Шишкин М.А. Индивидуальное развитие и эволюционная теория // Эволюция и биоценотические кризисы. М.: Наука, 1987. - С. 76–124.
УДК 591.3:575.8
ИНДИВИДУАЛЬНОЕ РАЗВИТИЕ И ЭВОЛЮЦИОННАЯ ТЕОРИЯ
М. А. Шишкин
Палеонтологический институт АН СССР
Дарвиновская теория естественного отбора имеет своим объектом целостные живые организмы, возникающие в ходе индивидуального развития и ведущие борьбу за существование на всем протяжении жизненного цикла. Поэтому решение ключевой для этой теории проблемы происхождения адаптации неизбежно требует выяснения законов эволюции онтогенеза, т. е. законов становления тех механизмов, которые обеспечивают виду надежное осуществление его фенотипической нормы. С позиций сегодняшних знаний можно видеть, что многие возражения, выдвигавшиеся против классического дарвинизма, могли казаться существенными лишь потому, что вопросы эволюции индивидуального развития остались в этом учении не разработанными.
Совершенно иную основу имеет господствующая ныне генетическая («синтетическая») теория эволюции, означающая, по выражению И. И. Шмальгаузена (1968а, с. 20), замену дарвинизма генетикой. Абстрагирование от проблем онтогенеза лежит в самих ее предпосылках, поскольку материалом отбора здесь по существу мыслятся не организмы, а наследственные факторы или их комбинации, т. е. структуры, предшествующие развитию; их судьба и должна определять ход эволюции. Вопрос о влиянии фенотипической реализации этих факторов на характер отбора остается в стороне, если не считать того, что учитывается возможность их проявления или непроявления и число теоретически признается влияние на эти процессы генотипической среды. Такое положение дел достаточно хорошо иллюстрируется тем, что до недавнего времени можно было встретить работы с изложением генетической теории, в которых понятие онтогенеза вообще не употреблялось (Шеппард, 1970). С другой стороны, характерно, что множащиеся в последние годы критические высказывания в адрес этой теории в первую очередь подчеркивают упрощенность и недостаточность ее предпосылок для понимания эволюционной роли процессов развития.
Между тем история дарвинизма знает две серьезных попытки понять взаимоотношения между естественным отбором и индивидуальным развитием — теорию зародышевой плазмы А. Вейсмана и учение о стабилизирующем отборе И. И. Шмальгаузена и К. Уоддингтона. Предложенные ими решения диаметрально противоположны. Если у Вейсмана эволюционные изменения онтогенеза составляют лишь пассивный результат отбора элементов зародышевой плазмы, то у Шмальгаузена, наоборот, материалом для селективного преобразования генотипа служат аберрации онтогенеза. Генетическая теория унаследовала от Вейсмана лишь его редукционистское понимание наследственности — расчленение ее на независимые факторы (типичное для теорий наследственности XIX в.) и замену последними целостных организмов в качестве субстрата отбора. Глав ный же объединяющий принцип вейсмановской концепции, сделавший ее подлинно синтетическим построением, остался незамеченным. Он заключался в ясном понимании того, что механизм наследования свойств организма выражается в способе их онтогенетического осуществления, и потому признание передачи этих свойств независимыми носителями может означать только их независимую реализацию в онтогенезе. Иначе говоря, идея дискретной наследственной детерминации неизбежно предполагает мозаичное (преформированное) развитие. Вейсман построил такую модель развития (опираясь на гипотезу неравнонаследственного деления), и именно ее несостоятельность логически повлекла за собой крушение всей его теории. Однако выводы, казалось бы, следовавшие отсюда для понимания механизма наследственности, так и не были сделаны вплоть до появления теории стабилизирующего отбора.
