Усова Е. Е., Кравченко М. А., Шабанов Д. А. Внутрипопуляционные онтогенетические стратегии у зеленых лягушек (Pelophylax esculentus complex) // Вісник Харківського національного університету імені В. Н. Каразіна. Серія: біологія. – 2015. – Вип. 25. – С. 223-238.
УДК: (57.022+57.032):591.84:591.158.1:597.851
Внутрипопуляционные онтогенетические стратегии у зеленых лягушек (Pelophylax esculentus complex)
Е.Е.Усова, М.А.Кравченко, Д.А.Шабанов
Харьковский национальный университет имени В.Н.Каразина (Харьков, Украина)
e.e.usova@gmail.com, marinakravchenko2106@gmail.com, d.a.shabanov@gmail.com
С использованием скелетохронологии исследовали 575 представителей Pelophylax esculentus complex (193 P. ridibundus, 348 диплоидных и 34 триплоидных P. esculentus) из Харьковской области. Разнообразие лягушек по возрасту и длине тела описали с помощью аппроксимации по методу наименьших квадратов двумя линиями регрессии, соответствующими относительно более мелким и более крупным одновозрастным особям. Эти две группы особей сравнили по популяционнобиологическим параметрам и зарегистрировали синдромы (комплексы связанных признаков), позволяющие рассматривать их как проявления внутрипопуляционных онтогенетических стратегий (ВОС) малоразмерности и крупноразмерности. Малоразмерность/крупноразмерность характеризуются низкой/высокой скоростью роста, ранним/поздним началом участия в нересте самок и поздним/ранним - самцов, высокой/низкой продолжительностью жизни, низкой/высокой плодовитостью самок и большим/малым количеством сезонов размножения, в которых принимает участие особь. С использованием имитационного моделирования показали, что отличия P. ridibundus и P. esculentus по ВОС могут обеспечивать их устойчивое сосуществование в гемиклональной популяционной системе.
Ключевые слова: Pelophylax esculentus complex, скелетохронология, стратегия, малоразмерность, крупноразмерность, имитационное моделирование, гемиклональные популяционные системы.
Внутрішньопопуляційні онтогенетичні стратегії у зелених жаб (Pelophylax esculentus complex)
О.Є.Усова, М.О.Кравченко, Д.А.Шабанов
З використанням скелетохронології дослідили 575 представників Pelophylax esculentus complex (193 P. ridibundus, 348 диплоїдних і 34 триплоїдних P. esculentus) з Харківської області. Різноманітність жаб за віком і довжиною тіла описали за допомогою апроксимації за методом найменших квадратів двома лініями регресії, що відповідають відносно дрібнішим та більшим особинам одного віку. Дрібніших та більших особин порівняли за популяційнобіологічними параметрами і зареєстрували синдроми (комплекси пов'язаних ознак), що дозволяють розглядати їх як прояви внутрішньопопуляційних онтогенетичних стратегій (ВОС) малорозмірності та крупнорозмірності. ВОС малорозмірності/крупнорозмірності характеризуються низькою/високою швидкістю росту, раннім/пізнім початком участі у нересті самок і пізнім/раннім – самців, високою/низькою тривалістю життя, низькою/високою плодючістю самок і великою/низькою кількістю сезонів розмноження, в яких бере участь особина. Із застосуванням імітаційного моделювання показали, що відмінності P. ridibundus і P. esculentus за ВОС можуть забезпечувати їх стійке співіснування в геміклональній популяційної системі.
Ключові слова: Pelophylax esculentus complex, скелетохронологія, стратегія, малорозмірність, крупнорозмірність, імітаційне моделювання, геміклональні популяційні системи.
The water frogs' (Pelophylax esculentus complex) intrapopulation ontogenetic strategies
O.E.Usova, M.O.Kravchenko, D.A.Shabanov
575 representatives of Pelophylax esculentus complex (193 P. ridibundus, 348 diploid and 34 triploid P. esculentus) from Kharkivska oblast have been studied using skeletochronology. With approximation by the LS method, frogs’ diversity on age and body length has been characterized by two regression lines, which correspond to relatively smaller and bigger individuals of the same age. We compared these individuals by population-biological parameters and registered syndromes (complexes of related traits), allowing their consideration as manifeatation of intrapopulation ontogenetic strategies (IOS). IOS of undersized/oversized is characterized by relatively low/high growth rate, high/low life expectancy, low/high fertility of females and relatively large/small number of breeding seasons in which the individual takes part. Participation in spawning begins early/late in undersized/oversized females and vice versa in males. By using simulation it has been shown that differences in IOS between P. ridibundus and P. esculentus can provide their sustainable coexistence in the hemiclonal population system.
Key words: Pelophylax esculentus complex, skeletochronology, strategy, undersized, oversized, simulation, hemiclonal population systems.
Введение
Гибридогенный комплекс зеленых лягушек, Pelophylax esculentus complex (=Rana esculenta complex) состоит из двух родительских видов: прудовой лягушки, Pelophylax lessonae (Camerano, 1882), и озерной лягушки, Pelophylax ridibundus (Pallas, 1771), а также их межвидовых гибридов различной плоидности (Plötner, 2005; Шабанов, Литвинчук, 2010). Для этих гибридов принято использовать имя, аналогичное видовому: съедобная лягушка, Pelophylax esculentus (Linnaeus, 1758). Для P. esculentus характерно гемиклональное наследование, при котором производимые ими гаметы несут или геном P. ridibundus, или геном P. lessonae, или и тот, и другой геном. Для P. esculentus характерно совместное обитание с представителями родительских видов в гемиклональных популяционных системах, ГПС (Кравченко, 2013; Шабанов, 2015). Известны также ГПС, состоящие исключительно из P. esculentus.
Один из способов обозначения типов ГПС связан с перечислением входящих в их состав форм лягушек, при этом литера L обозначает P. lessonae, R – P. ridibundus, а E – P. esculentus. Наличие в составе ГПС полиплоидов P. esculentus обозначается как Ep. В бассейне р. Северский Донец на территории Харьковской области описан Северско-Донецкий центр разнообразия Pelophylax esculentus complex, характеризующийся необычным составом ГПС (Borkin et al., 2004; Боркин и др., 2005; Mezhzherin et al., 2010; Шабанов, 2015). Для него характерны R‑E‑ГПС и R‑E‑Ep‑ГПС, а также популяции P. ridibundus, а половозрелые представители P. lessonae полностью отсутствуют.
Pанее (Шабанов и др., 2014) была высказана гипотеза, что разнообразие особей внутри одной популяции или ГПС (в случае зеленых лягушек) может быть описано как проявление внутрипопуляционных онтогенетических стратегий, ВОС. Онтогенетическая стратегия – это иерархия приоритетов, проявляющихся в развитии организма. Внутрипопуляционная стратегия – один из дискретных или объединенных в континуум вариантов реализации видоспецифичной стратегии, встречающихся у представителей одной популяции или ГПС. ВОС характеризуется определенным синдромом (комплексом связанных признаков), отражающим специфику адаптации к среде.
Целью данной работы была выяснение того, отражает ли внутрипопуляционное разнообразие представителей Pelophylax esculentus complex из Харьковской области различие их ВОС. Если это предположение верно, на одних и тех же этапах онтогенеза у одних особей приоритет имеет повышение их собственной жизнеспособности, а у других – размножение. Для достижения цели работы решались следующие задачи:
– определить формы, к которым относятся лягушки, измерить их длину, установить, с помощью скелетохронологии, их возраст и ретроспективно определить их динамику роста на протяжении последних лет жизни;
– изучить эмпирическое разнообразие изученных особей по размеру их тела в определенном возрасте, как по ведущему признаку (т.е. признаку, который может отражать ВОС);
– сравнить группы лягушек, отличающихся по ведущему признаку, по характеристикам, отражающим приоритеты в их онтогенезе: скорости роста, продолжительности жизни, плодовитости самок, времени начала участия в размножении;
– в случае значимых отличий между группами, выделенными по ведущему признаку, по связанным с онтогенетическими приоритетами характеристикам, описать особенности ВОС, реализуемых представителями этих групп;
– установить возможное значение разнообразия особей по их ВОС для устойчивости ГПС представителей Pelophylax esculentus complex.
Материал и методы
Состав изученной нами совокупности зеленых лягушек охарактеризован в табл. 1. Все лягушки были собраны в Харьковской области, 550 – в Северско-Донецком центре разнообразия Pelophylax esculentus complex (486 – в R‑E‑Ep‑ГПС и 64 – в R‑E‑ГПС), а 25 – в бассейне Днепра (7 – в R‑E‑ГПС и 18 – в L‑E‑R‑ГПС. Сбор лягушек проводился в время нереста в темное время суток.
Этапы обработки материала подробно описаны в публикации, касающейся части изученного в данной работе материала (Усова, 2014). Принадлежность особи к одной из форм Pelophylax esculentus complex определяли по внешним признакам и размеру эритроцитов (Бондарева и др., 2012). Состав геномов ряда изученных нами особей подтвердили с помощью проточной ДНК-цитометрии (Borkin et al., 2004). Проточную цитометрию проводили С.Н.Литвинчук и Ю.М.Розанов в ЦИН РАН (г. Санкт-Петербург, РФ).
