
БиоСтатистика — 10. Тема 6. Сравнение распределений
Сравнение выборок (и его частный случай — сравнение долей) — распространенная категория задач, решаемых в ходе биологического исследования
6.1. Приклади проблем, що потребують порівняння розподілів
Імовірно, порівняння розподілів — настільки ж часте, як і порівняння значень, завдання біологічних досліджень. У цьому тексті тема порівняння розподілів зачіпалася, щонайменше, тричі.
По-перше, прикладом порівняння розподілів є порівняння вибірок за критерієм Фішера, описане в пункті 4.3 (див. рис. 4.3.1). Це — параметричне порівняння, оскільки розподіли в цьому випадку порівнюються за одним параметром, заснованим на припущенні про те, що обидві вибірки мають нормальний розподіл.
По-друге, процедура порівняння розподілів методом
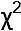
По-третє, ми вже розглянули ситуації, у яких доводиться перевіряти розподіли вибірок на нормальність. Ця перевірка має сенс у багатьох випадках, коли треба вибрати, параметричні чи непараметричні методи слід використовувати. Простий спосіб такої перевірки описаний у пункті 5.2 (див. рис. 5.2.1).
Як зазначалося раніше (див. пункт 1.5), далеко не всі ознаки виражаються числом з неперервного (метричні) або дискретного (меристичні) числового ряду. Багато важливих для біологічного дослідження ознак мають якісний (політомічний або дихотомічний), а також ранговий характер. Щоб порівняти вибірки за такими ознаками, необхідне порівняння розподілів. Наведемо приклади кількох завдань, що потребують використання методів, які розглядаються в цій темі:
— чи відрізняються сусідні локальні популяції за частотами характерних фенів (одиничних якісних зовнішніх ознак);
— чи відрізняються представники різних статей за схильністю до певного захворювання;
— чи справді використання якогось способу лікування приводить до частішого одужання пацієнтів;
— залежно чи ні варіюють у вивченій сукупності об'єктів дві (або більша кількість) якісних ознак.
Почнемо розгляд методів порівняння розподілів з використання критерію
6.2. Визначення зв'язку якісних ознак за допомогою кростабуляції
Звернемося до нашого файлу Pelophylax_example.sta. У ньому розглянуто 57 жаб, що належать до 5 різних генотипів і 2 різних статей. Розподіл за статтю і генотипом вивчених жаб не рівномірний: одні поєднання трапляються частіше, інші — рідше. Чи відображає спостережуваний розподіл зв'язок між статтю і генотипом? Чи можна припустити, що для одних генотипів у нас статистично значимо частіше траплялися самки, а для інших — самці?
Щоб відповісти на це питання, треба визначитися з тим, який розподіл ми б вважали рівномірним. Той, у якому всі 10 (2×5) можливих поєднань статі і генотипу представлені з рівною (або приблизно рівною частотою)? Але, вельми ймовірно, деякі генотипи (наприклад, LLR) трапляються рідше, а деякі (як RR) — частіше. Як нам побудувати дослідження, щоб різна представленість генотипів і різна представленість статей у нашій вибірці не заважала нам отримати відповідь на задане питання?
Для відповіді на це питання відкриємо файл Pelophylax_example.sta і пройдемо шляхом Statistics / Basic Statistics and Tables / Tables and banners.
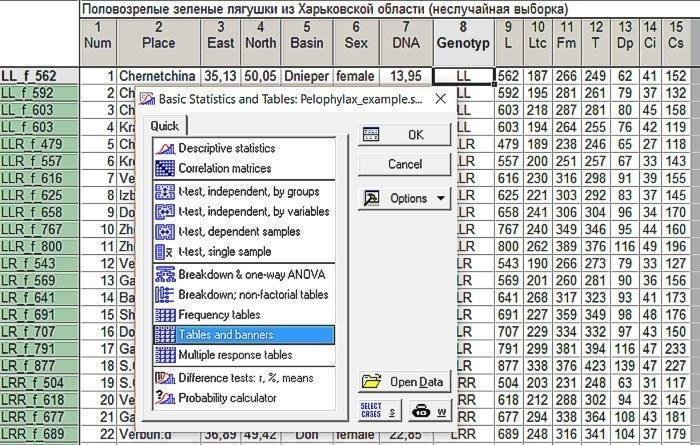
Рис. 6.2.1. Зручний діалог для порівняння розподілів міститься тут
Ми потрапимо на вкладку Crosstabulation Tables. Виберемо там змінні, що відповідають статі і генотипу.
