируемых гибридными формами. Самая общая из предложенных классификаций популяционных систем Rana esculenta complex (Uzzell, Berger, 1975; Лада, 1995) включает следующие 7 их типов:
L-тип — обычные популяции R. lessonae, прудовой лягушки;
R-тип — обычные популяции R. ridibunda, озерной лягушки;
E-тип — так называемые «чистые» популяции гибридной формы, R. esculenta;
L-R-тип — популяционные системы из озерной и прудовой лягушек, размножающихся без образования гибридов;
L-E-тип — популяционные системы из R. lessonae и гибридов (см. второй случай в табл. 1);
R-E-тип — аналогичные предыдущему случаю популяционные системы, в которых гибриды сосуществуют с R. ridibunda;
R-E-L-тип — смешанные системы, включающие оба родительских вида и их гибриды.
Для значительной части ареала (Западной Европы) «чистые» популяционные системы E-типа включают в себя значительную долю триплоидных гибридов. В популяционном воспроизводстве триплоиды могут заменять родительский вид, производя гаметы, дополняющие гаметы диплоидных гибридов. Такие системы рекомендуют относить к E-t-типу и не считать «чистыми», рассматривая как вид или клептон только диплоидных Rana esculenta (Plötner, 2005).
Следует отметить, что приведенная классификация не описывает все разнообразие популяционных систем зеленых лягушек. Эти системы могут отличаться друг от друга еще по нескольким параметрам, подробное рассмотрение которых выходит за рамки данной статьи. В их числе:
— количественное соотношение различных форм;
— половой состав каждой из форм, в первую очередь — гибридов, связанный с тем, что клональные геномы могут быть как мужскими, так и женскими;
— наличие или отсутствие особей с рекомбинированными геномами;
— наличие или отсутствие гибридов, отличающихся по плоидности (количеству геномов) и их составу;
— характер гамет (количество и состав геномов), производимых различными формами гибридов;
— состав родительских форм, каждая из которых может быть представлена различными геномами или даже криптическими видами (Plötner, 2005; Боркин и др., 2004).
Необходимо отметить, что за последние годы количество описанных таксонов — как видов, так и гибридных форм — европейских зеленых лягушек существенно увеличилось. По последней сводке (Plötner, 2005), в группу R. ridibunda входят также R. bedriagae Саmеranо, 1882, R. terentievi Mezhzherin, 1992, R. cretensis Beerli, Hotz, Tunner, Heppich & Uzzell, 1994 и R. epeirotica Schneider, Sofianidou & Kyriakopoulou-Sklavounou, 1984. ВгруппуR. lessonae входятR. bergeri Günther, 1985, R. shqiperica Hotz, Uzzell, Günther, Tunner & Heppich, 1987, R. perezi Seoane, 1885, иR. saharica Boulenger in Hartert, 1913. Не сближаются ни с одним из «основных» видов, но входят в тот же комплекс виды R. cerigensis Beerli, Hotz, Tunner, Heppich & Uzzell, 1994 и R. caralitana Arikan, 1988, а также некоторые другие, исследуемые в настоящее время, но еще не получившие свое название формы. Клептонами, кроме R. esculenta, являются R. hispanica Bonaparte, 1839 (результат скрещивания R. bergeri и R. ridibunda) и R. grafi Crochet, Dubois, Ohler & Tunner, 1995 (гибрид R. реrеzі и R. ridibunda). Можно предположить, что дальнейшее изучение этой удивительной группы животных приведет к еще большему увеличению числа включаемых в нее форм.
В ходе изучения популяционных систем зеленых лягушек следует учитывать, что они не ограничены жесткими рамками. Для этой группы животных характерно образование метапопуляций — совокупностей из многих постоянных или временных групп размножения, между которыми происходит обмен особями. Отдельные группы размножения могут отличаться друг от друга по своему составу, и благодаря миграции влиять на другие группы в рамках общей метапопуляции.
Из сказанного становится ясно, почему изучение зеленых лягушек привлекает большее внимание в европейской герпетологии, чем любая другая проблема.
Методы изучения гибридогенного комплекса лягушек
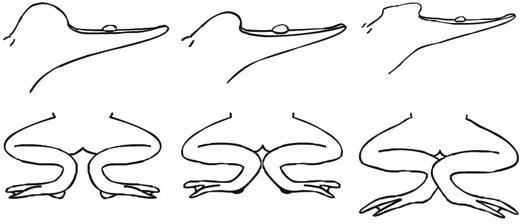
Для диагностикиформ лягушек различными исследователями был использован целый комплекс методов. Три основные формы достаточно хорошо отличаются друг от друга морфологически (табл. 2). Увы, существование индивидуальной изменчивости, наличие триплоидов и тетраплоидов, а также частично рекомбинантных форм делает морфологическую дискриминацию всего комплекса невозможной. Для узнавания трех основных форм Rana esculenta complex в полевых условиях полезным оказывается акустический критерий — разные формы лягушек отличаются по голосу. Кроме того, R. lessonae, R. ridibunda и R. esculenta отличаются (табл. 2) особенностями местообитаний, поведением, окраской (Лада, 1995, Broska, 1982, Ананьева и др., 1998) и даже запахом (наши данные). По большинству из этих параметров Rana esculenta обладает промежуточными между R. lessonae и R. ridibunda значениями признаков. В ряде работ (например, Plenete.a., 2000; Negovetice.a., 2001) зарегистрированы существенные экофизиологические отличия между названными формами зеленых лягушек. Экологические отличия между членами Rana esculenta complex приводят к определенному разделению их экологических ниш и смягчают межвидовую конкуренцию. Ниши головастиков разделены существенно меньше. Воспроизводство в рамках общих популяционных систем оказывается фактором, препятствующим более глубокому разделению ниш R. esculenta и ее родительских видов (Holenweg Peter, Reyer, 2002).