Таблица 1.
Состав изученной совокупности зеленых лягушек с точки зрения их формы (принадлежности к родительскому виду, диплоидным или триплоидным гибридам) и пола
|
Форма |
Незрелых, экз. |
Самок, экз. |
Самцов, экз. |
Всего, экз. |
|
P. ridibundus |
13 |
149 |
31 |
193 |
|
P. esculentus, 2n |
2 |
134 |
212 |
348 |
|
P. esculentus, 3n |
2 |
12 |
20 |
34 |
|
Всего |
17 |
295 |
263 |
575 |
У лягушек удаляли 2-й (длиннейший) палец задней ноги. Его 4-ю фалангу очищали, декальцинировали и получали срезы средней части диафиза (толщиной 20–22 мкм) на замораживающем микротоме. Срезы окрашивали гематоксилином Эрлиха, помещали в глицерин под покровным стеклом и фотографировали под микроскопом USB-камерой. На электронных фотографиях в программе Adobe Photoshop CS5 подсчитывали и измеряли линии склеивания. Результаты измерений переводили в микрометры, используя установленный при помощи фотографирования объект-микрометра коэффициент пересчета. Результаты измерений собирали и обрабатывали в базе данных, созданной в программе Statistica 8, StatSoft Inc.
При интерпретации скелетохронологических препаратов необходимо принять два решения. Во-первых, необходимо определить, какие линии склеивания образовались во время зимовок, а какие связаны с остановкой роста в теплый сезон. Во-вторых, следует принять предположение, сколько линий склеивания было резорбировано (разрушено) из-за расширения костномозговой полости, расположенной внутри кости. После определенного возраста расширение костномозговой полости прекращается, и она заполняется эндостальной костной тканью со своими линиями склеивания. Эти линии важно не спутать с линиями склеивания в периостальной части кости (Смирина, 1983).
Точно определить, сколько линий склеивания в периостальной части кости было резорбировано при расширении костномозговой полости, в ряде случаев невозможно. Однако обоснованное предположение о количестве таких линий можно принять, сравнивая препараты, полученные от разных индивидов из одного или близких местообитаний. Основаниями для принятия гипотезы о количестве резорбированных линий являются поперечник границы между эндостальной и периостальной частью кости (чем он больше, тем больше линий склеивания в периостальной кости могло быть резорбировано); размер первой полностью сохранившейся линии склеивания (которая по своим размерам может соответствовать первой, второй или последующим линиям склеивания, образующимся у лягушек в исследуемом местообитании); видимые на препарате остатки частично резорбированных линий. Принятые нами на этих основаниях предположения о количестве резорбированных линий у изученной совокупности лягушек отражены на рис. 1.
Вычисление длины тела лягушки во время прошлых зимовок основано на предположении, что P/L=li/Lj, где P – усредненный наружный поперечник кости (ее периостальной части) в момент исследования, L – длина тела в момент исследования, li – усредненный поперечник линии склеивания, образовавшейся в момент определенной зимовки, Lj – длина тела, которую особь имела во время соответствующей зимовки. В таком случае Lj=L×li/P (Усова, Шабанов, 2009). На основании оценок размера тела особи во время двух последовательных зимовок можно установить относительный годовой прирост длины тела gi=(Lj-Lj-1)/Lj-1.
Поиск оптимальных значений коэффициентов для каждой из моделей, аппроксимирующих зарегистрированное нами эмпирическое разнообразие лягушек, осуществляли с помощью надстройки «Поиск решения» в Microsoft Excel. Для этого вычисляли сумму квадратов отклонений между наблюдаемой и расчетной (в соответствии с каждой из моделей) длиной тела каждой особи. Затем надстройка «Поиск решения» минимизировала сумму квадратов отклонений, изменяя значения коэффициентов, подставляемых в соответствующие используемым моделям уравнения.
Плодовитость самок определяли, получая кладки икры в искусственных условиях. Кладки фотографировали цифровым фотоаппаратом, а потом производили подсчет икринок по фотографии.
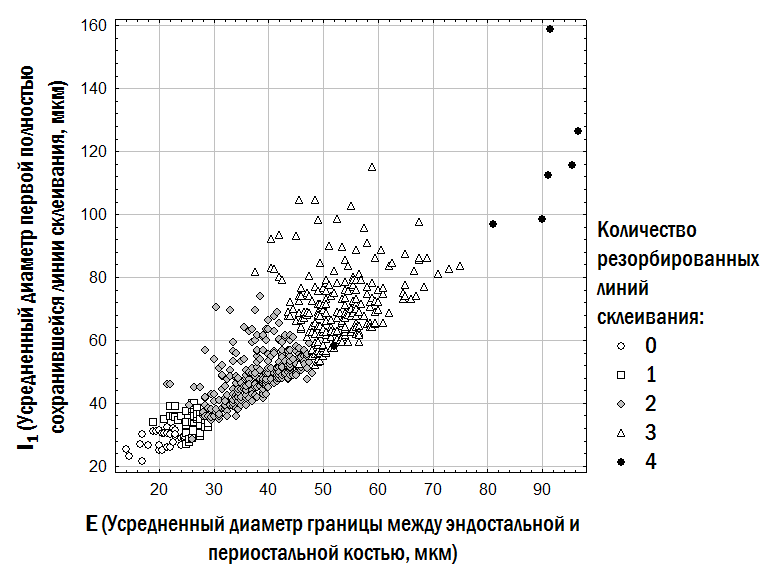
Рис. 1. Принятые в данной работе предположения о количестве резорбированных линий склеивания в периостальной кости у изученной совокупности лягушек
Изучение устойчивости ГПС Pelophylax esculentus complex проводили с использованием имитационной модели, разработанной в соответствии с поставленным нами заданием А.О.Леоновым при участии М.В.Владимировой, Г.Н.Жолткевича (ХНУ имени В.Н.Каразина), J.Newman и Q.Mair (Glasgow Caledonian University). Эта модель является консольным приложением для Java, которое имитирует изменения состава ГПС зеленых лягушек. На основании заданных начальных параметров модель пошагово проводит перестройку описываемой модельной ГПС (рис. 2).
К числу начальных параметров, которые задаются при моделировании, относятся:
– начальный состав (численность особей разных форм и возрастов) модельной ГПС;
– популяционно-биологические параметры всех рассматриваемых в ней групп лягушек (отличающихся по генотипам и возрасту), включающие ее жизнеспособность (вероятность выживания в неконкурентной среде), конкурентоспособность (вероятность сохранения в ходе конкурентного исключения), вероятность образования пары с партнером в ходе размножения, возраст первого размножения, максимальную продолжительность жизни, плодовитость и потребность в ресурсах;
– варианты всех возможных скрещиваний с указанием вероятностей появления различных генотипов в потомстве;
– емкость среды (количество доступных в ней ресурсов);
– сценарии иммиграции и эмиграции (при необходимости).
На каждом шаге работы модели судьба каждой особи в моделируемой ГПС (ее выживание и размножение) определяется случайными процессами и описывается вероятностями, заданными в начальных параметрах модели. С каждыми начальными условиями выполнялось по 10 имитаций. Исходы имитаций классифицировали в зависимости от состава форм лягушек, присутствующих в модельной ГПС через 500 циклов (соответствующих 500 годам), после чего определяли распределение вероятностей разных исходов имитаций в зависимости от начального состава ГПС и принятых параметров жизнеспособности.