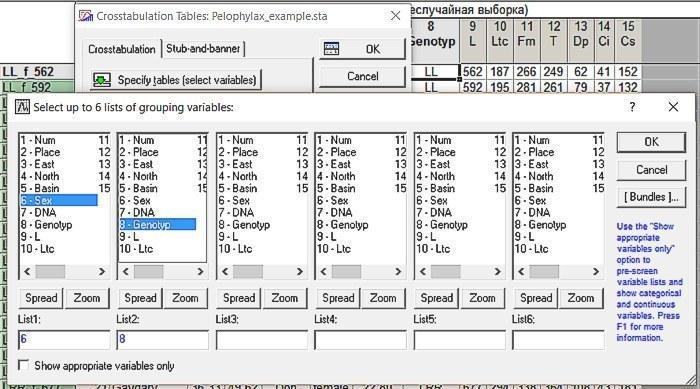
Рис. 6.2.2. Вибір змінних, зв'язок між якими треба проаналізувати
У вікні Crosstabulation Tables Results знімемо галочку Highlight counts, яка дозволяє виділити червоним кольором значення в тих комірках, куди потрапило відносно багато спостережень (за замовчуванням — 10). Поставимо галочки у віконці Expected frequencies, що виводить окреме вікно з очікуваними частотами (при спостережуваних крайових сумах), а також у віконці Pearson & M-L Chi-square, яке викликає порівняння спостережуваних і очікуваних частот за методом
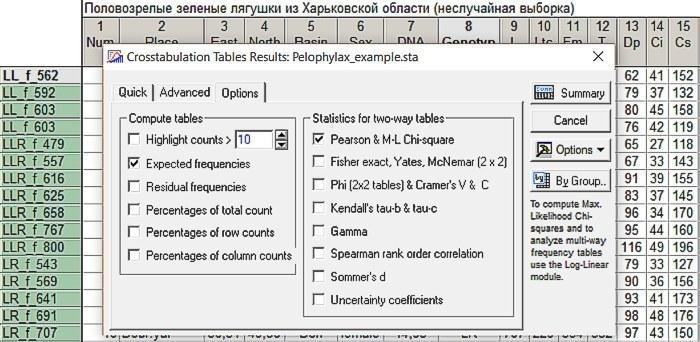
Рис. 6.2.3. Налаштування, потрібні для порівняння очікуваного і спостережуваного розподілів
Натиснувши на кнопку Summary, ми отримаємо одразу дві таблиці.
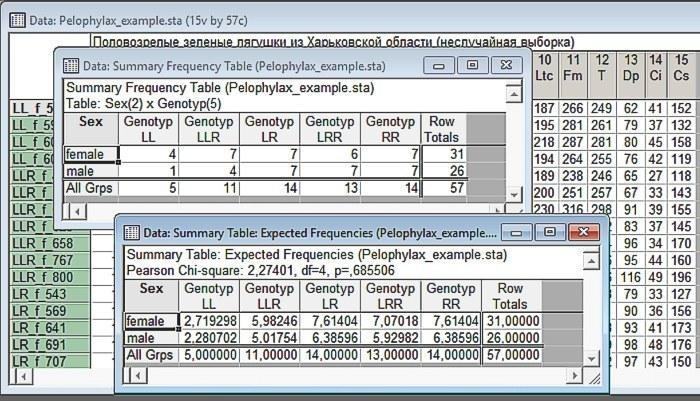
Рис. 6.2.4. Результат аналізу. Вище — спостережуваний розподіл, нижче — очікуваний (за тих самих крайових сум). Переконайтеся, що крайові суми (суми елементів у кожному рядку і кожному стовпці) не змінилися!
Інтерпретація першої з отриманих таблиць дуже проста. Це спостережувані частоти різних поєднань двох вибраних змінних. Тій змінній, яка була вибрана першою, відповідають рядки цієї таблиці, а другій змінній відповідають стовпці. До речі, за допомогою обговорюваного діалогу можна вивчати взаємозв'язок також і трьох або більшої кількості змінних, однак інтерпретація таких багатовходових таблиць виявляється непростою справою (спробуйте і переконайтеся самі!).
У таблиці спостережуваних частот пораховані крайові суми. У нашому випадку це загальна кількість самок і самців, а також особин різних генотипів. Зверніть увагу: жаб з генотипом LL досліджено 5 штук. Загальне співвідношення самок і самців у нашій вибірці 31:26. Якби в особин з генотипом LL спостерігалося те саме співвідношення, ми могли б очікувати в них співвідношення приблизно 2,7:2,8. З іншого боку, загальний розподіл генотипів у нашій вибірці 5:11:14:13:14. Якби особини були розподілені рівномірно, ті самі співвідношення часток спостерігалися б і в самок, і в самців.
Отже, таблиця, що містить Expectad Frequencies, будується на підставі емпірично зареєстрованих крайових сум, на підставі припущення про те, що значення розглядуваних змінних розподілені поза зв'язком одна з одною. Розподіли генотипів для різних статей відповідають розподілу генотипів у всій вибірці; розподіли статей для різних генотипів відповідають розподілу статей для всієї вибірки. Ця таблиця дає очікуваний розподіл статей у тому випадку, якщо нульова гіпотеза (що постулює відмінність зв'язку між змінними) вірна.