Что касается этой последней теории, анализируемой дальше, то отношение к ней современного селекционизма неопределенно. Хотя в отечественной литературе она нередко характеризуется как высшее на сегодня достижение дарвинизма (Шварц, 1969; Галл, 1980), все же, как это верно отмечают (Кирпичников, 1974), ее признание зачастую носит чисто словесный характер. Причины этого в большой мере понятны. Теория стабилизирующего отбора с ее установкой на объяснение эволюции целостной организации является преемственной по отношению к классическому дарвинизму (ср.: Яблоков, 1981), но не к генетической теории. Их отношения с последней, как будет показано ниже, являются в действительности антагонистическими. Поэтому для господствующего направления современного селекционизма невозможно принять учение о стабилизирующем отборе по существу, не ревизовав при этом своих собственных основ.
Если удовлетворительная эволюционная теория должна быть по сути теорией эволюции онтогенеза и если дарвиновское учение содержит возможность для этого, то его основополагающие понятия необходимо должны быть выражены на языке индивидуального развития. Это касается наследственности, изменчивости, приспособления (как процесса), а также самого механизма естественного отбора. Такому же анализу должны подвергнуться и понятия генетики, используемые современным эволюционизмом, если описываемые ими явления имеют отношение к реализации фенотипов и их адаптации. Речь идет о том, чтобы выяснить, действительно ли то, что носит название аллелей, гомо- и гетерозигот и т. д., суть «чистые» характеристики наследственной структуры, составляющей субстрат отбора, или же мы имеем дело с особенностями морфогенетических систем, созданных самим эволюционным процессом.
Прежде чем перейти к этим вопросам, необходимо выделить некоторые наиболее значимые в эволюционном плане особенности индивидуального развития. Эволюция есть прежде всего смена поколений, ведущая к смене адаптивных норм, и переход к каждой новой норме означает неизбежно то или иное нарушение предыдущей. Поэтому в эволюции популяций и видов должны существовать поколения с преобладанием устойчивых по своему результату индивидуальных циклов (формирующих норму) и поколения, характеризующиеся онтогенезами с более неопределенным исходом. Закономерности перехода от второго типа к первому должны объяснять механизм адаптациогенеза (как это и трактуется теорией стабилизирующего отбора). Поэтому в онтогенезе нас будет в основном интересовать соотношение устойчивых и неустойчивых, или нормальных и аберрантных, путей развития.
НАСЛЕДСТВЕННОСТЬ, ИЗМЕНЧИВОСТЬ, ПРИСПОСОБЛЕННОСТЬ И ОРГАНИЗОВАННОСТЬ КАК ХАРАКТЕРИСТИКИ УСТОЙЧИВОСТИ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ
Характерным свойством живых существ является устойчивость их типичной морфофизиологической организации, обозначаемой как адаптивная норма (Шмальгаузен, 1940а). Это, собственно, и делает возможным существование таких типологических методов исследования, как сравнительная морфология, систематика и филогенетика. В ряду поколений это свойство организмов проявляется, кроме того, как их способность к самореализации, т. е. как устойчивость наследования адаптивной нормы. Эти две особенности иногда указывают в качестве самостоятельных характеристик живого (Кастлер, 1967), но в действительности они имеют одну и ту же основу — устойчивость осуществления нормы в ходе индивидуального развития.