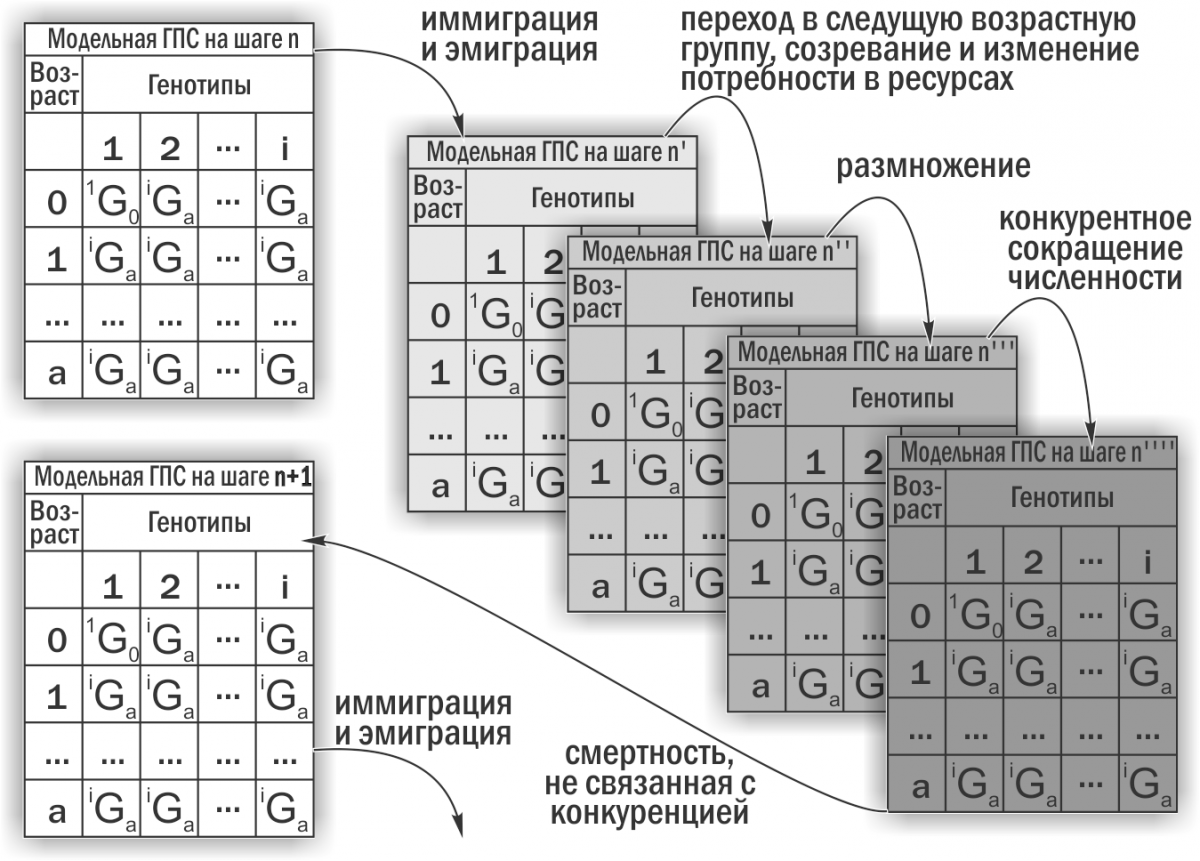
Рис. 2. Вычисления в каждом цикле работы имитационной модели (Шабанов, 2015)
Результаты и обсуждение
1. Описание зависимости размера лягушек от их возраста. Зависимость размера изученных лягушек от их возраста показана на рис. 3. Отраженное на этом рисунке разнообразие особей может быть описано с помощью разных моделей (табл. 2).
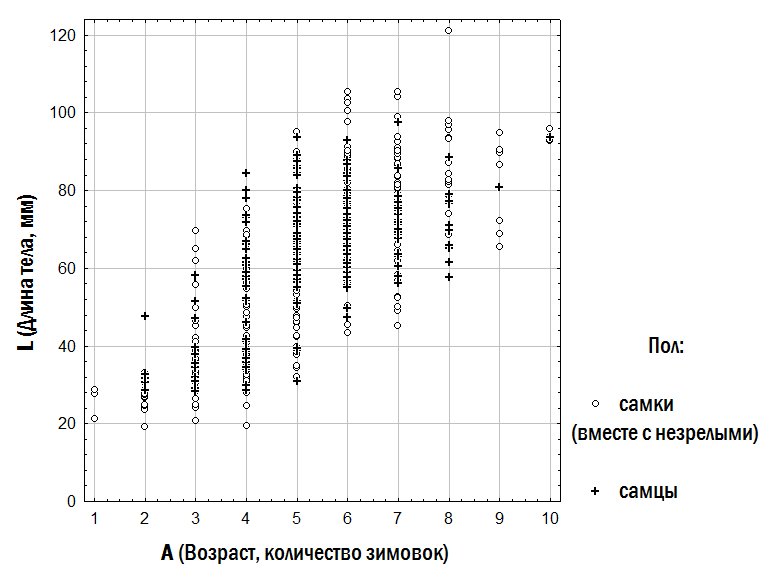
Рис. 3. Зависимость размера лягушек от их возраста
Таблица 2.
Результаты аппроксимации наблюдаемых соответствий возраста (A) и длины тела (L) зеленых лягушек с помощью трех различных моделей
|
Модели для описания эмпирических данных |
Аппроксимация |
Сумма квадратов отклонений |
|
|
Линейная |
L = a + b×A |
L = 16,6 + 8,6×A |
105 087,5 |
|
Квадратичная |
L = a + b×A + c×A2 |
L = –3,7 + 17,2×A – 0,8×A2 |
99 351,1 |
|
Две линейные зависимости для относительно меньших и относительно бóльших одновозрастных особей |
lL = la + lb×A; bL = ba + bb×A |
lL = 9,7 + 8,3×A; bL = 32,5 + 8,3×A |
34 906,5 |
|
Три линейные зависимости для относительно меньших, промежуточных и бóльших особей |
lL = la + lb×A; mL = ma + mb×A; bL = ba + bb×A |
lL = 3,7 + 8,4×A; mL = 14,6 + 9,1×A; bL = 32,9 + 9,1×A |
17 559,7 |
Третья из использованных нами моделей предусматривает, что разнообразие особей одного возраста по их размеру может быть описано с помощью двух линий регрессии, одна из которых соответствует относительно меньшим, а другая – относительно бóльшим одновозрастным особям. Обозначения, использованные в табл. 2, связаны с английскими словами lesser – меньший и bigger – больший. При аппроксимации наблюдаемого разнообразия возраст-размерных соответствий лягушек использованный нами алгоритм вычислял значения коэффициентов двух линейных зависимостей, минимизируя сумму квадратов расстояний от каждой точки до ближайшей к ней прямой. Мы задали ограничения, в соответствии с которыми ba ≥ la и bb ≥ lb (обозначения см. в табл. 2). Результат описания эмпирических данных с помощью двух линий регрессии, соответствующих относительно меньшим и относительно бóльшим одновозрастным особям, показан на рис. 4. Модель с тремя линиями регрессии (табл. 2) построена аналогичным образом; в ней добавляется линия регрессии, соответствующая промежуточным (medium) по относительному размеру особям.
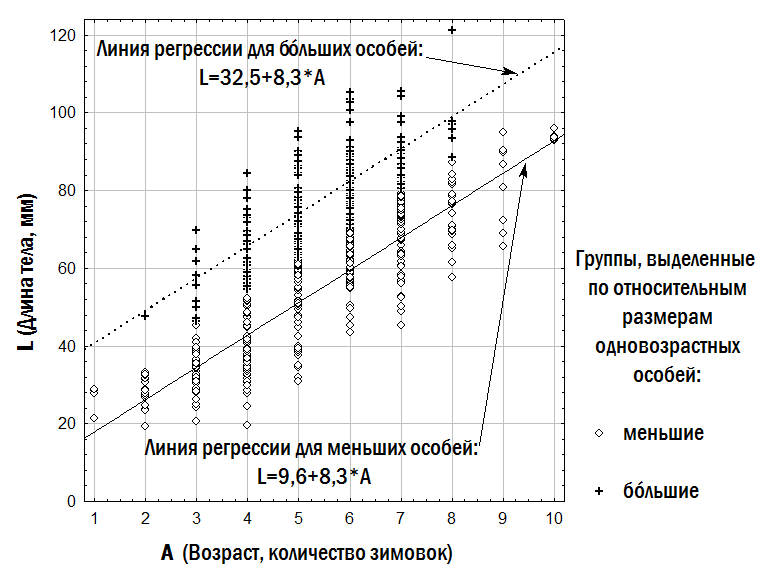
Рис. 4. Аппроксимация эмпирического возраст-размерного разнообразия зеленых лягушек с помощью двух линейных зависимостей
Количество моделей, с помощью которых можно описывать наблюдаемое разнообразие, потенциально неограниченно. Например, модель с тремя линиями регрессии дает лучший результат, чем с двумя. Для выбора оптимального уровня детальности описания можно использовать критерий каменистой осыпи Кэттеля (Халафян, 2007). Согласно этому критерию, для описания эмпирических данных следует использовать модель такого уровня детальности, которому соответствует наиболее острый перегиб зависимости суммы квадратов необъясненных ошибок от количества принимаемых в модели предположений. Этому критерию (рис. 5) соответствует модель с двумя линиями регрессии (рис. 4), которая и используется далее в данной работе.
2. Сравнение разных форм лягушек по ростовому показателю (G). Все рассмотренные нами формы лягушек (диплоиды и триплоиды P. esculentus, а также P. ridibundus) обоих полов присутствуют как среди относительно меньших, так и среди относительно бóльших особей. Чтобы сравнить относительные размеры особей разного возраста, необходима независимая от возраста мера. Такой мерой может быть предложенная нами величина G, названная ростовым показателем. Ростовой показатель вычисляли по формуле Gx(j)=2×(Lx-lLj)/(bLj-lLj)-1, где Gx(j) – ростовой показатель особи x возраста j; Lx – длина тела особи x, lLj – ожидаемое значение длины особи возраста j, соответствующее линии регрессии для относительно меньших особей (рис. 4), а bLj – ожидаемое значение длины особи возраста j, соответствующее линии регрессии относительно бóльших особей. Для лягушек, размер которых соответствует уравнению регрессии для относительно меньших особей, ростовой показатель принимает значение ‑1, для тех, которые лежат на линии регрессии для бóльших особей, G=1, а для тех, кто находится посредине между двумя линиями регрессии, – G=0.