Нам лишилося порівняти спостережуваний і очікуваний розподіл. У таблиці це зроблено з використанням критерію
Отже, у нашому порівнянні обчислене значення критерію
Як це відображено на попередньому малюнку, зв'язок між розглядуваними змінними в нашому прикладі виявився незначимим (p=0,686; ми маємо справу з тим випадком, коли Statistica не ставить 0 перед десятковим роздільником). Отже, матеріал у нашому дослідженні представлений нерівномірно, але значимого зв'язку між статтю і генотипом ми не зареєстрували.
6.3. Порівняння розподілів за допомогою модуля непараметричної статистики
У попередньому пункті ми прийняли зареєстровані нами крайові частоти (загальна кількість самців, самок, а також представників різних генотипів) у нашій вибірці як задані. Ми припустили, що вони відображають різну кількість представників різних статей і різних генотипів у тій генеральній сукупності, з якої ми отримали нашу вибірку. Однак у деяких випадках має сенс інша постановка питання. А чи можемо ми на підставі нашої вибірки припустити, що різні статі і генотипи розподілені в генеральній сукупності, яку ми вивчаємо, нерівномірно? Для цього нам треба порівняти зареєстрований розподіл з іншим, де в кожній комірці міститься однакова кількість особин (легко зрозуміти, як його визначити: 57/10=5,7). Для цього нам доведеться зробити файл з таким розподілом.
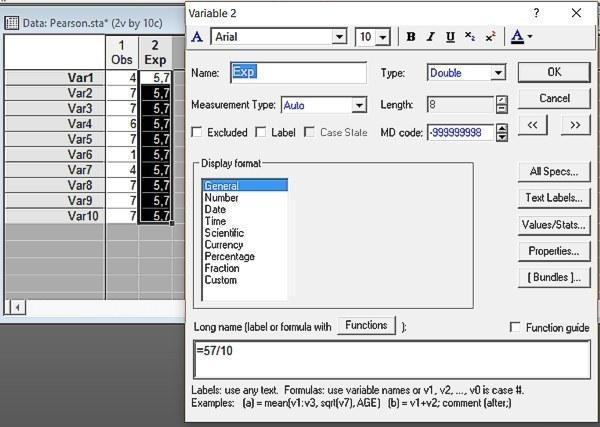
Рис. 6.3.1. У першому стовпці наведені зареєстровані частоти, у другому — очікувані частоти в тому випадку, якщо чисельність представників обох статей (а також різних генотипів) однакова
Для того способу аналізу, до якого ми плануємо приступити, важливо, щоб загальна чисельність обох розподілів була однакова. У нашому випадку ця умова виконується: ми згенерували очікуваний розподіл у перерахунку на зареєстровану кількість особин. Тим же способом можна порівнювати і два емпіричні розподіли, тільки при цьому потрібно перерахувати більший так, щоб кількість спостережень у ньому відповідала меншому.
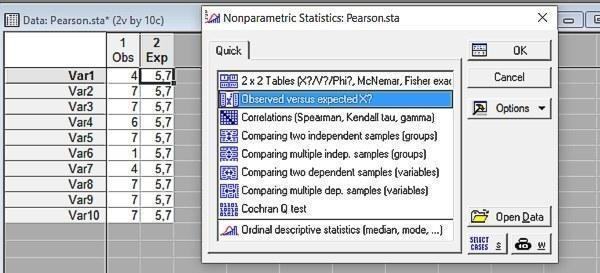
Рис. 6.3.2. Використовуваний нами метод належить до числа непараметричних
Лишилося вказати, які розподіли ми порівнюємо, і отримати результат.
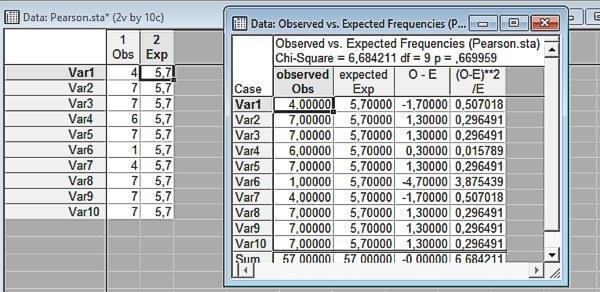
Рис. 6.3.3. Результат порівняння емпіричного і розрахункового розподілів. Різниця незначима
На останньому малюнку ми можемо побачити, як відбувається обчислення статистики
Отже, ми не знайшли підтверджень припущенню, що статі і генотипи жаб розподілені нерівномірно (а в минулому пункті — що вони пов'язані одне з одним). На підставі розглянутих даних у нас немає достатніх причин відкидати нульову гіпотезу. Важливе доповнення полягає в тому, що для розглядуваної проблеми відомо, що нульова гіпотеза невірна. На території вивчення частоти різних генотипів нерівні; для деяких генотипів досить часто спостерігається переважання самок чи самців. Однак ці твердження робляться не на підставі даних, що містяться у файлі Pelophylax_example.sta, а з урахуванням інших, значно представницькіших досліджень.
Таким чином, у нашому випадку нульова гіпотеза не відкинута. Як і в усіх інших випадках, це не означає, що вона вірна.