Приведенные в табл. 2 данные позволяют в полевых условиях достаточно надежно идентифицировать родительские виды и диплоидных гибридов, однако не подходят для идентификации полиплоидных гибридов. Изучение размера эритроцитов и икринок может дать информацию о числе хромосомных наборов (клетки три- и тетраплоидов крупнее), но не позволяет определить происхождение геномов. Изучение кариотипов, проводимое классическими методами (например, Манило, 2005), также дает лишь общую информацию о хромосомном наборе (позволяя регистрировать полиплоидные и анеуплоидные клетки), но не дает надежных методов для идентификации видовой принадлежности изучаемых хромосом. Неоднократно публиковавшиеся данные о морфологических отличиях хромосом R. lessonae, R. ridibunda противоречивы (например, Сурядная, 2003 и Сурядна, 2005) и не предоставляют надежного основания для определения генотипа изучаемых особей. Идентификация отдельных геномов возможна лишь с помощью технологий дифференциального окрашивания хромосом (например, Miura e.a., 1997).
Таблица 2. Сравнение по некоторым признакам трех основных форм в составе Rana esculenta complex (Лада, 1995 и др.)
|
Признак |
Прудовая лягушка (R. lessonae Camerano, 1882) |
Съедобная лягушка (R. esculenta Linnaeus, 1758). |
Озерная лягушка (R. ridibunda Pallas, 1771) |
|
Размер и форма внутреннего пяточного (метатарзального) бугра и длина первого пальца |
|
||
|
Относительная длина голени |
|||
|
Окраска резонаторов у самцов |
Белая или желтовато-белая |
Светло-серая |
Темно-серая — черная |
|
Общий тон окраски |
Обычно зеленая, иногда яркая светло-зеленая; наличие брачной окраски; желтый цвет на задних конечностях |
Промежуточное между родительскими видами; желтый цвет на задних конечностях |
От зеленой до почти черной |
|
Запах |
Слабый запах |
Сильный запах |
Резкий специфичный запах |
|
Брачная песня самца (после «запевки») |
Стрекочущая |
Промежуточная по характеру между родительскими |
Раскатистая, напоминающая хохот |
|
Поведение при опасности |
Проплывают некоторое расстояние под водой и выныривают |
Промежуточное между родительскими видами |
Затаиваются на дне |
|
Поведение самцов относительно конкурентов |
Весьма агрессивное |
Агрессивное |
Относительно нейтральное |
|
Зимовка |
На суше |
Вместе с родительским видом |
В воде |
|
Предпочитаемые местообитания |
Небольшие водоемы лесной зоны, вне периода нереста живет на суше |
Различные, кроме крайних типов, характерных для родительских видов |
Крупные водоемы открытых ландшафтов |
Значительный прогресс в изучении мерогемиклонального наследования и разнообразия зеленых лягушек был связан с изучением белковых маркеров методом гель-электрофореза. Этот метод позволяет также опознавать некоторые рекомбинантные особи (у которых рекомбинация затронула рассматриваемые маркеры), а благодаря эффекту дозы гена — определять триплоидов среди гибридов (Günther, 1975).
Удобным и мощным методом изучения гибридогенного комплекса оказался метод проточной ДНК-цитометрии, позволяющий определить количество ДНК, приходящееся на клетку (Боркин и др., 1987). Этот метод основан на измерении УФ-флуоресценции ДНК одиночных клеток (эритроцитов или сперматозоидов). Ошибка измерения при таком исследовании значительно меньше разницы в размере генома между родительскими видами, что позволяет не только достоверно определять количество и видовую принадлежность геномов взрослых особей, но и устанавливать, какие геномы несут сперматозоиды. Поскольку вес генома R. ridibunda составляет около 8 пг (пикограмм), а R. lessonae — около 7 пг, ясно, например, что особь с весом генома 22 пг является триплоидом LLR. Рекомбинантные особи, имеющие измененное количество ДНК в геноме, могут быть опознаны с помощью этого метода с существенно меньшей долей уверенности.
В последние годы получила распространение идентификация отдельных генотипов зеленых лягушек по характерным последовательностям микросателлитной ДНК (например, Christiansen e.a., 2005). Вероятно, в дальнейшем именно этот способ станет основным, используемым для изучения как зеленых лягушек, так и многих других групп организмов.
Интерес к гибридогенному комплексу европейских зеленых лягушек, вспыхнув около сорока лет назад, до сих пор не угасает. Изучением различных аспектов данной проблемы занимается ряд европейских университетов, музеев и лабораторий. На сегодня было проведено уже 3 международных совещания, посвященных проблемам изучения Rana esculenta complex. Два основных центра изучения зеленых лягушек, расположенных на территории бывшего СССР, находятся в Санкт-Петербурге и Киеве. В ЗИН и ЦИН РАН используется, в первую очередь, метод проточной ДНК-цитометрии. В Институте зоологии им. И.И. Шмальгаузена НАНУ и Зоомузее ННПМ НАНУ, кроме прочего, проводится электрофорез белковых маркеров, кариоанализ и иммунологические исследования. В последнее время полевые, морфометрические и цитогенетические исследования начаты в Харьковском университете авторами настоящей работы.
Как справедливо отметил Г.А. Лада (1995), «в проблеме среднеевропейских зеленых лягушек в один тугой узел связаны вопросы морфологии, экологии, этологии, систематики, цитологии, биохимии, генетики, палеонтологии и теории эволюции»; возможно, к этому списку следовало бы добавить также биологию индивидуального развития и биогеографию.