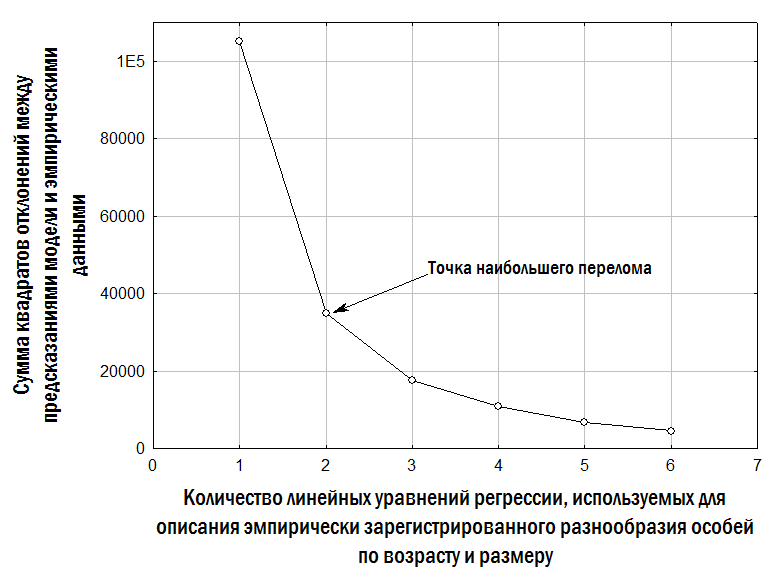
Рис. 5. Выбор сложности модели для описания эмпирически зарегистрированного разнообразия особей, с помощью критерия каменистой осыпи Кэттеля
С помощью критерия Краскела–Уоллиса (непараметрического аналога дисперсионного анализа) мы определили влияние на ростовой показатель (G) лягушек трех факторов: формы, пола и локалитетов, где собирали выборки. Мы сравнивали три локалитета с самыми многочисленными выборками: окрестности биостанции ХНУ имени В.Н.Каразина в с. Гайдары (132 особи), Иськов пруд в окрестностях с. Гайдары (137 особей) и Нижний Добрицкий пруд в окрестностях с. Великая Гомольша (130 особей). Все эти три местообитания расположены в Змиевском районе Харьковской области в Национальном природном парке «Гомольшанские леса» или в его окрестностях. Единственный фактор, влияние которого на ростовой показатель (G) оказалось значимым, – форма представителей Pelophylax esculentus complex. Влияние пола оказалось незначимым и даже меньшим, чем влияние локалитета (рис. 6).
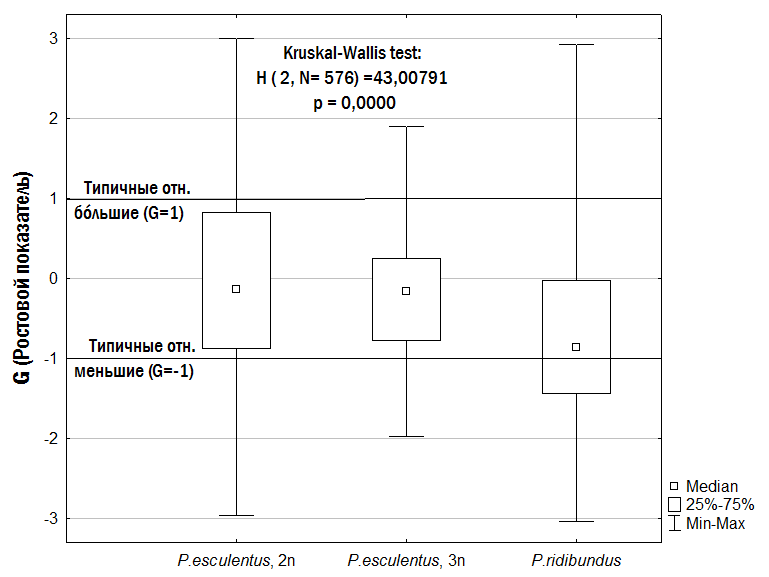
Рис. 6. Сравнение значений ростового показателя (G) у трех рассмотренных форм зеленых лягушек
Несмотря на то, что среди относительно меньших (G<0) и относительно бóльших (G>0) особей есть представители всех трех форм, представители родительского вида имеют более низкие значения ростового показателя (G), чем гибриды. Для всей изученной нами совокупности особей при попарном сравнении форм по Манну-Уитни ростовой показатель (G) значимо ниже у P. ridibundus, чем у диплоидов P. esculentus (p<0,000001), и чем у триплоидов P. esculentus (p=0,0028). Уровень отличий таков, что его следует считать значимым даже с учетом поправки на множественные сравнения. Отличия между диплоидами и триплоидами P. esculentus незначимы (p=0,59).
3. Связь относительных размеров одновозрастных особей со скоростью их роста. Показанные на рис. 4 линии регрессии описывают не динамику индивидуального роста каждой особи, а разнообразие особей, наблюдаемых в выборках из ГПС. Примеры динамики индивидуального роста показаны на рис. 7. На этом рисунке показаны все особи, для которых рассчитаны размеры для 7 последовательных зимовок (№№ 5–13 на рис. 7). Все эти особи (а также все, для которых получены оценки размеров для 6 зимовок) относятся к группе относительно меньших (G<0). Среди относительно бóльших особей (G>0) есть только 4, для которых рассчитаны размеры во время 5 зимовок (№№1–4 на рис. 7). Таким образом, на рис. 7 показаны особи обеих размерных групп, рост которых прослежен на протяжении наибольшего срока. Среди таких особей есть как P. ridibundus, так и P. esculentus; большинство этих особей – самки, но среди них есть и один самец (особь №8).
Разница между относительно меньшими (G<0) и относительно бóльшими (G>0) одновозрастными особями связана с разной скоростью их роста в возрасте 3–4 года (табл. 3). Переход из группы меньших особей в группу бóльших (примером которого может быть рост особи №2 на рис. 7) связан с периодом их ускоренного роста.
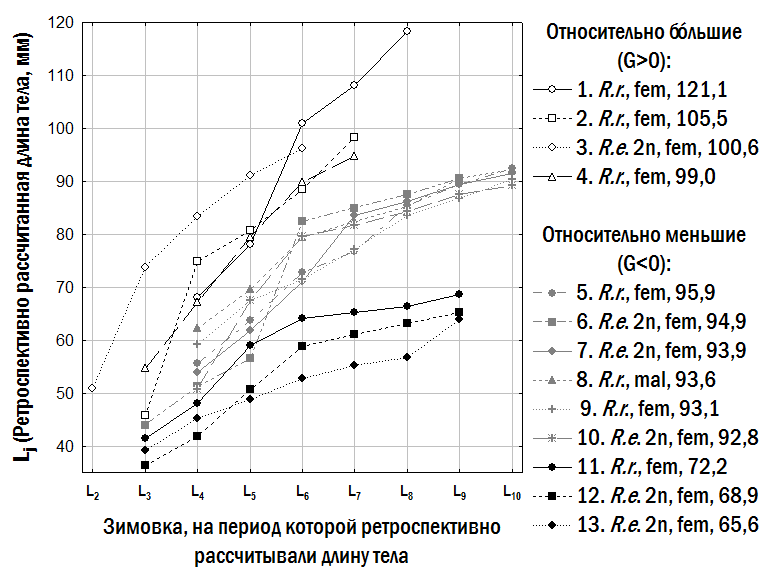
Рис. 7. Примеры динамики изменения длины тела отдельными представителями Pelophylax esculentus complex. Для каждой особи указана ее форма, пол и длина тела (в мм)
Таблица 3.
Сравнение относительных приростов на 3-м и 4-м годах жизни относительно меньших (G<0) и относительно бóльших (G>0) одновозрастных особей
|
Показатель |
Относительно меньшие особи (G<0) |
Относительно бóльшие особи (G>0) |
p (по Манну-Уитни) |
|
Медианное значение прироста на 3-м году |
lgme3 = 0,170 |
bgme3 = 0,450 |
0,001 |
|
Медианное значение прироста на 4-м году |
lgme4 = 0,176 |
bgme4 = 0,288 |
0,000007 |
4. Связь относительных размеров одновозрастных особей с плодовитостью самок. Чтобы проверить предположение (Шабанов и др., 2014), что относительно меньшие и бóльшие особи отличаются по их внутрипопуляционным онтогенетическим стратегиям (ВОС), надо сравнить эти группы особей по плодовитости самок, продолжительности жизни и сроку начала размножения.