В отношении нормальной организации как таковой эта основа самоочевидна, поскольку взрослое состояние организма, о котором идет речь, само является составной частью и итогом онтогенетического цикла. Столь же очевидна и связь между развитием нормы и ее наследованием, но здесь требуются некоторые разъяснения. В дарвиновской теории эволюции наследственностью называется передача фенотипических признаков в поколениях (Дарвин, 1951). Эта «передача» означает, конечно, воспроизведение признака заново, в ходе индивидуального развития, соединяющего в типичном случае гаметы родителей с фенотипом потомка; надежность результата этого процесса и определяет наследование. Поэтому, говоря о «наследственных» и «приобретенных» признаках, в действительности подразумевают лишь устойчивость или лабильность их онтогенетической реализации по отношению к условиям развития, и эти понятия предпочтительны, так как гораздо яснее передают суть дела (Woodger, 1953; Шмальгаузен, 1969, 1982; Шишкин, 1981, 1984а, б, в). Говоря словами А. Г. Гурвича (Gurwitsch, 1912; Гурвич, 1944), наследственность есть процесс осуществления типичного развития (ср.: Мейстер, 1934; Дубинин, 1973). Никакого иного содержания по отношению к фенотипам (для которых он и введен) термин «наследственность» не имеет. Обозначать им генетическую обусловленность признаков бессмысленно, так как все свойства организма являются продуктами взаимодействия генотипа и среды развития и в таком случае одновременно являются «унаследованными» и «приобретенными» (Johannsen, 1926; de Beer, 1963; Камшилов, 1972). Если же относить понятие наследственности к генотипам и их элементам, то не только меняется его исходный смысл, но оно попросту превращается в «вещь в себе», не поддающуюся иному определению, кроме тавтологического («наследственность есть передача наследственных факторов») [Распространенное утверждение, что объектом наследования является индивидуальная «норма реакции» (Johannsen, 1926; Dobzhansky, 1947) равносильно признанию наследования индивидуальных генотипов. Но последние непрерывно преобразуются в каждом поколении за счет ошибок репликации (в общем случае) или в ходе рекомбинации (при половом процессе)] . Этот важный момент приходится оговорить, поскольку вся история изучения наследственности несет на себе печать странного на первый взгляд дуализма, когда с одной стороны в ней видят свойство определенной категории признаков, а с другой — нечто существующее независимо от них и относящееся к зародышевой плазме (генотипу). Это противоречие исчезает лишь в одном случае — если полагать, что устойчивое воспроизведение (наследование) признаков обусловлено передачей через гаметы однозначно соответствующих им дискретных причинных факторов. Именно так и считала ранняя генетика, и это представление объективно остается фундаментом построенной на ней эволюционной теории, несмотря на стремление ее авторитетов (например, Майр, 1968, 1974) считать его изжитым.
Положение о том, что устойчивость организмов создается селективным процессом, является центральным для теории стабилизирующего отбора. Но если онтогенетическая устойчивость означает наследуемость, то очевидно, что и последняя есть продукт отбора. Этот вывод ясно формулируется указанной теорией в виде понятий о превращении лабильных изменений в наследственные, о создании отбором наследственных механизмов, замене внешних факторов развития внутренними (Шмальгаузен, 1940б, 1941, 1968б, 1982, с. 109, 110, 161, 214) и, наконец, представления о генетической ассимиляции адаптивных признаков (Waddington, 1953, 1957). Между утверждением, что наследственные изменения создаются отбором, и обычным представлением, что они создаются мутированием, лежит непроходимая мировоззренческая пропасть, которая должна остановить каждого, кто хотел бы согласовать суть учения Шмальгаузена—Уоддингтона с генетической теорией. Вдумаемся в смысл более привычного второго утверждения. Если речь в нем идет просто о генетических изменениях, то их объяснение мутациями представляет собой тавтологию. Если же имеются в виду изменения фенотипа, устойчиво сохраняемые в поколениях, то оно заведомо неверно. Из опыта экспериментальной генетики хорошо известно, что элементарные малые мутации не гарантируют устойчивого аффекта (их проявление всегда лабильнее нормы) и, более того, могут вообще не проявляться. Сохранение фенотипической нормы при непрерывном рекомбинировании генотипа в ксеногамных популяциях особенно хорошо иллюстрирует последнее. Таким образом, наследственность (устойчивость) и генетическая обусловленность — это разные вещи (ср.: Камшилов, 1967).
Все сказанное неизбежно ведет к переоценке другого привычного постулата генетического селекционизма о «наследственной изменчивости» как материале естественного отбора. Фактически он также основан на отождествлении признаков и их наследственных факторов. Он подразумевает, что элементарные (неприспособительные) фенотипические вариации особей должны наследоваться, если они обусловлены их генетическими различиями. Но на самом деле, как уже сказано, наличие генетической гетерогенности особей само по себе ничего не говорит о характере ее фенотипического выражения. Она может или вообще не проявляться (под покровом нормального фенотипа, см.: Четвериков, 1926), или проявляться неустойчиво (как это типично для мутаций). Теория стабилизирующего отбора, исходящая из этих фактов и рассматривающая само свойство наследования как продукт эволюции, естественно, должна считать сырым материалом последней неустойчивые признаки, т. е. лабильные элементарные реакции (морфозы) индивидуально различающихся геномов (Шмальгаузен, 1982; Шишкин, 1981, 1984а, б). Именно совокупность таких реакций, осуществляемых на гетерогенной основе, соответствует дарвиновскому понятию неопределенной изменчивости, которое позволяет говорить об изменениях, не только не адекватных факторам среды, но и неупорядоченных по характеру воспроизведения у ближайшего потомства.