Изучение зеленых лягушек в Харьковской области
Изучение населяющих Харьковскую область зеленых лягушек началось еще в середине XIXвека (Чернай, 1852). Еще ранее профессор Харьковского университета И.А. Криницкий (1837) описал из окрестностей Пятигорска вид Rana dentex Krynicki, 1837, в настоящее время сведенный в синонимы R. ridibunda. Один из этапов истории изучения лягушек связан с деятельностью А. М. Никольского, основателя школы герпетологии в России и СССР. Большая часть его научной деятельности прошла в Харькове, в том числе в Харьковском университете. Столкнувшись с наличием переходов между разными формами зеленых лягушек, Никольский рассматривал их как подвиды одного вида R. esculenta. Следует, впрочем, отметить, что даже в харьковский период своей жизни А.М. Никольский изучал в основном не местный материал, а сборы со значительной части территории бывшей Российской империи.
После открытия гибридной природы R. esculenta к зеленым лягушкам было привлечено внимание многих исследователей. Киевский герпетолог С. В. Таращук (1985) предложил мультипликативный индекс, предназначенный для дискриминации различных форм зеленых лягушек. Этот индекс был применен сотрудником Харьковского университета В. И. Ведмедерей (1984) для описания разнообразия лягушек из Харьковской области еще до публикации сведений об этом индексе в печати.
Изучение лягушек этого региона биохимическими методами начато сотрудниками Зоологического института РАН в 1989 году при участии А.М. Рудика. В 1995 и 1996 годах тамбовский герпетолог Г.А. Лада посетил Харьковскую область и описал на Иськовом пруду в окрестностях биостанции Харьковского университета в Гайдарах крупную «чистую» популяцию диплоидных R. esculenta (Лада, 1998).
Харьковская область находится на границе ареала одного из родительских видов, R. lessonae. Ареал R. esculenta в общих чертах совпадает с ареалом R. lessonae (R. ridibunda распространена намного шире), но кое-где выходит за его пределы. В Харьковскую область R. lessonae заходит лишь на ее северо-западе (Краснокутский и Богодуховский районы) и юго-западе (Зачепиловский район, урочище Русский Орчик). Возможно, существуют локальные популяции R. lessonae и в других северных районах области, но в настоящее время их существование достоверно не доказано. В то же время ареал R. esculenta выходит за эти пределы в юго-восточном направлении, распространяясь до Изюмского района Харьковской области и простирается далее вниз по течению Северского Донца. В этой части ареала гибридные лягушки существуют в основном в составе популяционных систем R-E-типа.
По предварительным данным, основанным на данных ДНК-цитометрии и полевых наблюдений, на сегодняшний день в Харьковской области зарегистрировано 6 разновидностей популяционных систем, относящихся к R-, R-E- и R-E-L-типам:
I. особи обоих полов с генотипом RR (R-тип);
II. особи обоих полов с генотипами RR и LR, а также одиночные неполовозрелые особи LL, вероятно, выщепляющиеся в результате гибридолиза (R-E-тип);
III. особи обоих полов с генотипами RR, LRR, LR, LLR, а также одиночные неполовозрелые особи LL и LLRR (R-E-тип с триплоидами);
IV. особи обоих полов с генотипами LR, а также одиночные половозрелые RR (так называемый «чистый» E-тип, который, по всей видимости, связан переходами с R-E-типом);
V. самцы LR и одиночные половозрелые особи RR (эту систему можно назвать Em-типом);
VI. особи обоих полов с генотипами RR, LR и LL (R-E-L-тип).
Полиплоидные гибриды достаточно широко распространены в Западной Европе. На территории бывшего Советского Союза, несмотря на систематические поиски, к 2001 году удалось зарегистрировать лишь несколько отдельных особей. В 2002 году в ходе совместной работы сотрудников Зоологического института РАН и Цитологического института РАН (Санкт-Петербург), а также Харьковского университета в выборках из Харьковской области была зарегистрирована значительная доля триплоидных гибридов (Borkin e.a., 2004). В настоящее время количество исследованных с помощью цитометрии представителей Rana esculenta complex из Харьковской области составило 813 особей из 48 пунктов (Боркин и др., 2005). Для самых распространенных форм зеленых лягушек определен состав продуцируемых самцами гамет. Получена предварительная оценка распределения на изученной территории различных форм зеленых лягушек и типов их популяционных систем (Borkin e.a., 2004; Боркин и др., 2005).
Результаты исследований в Харьковской области говорят о систематическом возникновении полиплоидов в популяционных системах зеленых лягушек, населяющих водоемы, расположенные в бассейне Северского Донца. В некоторых из них доля триплоидов составляет около 40% от общего количества гибридных лягушек. Обнаруженная зона распространения триплоидов отделена от подобных регионов значительным разрывом: ближайшие районы, где ранее были сделаны массовые находки полиплоидных особей, находятся на расстоянии около 1000 км (в Польше) и около 1500 км (в западной Венгрии).
Особый интерес представляет недавняя находкатриплоидных гибридных лягушек в водоемах поймы Северского Донца в Ростовской области (Borkin e.a., 2006). Доля триплоидов в этом районе даже выше, чем в Харьковской области. Вероятно, зарегистрированные в Харьковской и Ростовской областях феномены тесно связаны и относятся к единому центру разнообразия зеленых лягушек, расположенному в бассейне Северского Донца.