Ростовой показатель (G) значимо связан с плодовитостью самок: коэффициент непараметрической корреляции Спирмана rS равен 0,34 (p=0,009). Уравнение регрессии, описывающее связь плодовитости (F) с возрастом (A), имеет для относительно меньших самок вид F = ‑1325 + 335×A, а для относительно бóльших – F = 179 + 316×A. Сравнивая плодовитость самок из двух размерных групп (рис. 8) для тех возрастов, по которым мы можем провести сравнение (5, 6 и 7 лет), мы можем убедиться, что и средние значения, и медианы откладываемого количества икринок у относительно бóльших самок в 2-3 раза выше, чем у относительно меньших самок того же возраста.
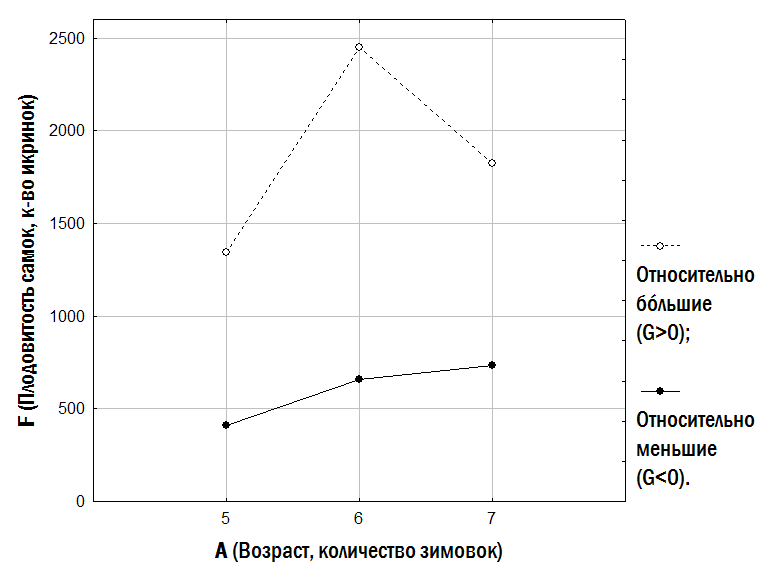
Рис. 8. Медианы количества икринок, откладываемых самками двух размерных групп
5. Связь относительных размеров одновозрастных особей с их продолжительностью жизни. Самые старые особи в изученной нами выборке относятся к группе меньших по размеру (рис. 4, рис. 7). Среди особей старше 6 лет возраст относительно меньших значимо (p=0,045 при сравнении по Манну-Уитни) превышает возраст относительно бóльших. Помимо этого, с ожидаемой продолжительностью жизни тесно связан прирост на 4-м году жизни (рис. 9).
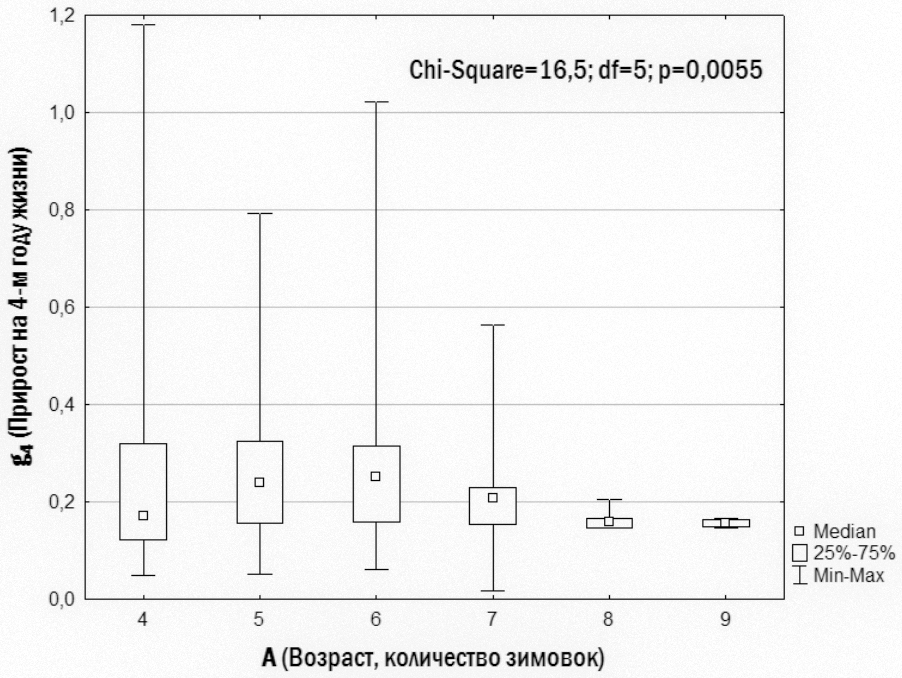
Рис. 9. Приросты 4-го года жизни, характерные для особей, доживших до разного возраста
На основании результатов, отраженных на рис. 9, можно установить, что у особей в возрасте от 4-х до 6-ти лет приросты 4-го года могут быть весьма различными: и небольшими (до 30%), и весьма значительными (до 100%). Большинство особей 7-ми лет и все особи более старших возрастов имеют небольшие приросты 4-го года. Это означает, что лягушки, которые быстро растут между третьей и четвертой зимовкой, имеют незначительные шансы оказаться «долгожителями».
6. Связь относительных размеров одновозрастных особей с наступлением половозрелости и началом участия в размножении. Для оценки времени созревания изучаемых нами лягушек могут быть использованы три группы данных.
Первый способ оценки времени наступления половой зрелости связан с изучением динамики роста особей в течение их жизни. До того, как особь начинает участвовать в нересте, вся полученная ею энергия, не затраченная на текущие потребности жизнедеятельности, тратится на ее рост и развитие. После того, как особь приступает к размножению, существенная часть затрат энергии (особенно у самок) оказывается связанной с размножением, и ее рост должен замедляться. Определив момент замедления роста в течение жизни особи, мы можем предположить, что он соответствует возрасту, в котором она начинает «инвестировать» не только в себя, но и в своих потомков. Подобный анализ можно выполнить по данным, представленным на рис. 10.
Снижение показанной на рис. 10 величины (отношения прироста в последующем году к приросту в предшествующем) означает замедление скорости роста в определенном возрасте, а значения, превышающие 1, – ускорение роста. На основании показанных на рис. 10 данных можно установить, что среди относительно меньших особей (G<0) замедление роста у самок происходит на 4-м году жизни, а у самцов – на 5-м и 6-м; среди относительно бóльших особей (G>0) у самок – на 6-м году жизни, а у самцов – на 5-м (снижение на 8-м году связано с тем, что показанное на рисунке значение вычислено по единственному самцу из этой группы, пережившему 8-ю зимовку).
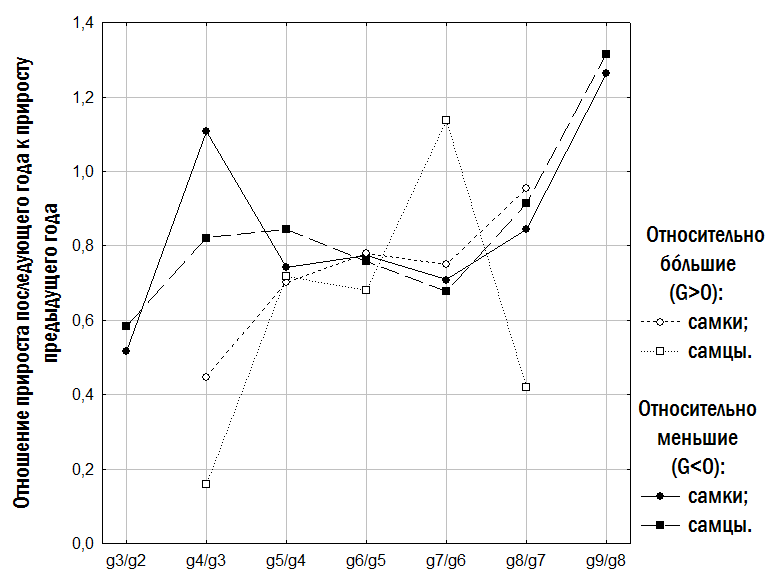
Рис. 10. Изменения средних отношений годовых приростов для двух последовательных годов жизни у самок и самцов групп, выделенных по относительным размерам одновозрастных особей
Второй способ оценки начала участия особей в размножении связан с анализом их возрастного состава в собранных на нересте выборках. Исследованных нами лягушек собирали на местах нереста во время нереста; единственное исключение было сделано для незрелых особей. Если лягушки младших возрастных классов представлены в наших сборах меньше, чем представители старших, это означает, что во время нереста они находятся не на нерестилищах, а где-то в другом месте. С этой точки зрения, возрасту, в котором все особи впервые принимают участие в нересте, должен соответствовать максимум на показанных на рис. 11 гистограммах. Сокращение следующих по численности возрастных классов связано, в первую очередь, с их смертностью.