Эволюционное происхождение биологической устойчивости может быть рассмотрено еще в одном аспекте. Само это понятие имеет много синонимов, издавна используемых как раз для обозначения явлений, требующих эволюционного объяснения. К их числу относятся целесообразность, приспособленность (Эшби, 1959, 1962; Шмальгаузен, 1968а, с. 139) и уравновешенность со средой (Спенсер, 1899).
Все они означают свойство индивидуумов реагировать на внешние возмущения таким образом, чтобы сохранять свою нормальную жизнеспособность, включая успешное размножение. Адаптивная организация — это организация, способная к персистированию (Wake et al., 1983). Историческое выживание наиболее приспособленных означает сохранение и создание отбором все более устойчивых типов организации, способных противостоять максимально широкому спектру возмущений. Чем шире и разнообразнее этот спектр, тем большее число нейтрализующих ответных реакций требуется от организма, чтобы в итоге он мог реализовать одно из допустимых для него изоморфных нормальных состояний (закон необходимого разнообразия; Эшби, 1959). Эти реакции должны быть скоординированы, ибо устойчивость системы невозможна без взаимодействия ее частей (Bertalanffy, 1969). Таким образом, рост приспособленности (устойчивости) в ходе отбора неизбежно ведет к усложнению и повышению интегрированности морфофизиологической организации. Этот дарвиновский принцип, постоянно оспариваемый, начиная от К. Негели и вплоть до современных эволюционистов (Wright, 1964), логически вытекает из рассмотрения организма как целостной системы; но он и в самом деле становится необъяснимым, как только мы пытаемся заменить организмы в качестве объектов отбора мозаикой их наследственных факторов.
Все виды, поскольку они обладают адаптивной нормой, одинаково приспособлены к своей среде обитания (т. е. к своему спектру допустимых возмущений) и, следовательно, равноценны в том качестве, которое можно назвать их относительной устойчивостью. Однако они могут быть в принципе сопоставлены и по абсолютной устойчивости, т. е. степени того разнообразия внешних факторов, эффект которых они в состоянии релаксировать. Этот показатель, как видно из вышесказанного, является мерой их организованности, т. е. и мерой прогресса. Чем более хаотичны и непредсказуемы колебания факторов используемой среды обитания, тем выше требования к сложности самого организма, и, наоборот, чем среда однороднее, чем они меньше. Простыми (предсказуемыми) являются, в частности, колебания высокоупорядоченных внутриорганизменных сред, используемых паразитами, что и объясняет их тенденцию к дегенерации.
Поскольку установкой физиологического поведения организмов является регуляция в сторону нормы, то оно направлено на противодействие необратимым изменениям, которые и составляют суть эволюции. Устойчивая система (истинно равновесная, или же квазиравновесная, какими являются живые организмы и вообще открытые системы), пока она остается таковой, по определению, «не запоминает» своих флюктуаций (модификаций) и, значит, не эволюирует. Причина эволюции лежит в нарушении устойчивости (Спенсер, 1899), т. е. в выходе значений динамических переменных системы за пределы, позволяющие регуляцию целого. Восстановление устойчивости на новом уровне (т. е. нового равновесия с измененной средой) происходит лишь с помощью естественного отбора, который в силу этого, строго говоря, всегда является стабилизирующим. Он представляет собой механизм надорганизменной регуляции индивидуальной устойчивости.