Rana esculenta-комплекс в окрестностях биостанции ХНУ и на территории НПП «Гомольшанские леса»
Для характеристики сложности задачи, решаемой при изучении разнообразия зеленых лягушек, можно рассмотреть состав наиболее изученной метапопуляции из Харьковской области (табл. 3). Она расположена в Змиевском районе, в окрестностях биостанции ХНУ и на территории Национального природного парка «Гомольшанские леса» (пойма Северского Донца и р. Гомольши, временные водоемы и пруды в Коряковом и Иськовом ярах в окрестностях с. Гайдары, а также в Добрицком яру в окрестностях с. Великая Гомольша). На этой территории размещены локальные популяционные системы зеленых лягушек, связанные путями миграций. На настоящее время сотрудниками ЗИН и ЦИН РАН определены генотипы более, чем 350 особей из этой метапопуляции. Для 38 диплоидных гибридных самцов, а также 3 триплоидов LRRбыл определен состав спермиев (Боркин и др., 2005). Кроме того, накоплен значительный объем данных по распределению и численности разных форм, которые были определены по морфологическим и биоакустическим признакам.
Среди диплоидных самцов R. esculenta было зарегистрировано три типа образования гамет (Боркин и др., 2005), представленных у приблизительно равного количества особей. Это самцы, производящие только сперматозоиды с геномом R, самцы, производящие только сперматозоиды с геномом L, и самцы, производящие смесь сперматозоидов с геномами R и L. Наибольший интерес представляют особи последней группы, способные производить одновременно гаметы двух разных видов. Цитогенетический механизм, обеспечивающий такой тип гаметогенеза, остается загадочным. Одним из его вероятных объяснений могла бы быть мозаичность таких особей, в том случае, если они состоят из смеси клеток с разными генотипами. Подтверждением того, что такая мозаичность возможна, могут быть недавно найденные в Житомирской области миксоплоиды зеленых лягушек (Манило, 2005). Возможно, самцы зеленых лягушек могут производить также и диплоидные спермии, однако с помощью проточной цитометрии их невозможно отличить от других диплоидных клеток, которые могут попадать в состав исследуемой пробы (С.Н. Литвинчук, личное сообщение).
Таблица 3. Состав метапопуляции зеленых лягушек в окрестностях биостанции ХНУ и на территории НПП «Гомольшанские леса», а также гаметы, производимые различными формами («+++» — более 10% особей соответствующей категории; «++» — от 1 до 10%; «+» — 1% или менее; «?» — данные отличаются для разных локальных популяционных систем)
|
|
RR |
LR |
LLR |
LRR |
LL |
LLRR |
||
|
Неполовозрелые |
+++ |
+++ |
+ |
+++ |
+ |
+ |
||
|
Самки |
+++ |
++ (?) |
+ |
++ (?) |
Возможно, не доживают до репродуктивного возраста |
|||
|
Самцы |
+++ |
+++ |
нет данных |
++ (?) |
||||
|
Гаметы самцов |
R |
L |
R |
L, R |
нет данных |
R |
||
|
Гаметы самок |
нет данных |
|||||||
В составе указанной метапопуляции можно выделить отдельные, особо интересные локальные популяционные системы. Одна из них населяет Иськов пруд, расположенный возле с. Гайдары. Как уже указывалось, Г.А. Лада (1998) исследовал этот пруд в 1995 и 1996 году и обнаружил там крупную «чистую» популяцию R. esculenta, отметив, что в ней идет эффективное популяционное воспроизводство. В 2001 году Иськов пруд был спущен, однако уже в 2002 году заполнен снова. В 2003 году в нем наблюдались особи R. esculenta обоих полов. Однако в 2005 году во время нереста на Иськовом пруду регистрировались только самцы R. esculenta, а из самок удалось найти лишь 2 особи R. ridibunda. Эти данные, а также результаты изучения других популяционных систем, напоминающих Иськов пруд, позволило выдвинуть предположение, что «чистые» популяционные системы R. esculenta, состоящие только из самцов, могут существовать благодаря приходящим в них из других местообитаний самкам R. ridibunda (Коршунов, 2005). Аналогичное объяснение выдвигалось в литературе для объяснения существования на Западной Украине чистых популяций R. esculenta, состоящих из одних самок (Межжерин и др., 2005).
Согласно нашей гипотезе, подавляющее большинство скрещиваний в Иськовом пруду (а также в других популяционных системах Em-типа) соответствует схеме, показанной в табл. 4.
Можно предположить, что ранее в Иськовом пруду обитали гибридные лягушки, клонально передающие разные типы геномов. При разрушении устойчивой популяционной структуры вследствие спуска пруда и смешивании с лягушками из других популяционных систем из различных клональных линий сохранилась только одна, с мужским клональным геномом R. lessonae. Самцы, передающие геном LY, при любых скрещиваниях с самками, продуцирующими R-гаметы, производят таких же самцов (а при скрещиваниях с гибридными самками, продуцирующими L-гаметы, возникают гибридолизные R. lessonae, которые не доживают до половозрелости). Популяционная система Иськова пруда оказалась в эволюционном тупике. Даже если в нее попадают иные самцы лягушек, из-за избытка самцов LYRX они имеют весьма невысокие шансы оплодотворить имеющихся самок. Согласно излагаемой гипотезе, эффективная численность популяционной системы зеленых лягушек в Иськовом пруду критически низка, и ее воспроизводство зависит от особей-мигрантов. Результатом этого должна стать деградация данной популяционной системы, которую следует ожидать в ближайшие годы.