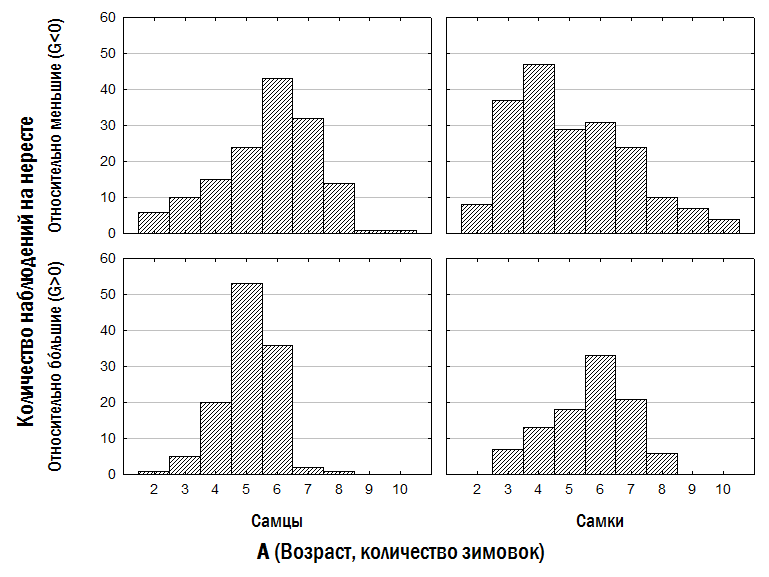
Рис. 11. Распределение лягушек в изученной нами выборке по полу и принадлежности к размерной группе
Бóльшие самцы приходят на нерест в полном составе раньше, чем меньшие: в 5 лет, а не в 6 (рис. 11). Бóльшие самки приходят позже, чем меньшие: в 6 лет, а не в 4. Эти результаты полностью соответствуют тем, которые были получены с использованием предыдущего метода оценки начала участия в размножении (рис. 10).
Наконец, третий способ определения срока созревания особей – определение наличия или отсутствия сформированных гамет у особей определенного возраста. Благодаря кариогенетическим исследованиям, которые проводила О.В.Бирюк, мы смогли установить наличие или отсутствие сформированных сперматозоидов в семенниках 37 молодых самцов всех трех рассматриваемых в данной работе форм (P. ridibundus, диплоидов и триплоидов P. esculentus). Среди 25 двухлеток сперматозоиды не производил 21, среди 9 трехлеток – 5. Все три изученных четырехлетних самца производили сперматозоиды. Среди двух- и трехлеток те, которые производили сперматозоиды, были мельче (незначимо), чем те, что еще не приступили к их продукции (Biriuk et al., 2015).
Резюмируя сказанное, мы можем установить, что, по всей видимости, физиологическое достижение половозрелости у всех лягушек в изученном нами регионе происходит к возрасту 3–4 года. Тем не менее, к полноценному участию в нересте (сопровождающемся присутствием на местах нереста и существенными затратами энергии на размножение) большинство лягушек переходит существенно позже.
7. Относительно бóльшие или меньшие размеры одновозрастных особей как проявления внутрипопуляционных онтогенетических стратегий (ВОС). Мы установили, что лягушки двух выделенных нами групп отличаются по синдрому (связанному комплексу) признаков, отражающему специфику их адаптации к среде. Эти отличия соответствуют ранее изложенным представлениям о внутрипопуляционных онтогенетических стратегиях (Шабанов и др., 2014). Согласно данным в указанной статье определениям, тугорослость – ВОС, которая характеризуется относительно низкой скоростью роста, замедленным созреванием, уменьшенным количеством потомков за каждый цикл размножения и относительно большей продолжительностью жизни. В свою очередь, скороспелость – ВОС, которая характеризуется относительно высокой скоростью роста, ранним созреванием, повышенным количеством потомков за каждый цикл размножения и относительно более короткой продолжительностью жизни.
Изложенные в упомянутой статье предположения подтвердились во всех отношениях, кроме одного. В ней предполагалось, что связанная с меньшими размерами стратегия характеризуется замедленным созреванием, а связанная с бóльшими размерами – с ускоренным созреванием. Это предположение основывалось в первую очередь на результатах исследования Bufo bufo (L., 1758), серых жаб (Маро и др., 2008). Как мы убедились, в отношении представителей Pelophylax esculentus complex это предположение не оправдывается; при переходе от одной стратегии к другой сроки созревания разных полов изменяются в разном направлении. В связи с этим, в табл. 4, обобщающей отличия зарегистрированных нами ВОС, рассматриваемые нами стратегии названы малоразмерностью и крупноразмерностью. Эти стратегии в целом соответствуют ранее описанным тугорослости и скороспелости (Шабанов и др., 2014), отличаясь от них более сложным характером вступления в размножение.
Таблица 4.
Сравнение внутрипопуляционных онтогенетических стратегий (ВОС) малоразмерности и крупноразмерности
|
ВОС
Характеристики |
Малоразмерность (undersized); G<0 |
Крупноразмерность (oversized) ; G>0 |
||
|
Самки |
Самцы |
Самки |
Самцы |
|
|
Размеры |
Относительно меньшие: lL = 9,7 + 8,3×A |
Относительно бóльшие: bL = 32,5 + 8,3×A |
||
|
Скорость роста |
Относительно низкая: lgme3 = 0,170; lgme4 =0,176 |
Относительно высокая: bgme3 =0,450; bgme4 =0,288 |
||
|
Полноценное участие в нересте |
Относит. раннее: с 4 лет |
Относит. позднее: с 6 лет |
Относит. позднее: с 6 лет |
Относит. раннее: С 5 лет |
|
Продолжительность жизни |
Относительно высокая: до 10 лет |
Относительно низкая: до 8 лет |
||
|
Плодовитость самок |
Относит. низкая: |
‒ |
Относит. высокая: |
‒ |
|
Количество сезонов размножения |
Особо большое: |
Относ. большое: |
Особо малое: |
Относит. малое: |
Следует особо подчеркнуть, что малоразмерные (G<0) и крупноразмерные (G>0) особи встречаются в одних и тех же местообитаниях и принадлежат как к P. ridibundus, так и к диплоидам и триплоидам P. esculentus. Тем не менее, как установлено нами (рис. 6), у P. esculentus ВОС крупноразмерности встречается значимо чаще, чем у P. ridibundus.
8. Возможное значение разнообразия ВОС, реализуемых представителями Pelophylax esculentus complex в одной ГПС. Поскольку ВОС малоразмерности и крупноразмерности встречаются у зеленых лягушек, совместно обитающих в одной ГПС, мы рассматриваем это разнообразие как приспособление к внутрипопуляционным взаимодействиям. Для изучения его значения может быть полезно имитационное моделирование. Описанный ниже феномен был впервые зарегистрирован нами в ходе экспериментов с имитационной моделью ГПС зеленых лягушек, выполненной в среде Microsoft Excel (Кравченко, 2013). Затем он был детальнее исследован нами с использованием описанной выше модели А.О.Леонова (Shabanov et al., 2015).
Мы рассматривали R-E-ГПС, состоящие из P. ridibundus и диплоидных P. esculentus, которые передают в гаметах женский геном P. lessonae. Для обозначения геномов P. ridibundus мы используем литеру R, а для обозначения геномов P. lessonae – литеру L. Мужские геномы обозначаются надстрочным знаком Y, а женские – X (гетерогаметным полом у зеленых лягушек является мужской; Plötner, 2005). Клональный характер передачи генома обозначается заключением его символа в скобки. Используя эти обозначения, можно перечислить все типы скрещиваний, которые могут происходить в описываемой ГПС:
– воспроизводство родительского вида: ♀XRXR×♂XRYR → ♀♀XRXR : ♂♂XRYR;
– скрещивание представителей родительского вида с гибридами, при котором все потомство состоит из гибридов: ♀XRXR×♂YR(XL) → ♀♀XR(XL); ♀XR(XL)×♂XRYR → ♀♀XR(XL) : ♂♂YR(XL);
– скрещивание гибридов с образованием нежизнеспособных представителей отсутствующего в данной ГПС родительского вида: ♀XR(XL)×♂YR(XL) → ♀♀(XL)(XL) → † (Plötner, 2005; Шабанов, 2015).
Трансформации описанной R-E-ГПС могут привести к одному из трех исходов:
исход №1: переход к популяции P. ridibundus; вследствие исчезновения P. esculentus;
исход №2: сохранение R-E-ГПС вследствие сосуществования P. ridibundus и P. esculentus;
исход №3: гибель ГПС вследствие исчезновения в ней P. ridibundus.
Если популяционнобиологические параметры P. ridibundus и P. esculentus, принимаемые при моделировании, идентичны, доля P. esculentus в ГПС непрерывно растет, вследствие их более эффективного воспроизводства. В таком случае трансформации R-E-ГПС приводят к исходу №3. Такой результат означает, что представление об идентичности параметров жизнеспособности у P. ridibundus и P. esculentus не соответствует действительности. Если бы это предположение было справедливо, R-E-ГПС существовали бы относительно недолго и закономерно исчезали. В то же время нам известно, что такие системы широко распространены в бассейнах р. Мжа и р. Уды, правых притоков р. Северского Донца (Шабанов, 2015).