Идеальная устойчивость, т. е. способность ответить флюктуацией на любое внешнее или внутреннее воздействие, остается, конечно, недостижимой для организмов, но чем более высок ее абсолютный показатель, тем менее они уязвимы для прямой элиминации (все более уступающей место дифференциальному размножению) и тем больше возрастает их способность предварять любое объективно необходимое элементарное эволюционное изменение соответствующей адаптивной модификацией, т. е. отчасти смоделировать его из «наличных возможностей» своей морфогенетической системы. В этом смысл утверждения Г. Спенсера (1899) о том, что в процессе эволюции органического мира естественный отбор уступает место прямому приспособлению; хотя оно и неверно буквально, сама тенденция здесь понята правильно. В этой же возможности предварительного моделирования состоит и суть идеи «органического отбора» Моргана—Болдуина, обосновывающей эволюционную роль модификаций (Шишкин, 1984б).
Поскольку рост абсолютной устойчивости, или приспособленности, сопряжен с усложнением организации, т. е. движением ко все менее вероятному состоянию, то эволюция уводит организмы все далее от термодинамического равновесия, что возможно лишь за счет все более высокого уровня потребления энергии извне. Таким образом, рост организованности (устойчивости) связан с увеличением энергетических затрат, и скорость продукции энтропии является ее существенным показателем (Goodwin, 1970). Эти затраты окупаются теми самоочевидными преимуществами, которые дает высокая приспособленность в борьбе за существование.
УСТОЙЧИВОСТЬ НОРМЫ И ПРИНЦИПЫ ТЕОРИИ НОРМАЛЬНОГО РАЗВИТИЯ
Устойчивость развития типичной организации, обеспечивающая ее самоподдержание (приспособленность) и самореализацию (наследуемость), а также связь этого явления с регуляционными механизмами онтогенеза отмечены эмбриологией давно. Уже К. Бэр (Ваеr, 1828) сравнительным путем установил уменьшение эмбриональной изменчивости в ходе развития, показывающее, что свойством последнего является устремленность к определенному конечному состоянию. Для механики развития, возникшей в конце XIX в. и поставившей своей задачей экспериментальное установление и локализацию непосредственных действующих факторов морфогенеза, это свойство оказалось непреодолимым препятствием при попытках построить общую теорию развития на основе каузально-аналитического метода (Бляхер и др., 1935).
Результат процесса не поддавался интерпретации в качестве суммы эффектов определенных начальных причин, показывая значительную независимость от их вариаций. Устойчивость этого результата по отношению к способам его достижения (Roux, 1895), продемонстрированная многими экспериментами, сделала невозможным принятие мозаичной неопреформистской концепции В. Ру, сводящей развитие к независимой дифференциации начального набора зачатков. Теоретические следствия из явлений саморегуляции развития и принципиальной неразложимости последнего на независимые причинно-следственные цепи были осознаны Г. Дришем (Driesch, 1908, Дриш, 1915), который охарактеризовал зародыш как «гармоническую эквипотенциальную систему», т. е. комплекс частей с одинаковыми возможностями (проспективными потенциями), управляемый как целое в своих преобразованиях внутренним нематериальным упорядочивающим фактором — энтелехией. Этот неразложимый фактор определяет фактическую судь бу (проспективное значение) зачатков в соответствии с их положением в целом и контролирует согласованность их изменений в течение всего развития, в том числе и на этапах самодифференцировки, когда экспериментально между зачатками не обнаруживается взаимозависимостей. В этой концепции важно, конечно, не конкретное «решение» проблемы (которое просто заменено символом энтелехии), а ясное понимание того факта, что развитие есть целостный процесс, свойства которого сверхсуммативны и устойчивы по отношению к составляющим его элементам. Это было началом системного подхода к развитию, в основе которого лежит аристотелевский принцип «целое существует раньше частей». Его правомерность была показана еще ранее в таком сугубо эмпирическом обобщении, как закон Бэра (Ваеr, 1828), который буквально утверждает то же самое (общее в развитии возникает раньше специального); но лишь начиная с Дриша он стал использоваться как основа теории развития. На нем строится современное учение об эмбриональной детерминации, которое, однако, в отличие от теории Дриша признает фактор целостности материальным и познаваемым (Gurwitsch, 1910, 1912; Светлов, 1964, 1978; Белоусов, 1963).