Таблица 4. Гипотетический механизм воспроизводства в популяциях «Em-типа» на примере Иськова пруда в с. Гайдары. Индекс yсоответствует геному, содержащему мужскую (гетерогаметную) половую хромосому, а X — женскую (гомогаметную)
|
|
||||||||||||||||||||||||||||
Динамика и стабильность различных типов популяционных систем
Отраженная в табл. 4 гипотеза иллюстрирует то, что разные типы популяционных систем зеленых лягушек могут отличаться по своей стабильности. Это означает, что такие системы следует изучать не только в статике, но и в динамике. Рядом авторов зарегистрированы различные преобразования популяционных систем (например, Holenweg Peter, 2001; Vorburger, Reyer, 2003; Межжерин и др., 2005), а также использованы методы математического моделирования происходящих в них процессов (Hellriegel, Reyer, 2000; Reyer e.a., 2004 и др.). С нашей точки зрения, описанные процессы могут рассматриваться, как части общей последовательности возможных преобразований популяционных систем. Для изучения их разнообразия мы предлагаем общую схему изменения популяционных систем Rana esculenta complex (рис. 1). С нашей точки зрения, такая схема может быть полезна для моделирования динамики описываемых процессов и их изучения в полевых условиях.
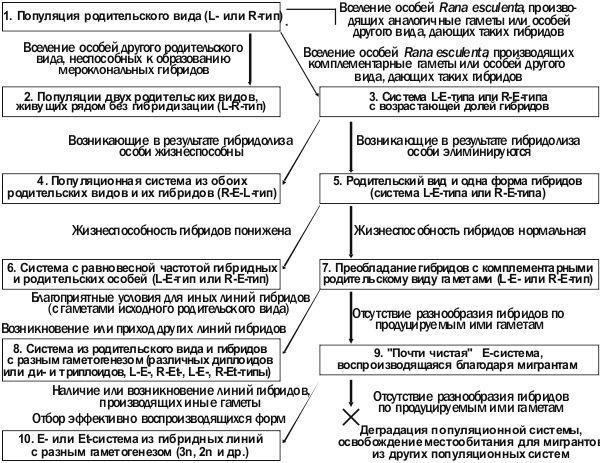
Рис. 1. Возможная последовательность преобразований различных типов популяционных систем Rana esculenta complex
Перед подробным рассмотрением предложенной схемы необходимо указать, что она не предусматривает учет трех обстоятельств, существенных для анализа динамики популяционных систем зеленых лягушек в каждом конкретном случае. Во-первых, это отличия между R. lessonae и R. ridibunda, отношения которых с гибридными формами не вполне симметричны. Например, в силу предпочтения самцами лягушек более крупных самок, скрещивание ♀ R. ridibunda x ♂ R. lessonae намного более вероятно, чем ♀ R. lessonae x ♂ R. ridibunda. Аналогичная избирательность проявляется и в скрещиваниях, в которых участвует R. esculenta (Abt, Reyer, 1993). Такие и иные проявления сексуальной избирательности могут оказывать существенное влияние на состав популяционных систем лягушек (Som e.a., 2000). Во-вторых, это различия популяционных систем по половому составу форм в их составе и половой принадлежности клональных геномов. Так, женский клональный геном может передаваться и самкам, и самцам гибридных форм, а мужской — только самцам. В-третьих, в предложенной схеме не учтена специфика местообитаний, в разной степени соответствующая запросам трех форм в составе Rana esculenta complex и тем самым влияющая на их соотношения в популяционных системах.
В качестве исходной точки эволюции популяционных систем следует рассмотреть популяцию одного из родительских видов, куда вселяются представители другого вида или мерогемиклональные гибриды (рис. 1, этап 1). Если вселяющиеся особи другого вида не смогут образовывать с видом-хозяином мерогемиклональные гибриды, но займут свою нишу, популяционная система перейдет в L-R-тип (рис. 1, этап 2). Вселение другого родительского вида, образующего с видом-хозяином мерогемиклональные гибриды эквивалентна вселению самих этих гибридов. Гибриды могут быть представлены различными линиями, отличающимися по тому, какие клональные геномы ими передаются. Эти клональные геномы могут отличаться по видовой принадлежности, полу и генетическим особенностям.
Успешно воспроизводиться в популяции родительского вида смогут только те линии гибридов, которые будут производить комплементарные гаметы (например, гаметы R. lessonae при вселении впопуляцию R. ridibunda, см. варианты Б и В в табл. 1). Линии, производящие гаметы, аналогичные преобладающему в популяционной системе родительскому виду, попросту растворятся при скрещиваниях. Итак, популяции родительского вида, доминирующего в том или ином регионе, действуют как фильтр, оставляющей одну из гибридных форм.
При совместном обитании преобладающего по численности родительского вида и гибрида, производящего комплементарные гаметы, доля гибридных особейбудет возрастать (рис. 1, этап 3; Vorburger, Reyer, 2003 и другие работы). Причина этого — в том, что потомство от скрещивания особей родительского вида и гибридов будет полностью гибридным. По мере возрастания доли гибридов будет увеличиваться количество особей другого родительского вида, выщепляющегося в результате гибридолиза. Если продукты гибридолиза окажутся жизнеспособными и плодовитыми, популяционная система преобразуется в R-E-L-тип (рис. 1, этап 4). Если гибридолизные особи будут гибнуть, возрастание доли гибридов приведет к увеличению генетического груза в популяции. Тем не менее, по мере скрещивания особей родительского вида с гибридами, доля последних будет возрастать и далее (рис. 1, этап 5).
Как показывают полевые наблюдения, во многих случаях в естественных популяционных системах достигается определенная стабильная доля гибридов (рис. 1, этап 6). Вероятно, это связано с различными проявлениями гибридного дисгенеза: повышенной смертностью гибридов, нарушениями их развития, снижением фертильности и повышением доли анеуплоидных гамет, из которых развивается нежизнеспособное потомство (Christiansen e.a., 2005 и др.). В таком случае равновесное отношение между гибридами и родительскими особями будет отражать их относительную жизнеспособность (Holenweg Peter, 2001; Reyer e.a., 2004).