Мы предположили, что устойчивое сосуществование P. ridibundus и P. esculentus может объясняться более низкой жизнеспособностью P. esculentus. В соответствии с этим предположением, снижение жизнеспособности гибридов должно компенсировать преимущество в их воспроизводстве. Как показано на рис. 12, это предположение не оправдалось: в зависимости от соотношений смертности P. ridibundus и P. esculentus реализуется или исход №1, или исход №3.
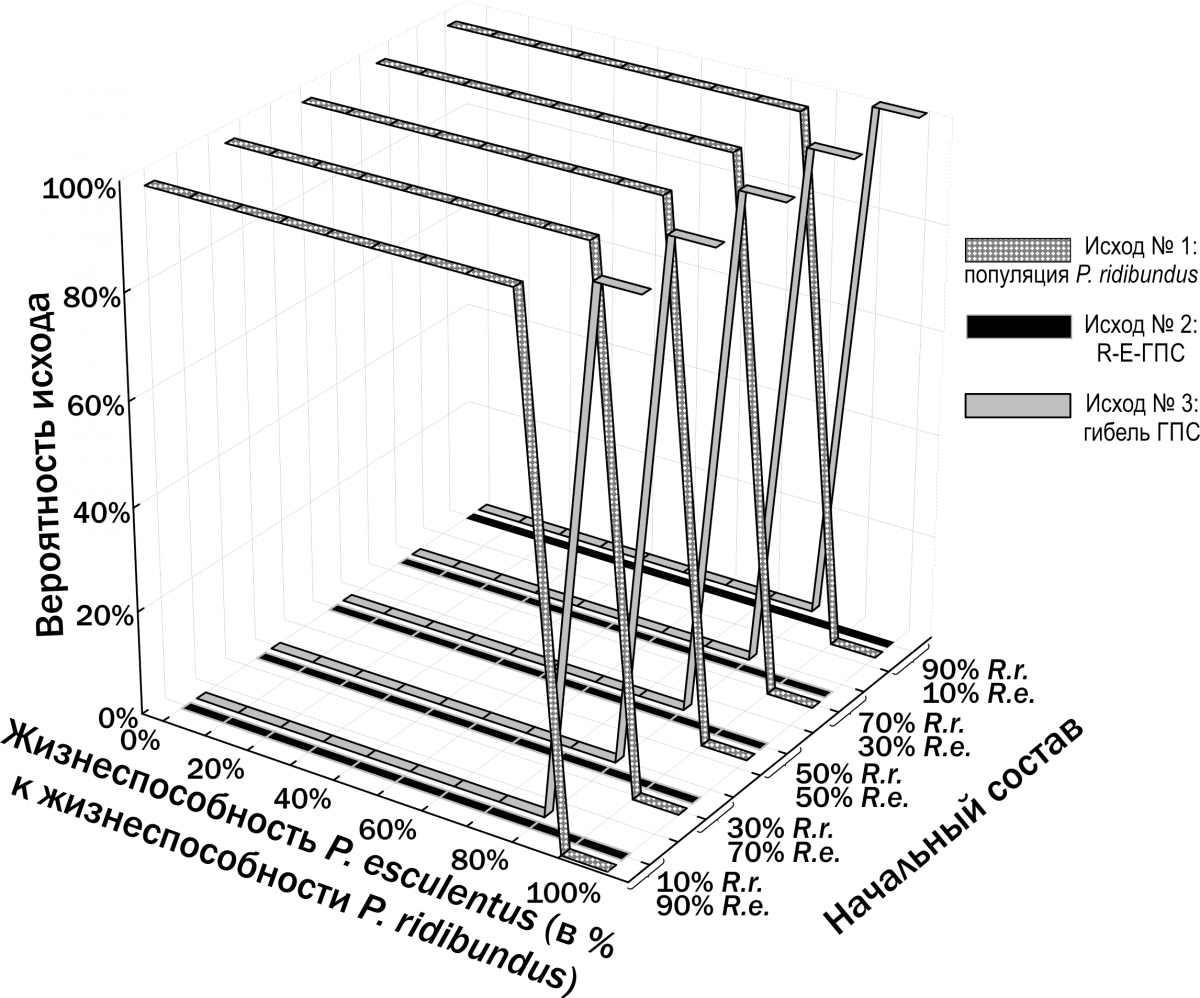
Рис. 12. Результаты моделирования трансформаций R-E-ГПС при предположении, что P. ridibundus и P. esculentus отличаются только по вероятности их гибели
Иной результат наблюдается в случае, если P. ridibundus и P. esculentus реализуют различные ВОС. На рис. 13 показаны результаты моделирования в том случае, если P. ridibundus реализует ВОС тугорослости, а P. esculentus – ВОС скороспелости (Шабанов и др., 2014). В этом случае возникает зона соотношения жизнеспособностей двух форм, где возможно их устойчивое сосуществование.
Мы предполагаем, что устойчивое существование R-E-ГПС связано с разнообразием внутрипопуляционных онтогенетических стратегий особей в их составе.
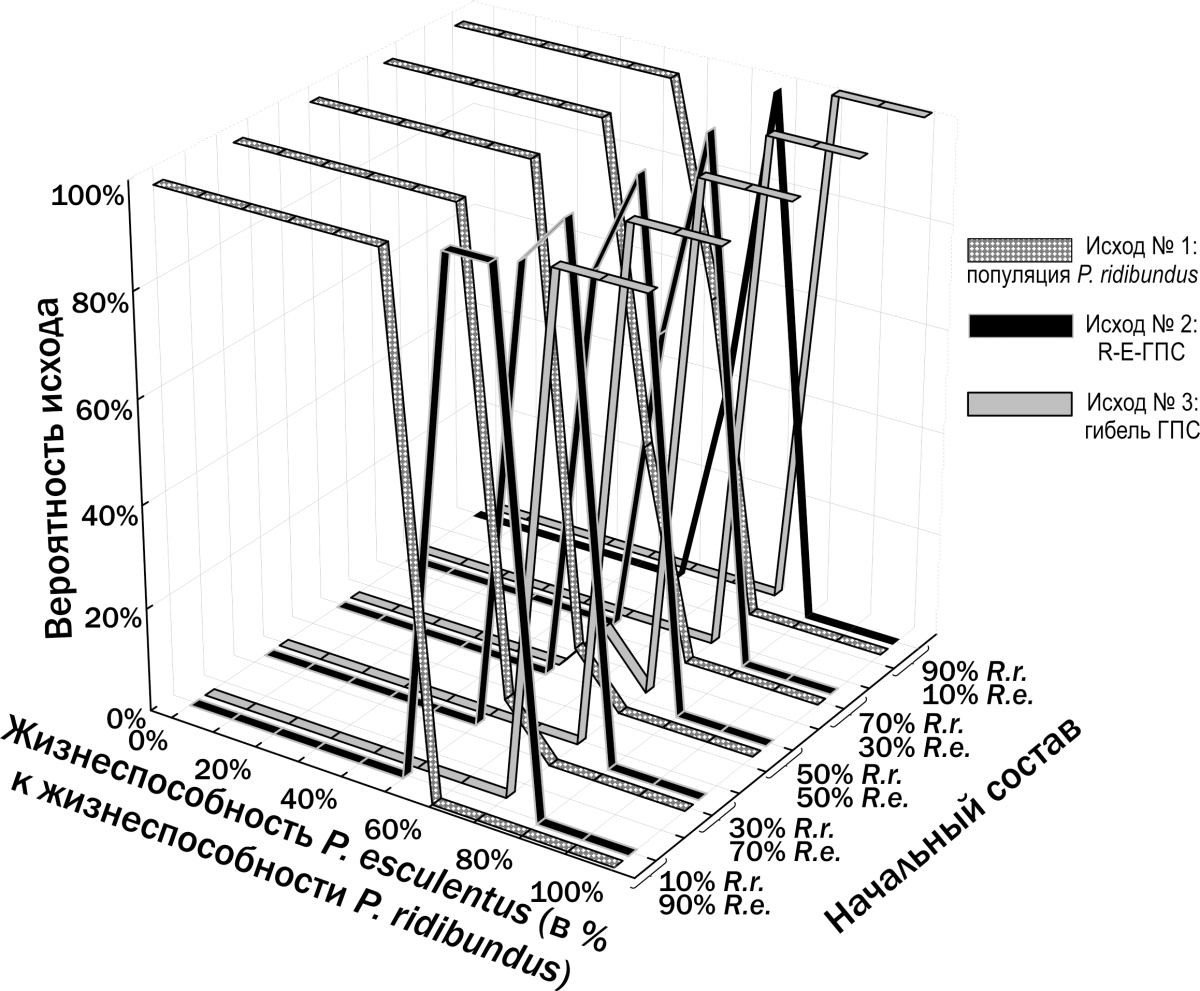
Рис. 13. Результаты моделирования трансформаций R-E-ГПС при предположении, что P. ridibundus и P. esculentus отличаются по их ВОС
Благодарности
Авторы выражают глубокую благодарность классику скелетохронологических исследований амфибий Э.М.Смириной (г. Москва) за помощь в освоении использованной в работе методики, О.В.Бирюк за кариогенетические исследования ряда изученных нами особей лягушек, членам группы популяционной экологии амфибий Харьковского национального университета имени В.Н.Каразина А.В.Коршунову, Е.В.Мелешко и другим за помощь в сборе и обработке материала, сотрудникам ЦИН РАН (г. Санкт-Петербург) С.Н.Литвинчуку и Ю.М.Розанову за определение генотипов ряда изученных особей с использованием проточной ДНК-цитометрии, а также А.О.Леонову, М.В.Владимировой, Г.Н.Жолткевичу (ХНУ имени В.Н.Каразина), J.Newman и Q.Mair (Glasgow Caledonian University) за помощь в имитационном моделировании. Кроме того, авторы выражают благодарность А.А.Атемасову за конструктивную критику данной работы.