Таким образом, поиски причинного объяснения помехоустойчивости нормального развития привели эмбриологию к рассмотрению этого процесса как иерархической системы (целое и его части), управляемой своим верхним уровнем, т. е. свойствами целого. Позднее (и по существу на три же самой основе) было получено и историческое объяснение указанного свойства развития. Мы снова имеем в виду теорию стабилизирующего отбора (Шмальгаузен, 1940б, 1941, 1968б, 1982), опирающуюся на понятие адаптивной нормы. Последняя (т. е. нормальный фенотип) исторически меняется много медленнее, чем способ ее онтогенетической реализации, который непрерывно преобразуется отбором в сторону повышения его надежности. Тем самым повышаются возможности варьирования процессов, составляющих развитие, без ущерба для устойчивого осуществления нормы. Последняя выступает здесь как фактор целостности, управляющий (через отбор) изменением своих морфогенетических механизмов и определяющий допустимое пространство индивидуального варьирования их элементов (Шишкин, 1981). Принципиальное тождество эмбриологического и исторического объяснений устойчивости представляется здесь самоочевидным.
Нас, однако, интересует вначале эмбриологическое объяснение, т. е. схема причинно-следственных отношений, пригодная для описания отдельного цикла развития. Но сперва необходимо остановиться на общих теоретических требованиях, которым должно удовлетворять это описание.
Устойчивость результата нормального развития означает целенаправленность этого процесса. Оба эти определения характеризуют одно и то же — способность к саморегуляции конечного состояния. Целеполагающее (телеономическое) поведение устойчивой материальной системы проявляется в том, что, будучи выведена из состояния равновесия, она реагирует так, что в конечном итоге возвращается к нему. Соответственно для описания таких процессов в физике и химии используются финалистические формулировки (принцип Ле-Шателье и т. п.). Для закрытых систем состояние устойчивости соответствует термодинамическому равновесию, а в открытых, включая живые организмы, находится на удалении от него и характеризуется как «проточное равновесие» (Bertalanffy, 1949), или «устойчивое неравновесие» (Бауэр, 1935), или же как стационарное состояние. Движение к любому типу равновесия, или «поиск цели», осуществляется через замкнутые циклы событий с обратной связью, когда, например, элемент А при возмущении воздействует на Б таким образом, что последний своим изменением корректирует состояние А в сторону значения, вызывающего уменьшение дальнейшей его коррекции. Система «управляется своей ошибкой» (Эшби, 1962), и ее устойчивость зиждется на взаимодействии ее элементов. В открытых системах циклы коррекции осуществляются постоянно; в закрытых же амплитуда взаимодействий затухает по мере роста энтропии и устанавливается «устойчивость по отношению к точке» (Goodwin, 1970).
Представление о целенаправленном поведении системы не означает, конечно, признания зависимости протекающих событий от будущих условий. Оно лишь отражает тот факт, что конечные результаты элементарных изменений в системе определяются общими свойствами ее самой и не могут быть сведены к прямым механическим следствиям этих изменений. Система как целое или вообще не реагирует на элементарное воздействие, или переходит в одно из своих альтернативных состояний (модификаций). Другими словами, телеономическая зависимость обнаруживается при сопоставлении событий или свойств, отвечающих разным иерархическим уровням системы, а именно при сопоставлении ее медленно меняющихся параметров (характеризующих ее целостное поведение) и быстро варьирующих значений ее элементов (динамических переменных). Финалистическая форма описания таких соотношений отражает принципиальную невозможность их каузального описания, ибо свойства целого несводимы однозначно к состояниям его элементов. «То, что представляется как устойчивая структура определенного уровня, на самом деле удерживается непрерывным обменом компонентов ближайшего нижнего уровня» (Bertalanffy, 1969), т. е. одно и то же свойство целого сохраняется при разных комбинациях элементарных взаимодействующих причин. Другими словами, для межуровневых отношений характерна резкая асимметрия причин и следствий (Белоусов, Чернавский, 1977), не свойственная процессам, поддающимся каузальному описанию [С этой точки зрения виталистическая концепция развития Дриша предстает как характерная реакция естествоиспытателя, констатирующего отсутствие привычной для него однозначной причинной зависимости между связанными явлениями и не видящего другой альтернативы для описания их связи, кроме введения нематериальных факторов]. Поэтому нельзя согласиться с распространенным представлением, что финалистическая и каузальная формулировки — это лишь два равноправных способа описания изменений в одних и тех же циклических причинных цепях. Иерархически равноправные элементы таких циклов не имеют асимметричных соотношений между собой и, наоборот, свойства разных уровней системы не связаны каузальной зависимостью.