Как уже было указано, при вселении гибридов в популяцию родительского вида отбор должен способствовать линиям, производящим комплементарные родительскому виду гаметы. Однако отбор линий гибридов носит частотно-зависимый характер, и по мере увеличения доли гибридов одной линии благоприятные возможности откроются для других форм гибридов. Так, в популяции, где преобладают гибриды, производящие гаметы L (рис. 1, этап 7), отбор может способствовать распространению диплоидных и триплоидных особей, производящих гаметы R (табл. 3). Равновесную долю представителей этих линий в популяции определят, кроме прочего, уровень жизнеспособности и особенности гаметогенеза гибридных особей, получивших клональный геном Lот одного из родителей, и клональный геном R— от другого.
Таким образом, одним из результатов эволюции популяционной системы может быть переход к системе с несколькими цитогенетическими формами гибридов (рис. 1, этап 8). Однако если он по каким-то причинам (отсутствие необходимых линий гибридов или их нежизнеспособность) не произойдет, комплементарные исходному родительскому виду гибриды могут вытеснить его полностью или почти полностью. Результатом этого может стать популяционная система, воспроизводство которой связано с притоком мигрантов и эффективная численность которой снижена до критического уровня (рис. 1, этап 9).
Отбор в таких популяционных системах будет способствовать распространению линий гибридов, производящих иные типы гамет. Если в популяционной системе сложится вариант соотношения разных форм лягушек, отличающихся по своему генотипу и по продуцируемым гаметам, который обеспечивает эффективное воспроизводство, она сохранится; если нет — деградирует. Через какое-то время освободившееся местообитание будет заселено мигрантами из других локальных популяций. Вероятно, Иськов пруд находится на этом этапе эволюции популяционной системы лягушек. Кроме того, в нем, вероятно, в результате временного разрушения и возобновления пруда, был потерян клонально передающийся женский геном (табл. 4).
«Чистые» популяционные системы съедобной лягушки могут быть примером стабильного типа воспроизводства, сложившегося после вытеснения гибридами родительского вида. Как показывают примеры западноевропейских популяционных систем, стабильным вариантом может быть сочетание ди- и триплоидных гибридов. Частотно-зависимый отбор может устанавливать оптимальное соотношение таких форм в общих популяционных системах. Согласно концепции сетчатой эволюции Л.Я. Боркина и И.И. Даревского (1980), следующим этапом эволюции «чистых» популяционных систем может быть их переход к партеногенетическому размножению.
Необходимо отметить, что диплоидные гибриды, производящие одновременно два типа гамет (см. табл. 3), могут быть единственной формой гибридов, способных образовывать популяции, состоящие из однотипных особей. Впрочем, в таких популяциях (при условии равенства продукции гамет Lи R) половину потомства составят гибридолизные особи, которые должны гибнуть на том или ином этапе онтогенеза до достижения половозрелости.
Как ясно из приведенных рассуждений, эволюция той или иной популяционной системы лягушек зависит от свойств входящих в ее состав гибридов. Многие исследователи указывают, что клональная передача генома должна способствовать накоплению в нем мутаций. Для описания этого процесса привлекается, в частности, модель, называемая «храповиком Мюллера» (Plötner, 2005; Хедрик, 2003; храповик — шестерня с загнутыми зубцами, которая может крутиться только в одну сторону). В соответствии с этой аналогией можно ожидать, что в клональном геноме будет происходить лишь накопление мутантных аллелей без их удаления в ходе рекомбинации. Именно вращение такого «храповика» приводит к нежизнеспособности особей, выщепляющихся в результате гибридолиза. Гибридные лягушки, возникающие от скрещивания разных клональных линий, получают различные клональные геномы, для которых маловероятно совпадение летальных мутаций. То, что такие особи зачастую оказываются жизнеспособными (Vorburger, 2001), подтверждает предположение, что изменения клонального генома оказываются по преимуществу разрушительными. Однако, с нашей точки зрения, следует рассмотреть и другие предположение.
У мерогемиклональных гибридов, в отличие от подавляющего большинства других организмов, геномы выполняют разные функции. Клональный геном обеспечивает свою передачу из поколения в поколение и элиминацию неклональных геномов. Неклональный геном выполняет обычную роль, если не учитывать того, что он функционирует совместно с клональным и оказывается исключен из передачи следующим поколениям. Изменения (как адаптивные с точки зрения выполняемых функций, так и стохастические) в неклональном геноме теряются, а в клональном — накапливаются и определяют его специфическую эволюцию. Как было отмечено выше, клональные линии R. esculenta воспроизводятся намного эффективнее, чем гибриды, получающиеся от скрещивания родительских форм (Plötner, 2005). Вероятно, это обстоятельство является следствием адаптации клонального генома к мерогемиклональному воспроизводству. Конкуренция между различными гибридными линиями может приводить к увеличению доли клональных геномов, более успешно обеспечивающих свое воспроизводство. Такая клональная геномная эволюция принадлежит к теоретически новой, практически не изученной категории эволюционных изменений.
Важнейший вывод из приведенных рассуждений заключается в том, что разнообразие типов популяционных систем лягушек отражает постоянно идущий в них эволюционный поиск. Для изучения этих феноменов необходимо сочетание полевых и цитогенетических исследований, а также использование математического моделирования.