Список литературы
Бондарева А.А., Бибик Ю.С., Самило С.М., Шабанов Д.А. Цитогенетические особенности эритроцитов зеленых лягушек из Северско-Донецкого центра разнообразия Pelophylax esculentus complex // Вісник Харківського національного університету імені В.Н.Каразіна. Серія «Біологія». – 2012. – Вип.15 (№1008). – С. 116–123. /Bondareva A.A., Bibik Yu.S., Samilo S.M., Shabanov D.A. Tsitogeneticheskiye osobennosti eritrotsitov zelenykh lyagushek iz Seversko-Donetskogo tsentra raznoobraziya Pelophylax esculentus complex // Vіsnyk Kharkіvs'kogo natsional'nogo unіversitetu іmenі V.N.Karazіna. Serіya «Bіologіya». – 2012. – Vyp.15 (№1008). – S. 116–123.
Боркин Л.Я., Зиненко А.И., Коршунов А.В. и др. Массовая полиплоидия в гибридогенном комплексе Rana esculenta (Ranidae, Anura, Amphibia) на Востоке Украины // Матеріали I конференції Українського Герпетологічного Товариства – К.: Зоомузей ННПМ НАНУ, 2005. – С. 23–26. /Borkin L.Ya., Zinenko A.I., Korshunov A.V. i dr. Massovaya poliploidiya v gibridogennom komplekse Rana esculenta (Ranidae, Anura, Amphibia) na Vostoke Ukrainyi // MaterIali I konferentsiyi Ukrayinskogo Gerpetologichnogo Tovaristva – K.: Zoomuzey NNPM NANU, 2005. – S. 23–26.
Кравченко М.О. Екологічна стійкість популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex). Автореф. дис. … канд. біол. наук / 03.00.16 – екологія. – Дніпропетровськ, 2013. – 20с. /Kravchenko M.O. Ekologіchna stіykіst' populyatsіynykh system gіbrydogennogo kompleksu zelenykh zhab (Pelophylax esculentus complex). Аvtoref. dys. … kand. bіol. nauk / 03.00.16 – ekologіya. – Dnіpropetrovs'k, 2013. – 20s.
Маро А.Н., Шабанова А.В., Шабанов Д.А. Могут ли условия развития головастиков Bufo bufo определять темпы постметаморфического роста и созревания жаб? // Вопросы герпетологии. Материалы III съезда Герпетологического общества им. А.М.Никольского. – СПб, 2008. – С. 274–280. /Maro A.N., Shabanova A.V., Shabanov D.A. Mogut li usloviya razvitiya golovastikov Bufo bufo opredelyat' tempy postmetamorficheskogo rosta i sozrevaniya zhab? // Voprosy gerpetologii. Materialy III s'yezda Gerpetologicheskogo obshchestva im. A.M.Nikol'skogo. – SPb, 2008. – S. 274–280.
Смирина Э.М. Прижизненное определение возраста и ретроспективная оценка размеров тела серой жабы (Bufo bufo) // Зоол. журн. – 1983. – Т.63, №3. – С. 437–444. /Smirina E.M. Prizhiznennoye opredeleniye vozrasta i retrospektivnaya otsenka razmerov tela seroy zhaby (Bufo bufo) // Zool. zhurn. – 1983. – T.63, №3. – S. 437–444./
Усова Е.Е. Возраст и скорость роста зеленых лягушек (Pelophylax esculentus complex) Нижнего Добрицкого пруда (Змиевской район Харьковской области) // Вісник Харківського національного університету імені В.Н. Каразіна. Серія: біологія. – 2014. – Вип.20, №1100. – С. 204–212. /Usova Ye.Ye. Vozrast i skorost' rosta zelenykh lyagushek (Pelophylax esculentus complex) Nizhnego Dobritskogo pruda (Zmiyevskoy rayon Khar'kovskoy oblasti) // Vіsnyk Kharkіvs'kogo natsional'nogo unіversytetu іmenі V.N. Karazіna. Serіya: bіologіya. – 2014. – Vyp.20, №1100. – S. 204–212.
Усова Е.Е., Шабанов Д.А. Об оптимизации методики ретроспективной оценки динамики размеров тела представителей Pelophylax esculentus complex (Amphibia, Ranidae) при помощи скелетохронологии // Zoocenosis-2009. Біорізноманіття та роль тварин в екосистемах. – Дніпропетровськ: Ліра, 2009. – С. 278–280. /Usova Ye.Ye., Shabanov D.A. Ob optimizatsii metodiki retrospektivnoy otsenki dinamiki razmerov tela predstaviteley Pelophylax esculentus complex (Amphibia, Ranidae) pri pomoshchi skeletokhronologii // Zoocenosis-2009. Bіorіznomanіttya ta rol' tvaryn v ekosistemakh. – Dnіpropetrovs'k: Lira, 2009. – S. 278–280.
Халафян А.А. Statistica 6. Статистический анализ данных. – М.: ООО «Бином-Пресс», 2007. – 512с. /Khalafyan A.A. Statistica 6. Statisticheskiy analiz dannykh. – M.: OOO «Binom-Press», 2007. – 512s./
Шабанов Д.А. Еволюційна екологія популяційних систем гібридогенного комплексу зелених жаб (Pelophylax esculentus complex) Лівобережного лісостепу України. Aвтореф. дис. … д-ра біол. наук / 03.00.16 – екологія. – Дніпропетровськ, 2015. – 36с. /Shabanov D.A. Evolyutsіyna ekologіya populyatsіynykh system gіbrydogennogo kompleksu zelenykh zhab (Pelophylax esculentus complex) Lіvoberezhnogo lіsostepu Ukrayiny. Avtoref. dys. … d-ra bіol. nauk / 03.00.16 – ekologіya. – Dnіpropetrovs'k, 2015. – 36s.
Шабанов Д.А., Коршунов А.В., Кравченко М.А. и др. Внутрипопуляционные онтогенетические стратегии скороспелости и тугорослости: определение на примере бесхвостых амфибий // Вісник Харківського національного університету імені В.Н.Каразіна. Серія: біологія. – 2014. – Вип.22, №1126. – С. 115–124. /Shabanov D.A., Korshunov A.V., Kravchenko M.A. i dr. Vnutripopulyatsionnyye ontogeneticheskiye strategii skorospelosti i tugoroslosti: opredeleniye na primere beskhvostykh amfibiy // Vіsnyk Kharkіvs'kogo natsional'nogo unіversitetu іmenі V.N. Karazіna. Serіya: Bіologіya. – 2014. – Vyp.22, №1126. – S. 115–124.
Шабанов Д.А., Литвинчук С.Н. Зеленые лягушки: жизнь без правил или особый способ эволюции? // Природа. – 2010. – №3. – С. 29–36. /Shabanov D.A., Litvinchuk S.N. Zelenyye lyagushki: zhizn' bez pravil ili osobyy sposob evolyutsii? // Priroda. – 2010. – №3. – S. 29–36.
Biriuk O., Usova O., Meleshko O., Shabanov D. Composition and characteristic of subadult water frogs sample (Pelophylax esculentus complex) // Book of abstracts of the 3rd International workshop-conference: Research and conservation of European herpetofauna and its environment: Bombina bombina, Emys orbicularis, and Coronella austriaca. – Daugavpils, Latvia, 2015. – P.8.
Borkin L.J., Korshunov A.V., Lada G.A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian Journal of Herpetology. – 2004. – Vol.11, No 3. – P. 194–213.
Plötner J. Die westpaläarktichen Wasserfrösche. – Bielefeld: Laurenti-Verlag, 2005. – 161s.
Mezhzherin S.V., Morozov-Leonov S.Yu., Rostovskaya O.V. et al. The ploidy and genetic structure of hybrid populations of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol.44, No 3. – P. 212–216.
Shabanov D., Usova O., Kravchenko M. et al. Sustainable coexistence of the parental species and hemiclonal interspecific hybrids is provided by the variety of ontogenetic strategies // Herpetological Facts Journal. – 2015. – Vol.2. – P. 35–43.
Представлено: Т.Ю.Маркіна / Presented by: T.Yu.Markina
Рецензент: А.А.Атемасов / Reviewer: A.A.Atemasov
Подано до редакції / Received: 1.10.2015