Перспективы дальнейшего изучения комплекса Rana esculenta в Харьковской области
Не вызывает сомнения, что центр разнообразия зеленых лягушек, обнаруженный в Харьковской области, нуждается в дальнейшем изучении. Среди важнейших задач, которые требуют своего решения, следует назвать:
— определение границ распространения различных форм лягушек и типов их популяционных систем в бассейне Северского Донца;
— описание экологических, этологических, морфологических и других отличий между особями, принадлежащим к разным цитогенетическим формам (отличающихся по количеству, составу и происхождению их геномов и составу продуцируемых ими гамет);
— регистрация состава популяционных систем в разных типах естественных местообитаний и установление системы их воспроизводства (подбора пар родительских особей, состава их гамет и состава их потомства);
— выяснение особенностей мейоза и гаметогенеза у триплоидных особей;
— установление цитологических механизмов, позволяющих некоторым гибридным особям лягушек одновременно продуцировать гаметы обоих родительских видов;
— разработка математических моделей популяционного воспроизводства у зеленых лягушек и проверка полученных на основании этих моделей прогнозов на естественных популяционных системах.
Для решения сложных цитогенетических проблем перспективным представляется кариоанализ в давленых препаратах мацерированных тканей по модифицированной методике В.В. Клименко (Klymenko, 2001). Вероятно, с помощью этого метода можно будет определять кариотип яйцеклеток, регистрировать диплоидные сперматоциты, а также изучать топографию клеток с различными кариотипами в тканях лягушек, что практически невозможно сделать другими способами.
Авторы выражают искреннюю признательность коллегам по изучению зеленых лягушек в Харьковской области: Л.Я. Боркину, М.В. Владимировой, Г.А. Ладе, С.Н. Литвинчуку, В.В. Манило, С.Ю. Морозову-Леонову, Ю.М. Розанову, Т.С. Фоменко и А.В. Шабановой, а также благодарят за ценные консультации профессоров Л.А. Атраментову, Г.Н. Жолткевича и В.В. Клименко.
Литература
Ананьева Н.Б., Боркин Л.Я., Даревский И.С., Орлов Н.Л. Земноводные и пресмыкающиеся. Энциклопедия природы России. — М.: ABF, 1998. — 576 с.
Боркин Л.Я., Виноградов А.Е., Розанов Ю.М., Цауне И.А. Полуклональное наследование в гибридогенном комплексе Rana esculenta: доказательство методом проточной ДНК-цитометрии // Докл. АН СССР. — 1987. — Т. 295. № 5. — С. 1261–1264.
Боркин Л.Я., Даревский И.С. Сетчатое (гибридогенное) видообразование у позвоночных // Ж. общ. биол. — 1980. — Т. 41, № 4. — С. 485–506.
Боркин Л.Я., Зиненко А.И., Коршунов А.В., Лада Г.А., Литвинчук С.Н., Розанов Ю.М., Шабанов Д.А. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины // Матеріали Першої конференції Українського Герпетологічного Товариства — К.: Зоомузей ННПМ НАНУ, 2005. — С. 23–26.
Боркин Л.Я., Литвинчук С.Н., Розанов Ю.М., Скоринов Д.В. О криптических видах (на примере амфибий). — Зоологический журнал. — 2004. — Т. 83, № 8. — С. 936–960.
Ведмедеря В.И. Некоторые данные о лягушках рода Rana в Харьковской области (по материалам Музея природы ХГУ) // Вестн. Харьк. ун-та, Харьков. — 1984. — Т. 262. — С. 99–101.
Коршунов А.В. Существуют ли в Харьковской области чистые популяционные системы Rana esculenta ? // Биоразнообразие и роль зооценоза в естественных и антропогенных экосистемах. — Днепропетровск: Изд-во ДНУ, 2005. — С. 363–365.
Коршунов О.В., Бабініч Т.В., Зіненко О.І., Шабанов Д.А. Різноманіття зелених жаб (Rana esculenta complex) в Харківській області: морфологічний аспект вивчення // Біологія та валеологія. — Вип. 6. — Харків: ХДПУ, 2004. — С. 24–30.
Лада Г.А. О необходимости сохранения уникальных «чистых» популяций диплоидной съедобной лягушки (Rana esculenta Linnaeus, 1758) в Белгородской и Харьковской областях // Проблемы охраны и рационального использования природных экосистем и биологических ресурсов. — Пенза, 1998. — С. 333–335.
Манило В.В. Миксоплоидия у Rana ridibunda ridibunda и Rana esculenta (ANURA, RANIDAE) из Житомирской области Украины // Матеріали Першої конференції Українського Герпетологічного Товариства — К.: Зоомузей ННПМ НАНУ, 2005. — С. 99–104.
Межжерин С.В., Морозов-Леонов С.Ю., Некрасова О.Д., Куртяк Ф.Ф., Жалай Е.И. Пространственная структура гибридогенного комплекса зеленых лягушек Rana esculenta (ANURA, RANIDAE) на территории Украины // Матеріали Першої конференції Українського Герпетологічного Товариства — К.: Зоомузей ННПМ НАНУ, 2005. — С. 110–144.
Сурядная Н.Н. Материалы по кариологии зеленых лягушек (Rana ridibunda Pallas, 1771; Rana lessonae Camerano, 1882; Rana esculenta Linnaeus, 1758) с территории Украины // Вестник зоологии. – 2003. – Т. 37, № 1. – С. 33-40.
Сурядна Н.М. Зелені жаби фауни України: морфологічна мінливість, каріологія та особливості біології: Автореферат дис… к-та біолог. наук: 03.00.08 / НАН України, Ін-т зоології ім. І. І.Шмальгаузена. — Київ, 2005.
Таращук С.В. К методике определения европейских зеленых лягушек группы Rana esculenta (Amphibia, Ranidae) // Вестн. зоол. — 1985. — № 3. — С. 83–85.
Хедрик Ф. Генетика популяций. — М.: Техносфера, 2003. — 592 с.
Чернай А. Фауна Харьковской губернии и прилежащих к ней мест. — Харьков, 1852 — 50 с.
Abt G., Reyer H.-U. Mate choice and fitness in a hybrid frog: Rana esculenta females prefer Rana lessonae males over their own//Behav Ecol Sociobiol. —1993. — Vol. 32. — P. 221-228.
Berger L.Is Rana esculenta lessonae Camerano a distinct species? // Ann. Zool. PAN. — 1964. — Vol. 22, № 13. — P. 245-261.
Borkin L.J., Korshunov A.V., Lada G.A., Litvinchuk S.N., Rosanov J.M., Shabanov D.A., Zinenko A.I. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology. — 2004. — Vol. 11, № 3. — P. 194–213.
Borkin L.J., Lada G.A., Litvinchuk S.N., Melnikov D.A., Rosanov J.M. The first record of mass triploidy in hybridogenic green frog Rana esculenta in Russia (Rostov oblast’) // Russian Journal of Herpetology. — 2006. — in press.
Broska J. Vocal response of male European water frogs (Rana esculenta complex) to mating and territorial calls // Behav. Process. — 1982. — V.7, №1. — P. 649-659.
Christiansen D.G., Fog K., Pedersen B.V., Boomsma J.J. Reproduction and hybrid load in all-hybrid populations of Rana esculenta water frogs in Denmark // Evolution. — 2005. — Vol. 59, № 6. — P. 1348–1361.
Dubois A., Günther R. Klepton and Synklepton: two new evolutionary systematics categories in zoology // Zool. Jb. Syst.—1982. — Bd. 109. — P. 290–305.
Günther R. Die europaische Wasserfrosch-Gruppe - ein evolutionsbiologischer sonderfall // Biol. Rdsch. 1979. — Bd. 17. № 4. — S 217–228.
Günther R. Untersuchungen der Meiose bei Mannchen von Rana ridibunda Pall., Ranna lessonae Cam. und der Bastardform "Rana esculenta" L. (Anura) // Biologisches Zentralblatt. — 1975. — Bd. 94, N. 3. — S. 277–294.
Hellriegel B., Reyer H.-U.Factors influencing the composition of mixed populations of a hemiclonal hybrid and its sexual host//Journal of Evolutionary Biology. —2000. — Vol. 13— P. 906-918.
Holenweg Peter A.K. Survival in adults of the water frog Rana lessonae and its hybridogenetic associate Rana esculenta // Canadian Journal of Zoology. — 2001. — Vol. 79. — P. 652-661.
Holenweg Peter A.-K., Reyer H.-U., Abt Tietje G. Species and sex ratio differences in mixed populations of hybridogenetic water frogs: The influnce of pond features // Ecoscience. — 2002 — Vol. 9 — P. 1-11.
Klymenko V.V. Parthenogenesis and cloning in the silk worm Bombyx mori: problems and prospects // J. Insect Biotechnol. And Sericol. — 2001. — Vol. 70. — P. 155–165.
Krynicki I.A. Observationes quaedam de reptilibus indigenis, Bull. Soc. Imp. Nat. Moscou. — 1837. — Vol. 10, № 3. — P. 46–69.
Miura I., Ohtani H., Nakamura M., Saitoh K. Fluorescence replication banding of frog chromosomes//Cellular and Molecular Life Sciences. —1997. — Vol. 53. — P. 73-77.
Negovetic L., Anholt B.R., Semlitsch R.D., Reyer H.-U. Specific responses of sexual and hybridogenetic European waterfrogs tadpoles to temperatures // Ecology. — 2001. — Vol. 82. — P. 766-774.
Plenet S., Hervant F., Joly P. Ecology of the hybridogenetic Rana esculenta complex - differential oxygen requirements of tadpoles // Evolutionary Ecology. — 2000. — Vol. 14. — P. 13-23.
Plötner J. Die westpaläarktichen Wasserfrösche. Bielefeld: Laurenti-Verlag, 2005. — 161 S.
Reyer H.-U., Walti M.-O., Battig A., Altwegg R., Hellriegel B. Low proportions of reproducing hemiclonal females increase the stability of a sexual parasite-host system (Rana esculenta, R. lessonae) // Journal of Animal Ecology. — 2004. — Vol. 73. — P. 1089-1101.
Schultz R.L. Hybridization, unisexuality, and polyploidy in teleost Poeciliopsis (Poeciliidae) and other vertebrates // Amer. Natur. — 1969. — Vol. 103. — P. 605–619.
Som C., Anholt B.R., Reyer H.-U. The effect of assortative mating on the coexistence of a hybridogenetic waterfrog and its sexual host // American Nauralist. — 2000. — Vol. 156. — P. 34-46.
Tunner H.G. Die Klonale Struktur einer Wasserfroschpopulation // Z. zool. Syst. und Evolut.-forsch.—1974. — Bd. 12, № 4. — P. 309–314.
Uzzell T.M., Berger L. Electrophoretic phenotypes of Rana ridibunda, Rana lessonae and their hybridogenic associate Rana esculenta // Proc. Acad. nat. Sci. Phila. — 1975. — Vol. 127. — P. 13-24.
Vorburger C. Fixation of deleterious mutations in clonal lineages: evidence from hybridogenetic frogs // Evolution. —2001. — Vol. 55. — P. 2319-2332.
Vorburger C., Reyer H.-U. A genetic mechanism of species replacement in European waterfrogs? // Conservation Genetics. — 2003 — Vol. 4. — P. 141-155.