Все сказанное имеет прямое отношение к пониманию механизма индивидуального развития. Мы приходим к выводу, что его каузальное объяснение возможно лишь в том случае, если вся цепь причинных событий, ведущих к целостному конечному результату (нормальной организации) будет представлена как последовательность равноправных (одноуровневых) целостных состояний. Наличие в ходе развития цепи таких состояний с устойчивыми характеристиками есть не только теоретическое требование, но и экспериментально установленный факт. Здесь действительно обнаруживаются последовательные периоды, характеризующиеся внутренней целостностью (топологической изоморфностью) и направленностью преобразований, при пониженной чувствительности к экспериментальным нарушениям (Светлов, 1960; Белоусов, 1979). В то же время индивидуальное развитие, так же как и любое другое необратимое изменение, должно быть связано с движением через фазы нарушения устойчивости (Bertalanffy, 1969; Волькенштейн, 1981б). Эти фазы так же обнаруживаются в индивидуальном развитии в виде «чувствительных периодов», характеризующихся лабильностью детерминации и трансформацией топологических рисунков. Следовательно, общая теория нормального онтогенеза должна минимально включать в себя следующие предпосылки.
1. Развитие есть цепь обусловливающих друг друга структурно целостных состояний. 2. Каждое из них на период своего существования определяет ход и согласование отдельных морфогенетических процессов (т. е. действует как «энтелехия» по Дришу). 3. Реализация этих процессов каждый раз имеет следствием определенное нарушение устойчивости целого и восстановление ее затем на новом уровне, контролирующем дальнейшую дифференциацию. 4. Поскольку в ходе развития организация зародыша усложняется, каждое новое состояние целостности стабилизируется на все большем удалении от истинного равновесия.
По-видимому, единственной концепцией развития, согласующейся в основе с этими предпосылками, является на сегодня теория биологического поля, выдвинутая А. Г. Гурвичем (Gurwitsch, 1922; Гурвич, 1944). Понятие физического поля, т. е. пространства, свойства которого определяют поведение находящихся в нем частиц, хорошо отвечает представлению об искомом материальном факторе целостности, контролирующем всю совокупность процессов развития. Как верно отметил П. Г. Светлов (1964), принцип поля ясно выражен уже в концепции Дриша, указавшего, что проспективное значение отдельного элемента в развитии есть функция от его положения в целом [Это представление Дриша вступало в очевидное противоречие с его определением фактора целостности как «неэкстенсивной» величины, не имеющей пространственных характеристик] . Теория Гурвича, если отвлечься от ее дальнейшей детализации (1944), связанной с идеей клеточного поля, позволяет приблизиться к пониманию простых и общих законов, лежащих в основе онтогенетического процесса. Принимается, что, начиная с яйцеклетки, зародыш образует вокруг себя анизотропное векторное поле, структура которого предопределяет результат развития на ближайшем бесконечно малом его этапе. После заполнения пространства поля последнее «изживает себя» и реорганизуется в поле с новыми параметрами, обусловленными конечным состоянием зародыша, достигнутым, на предыдущем этапе. Этим создается установка развития на новый ближайший отрезок и т. д. В ходе процесса происходит также формирование полей отдельных зачатков, подчиненных полю целого. В этой концепции фактор целостности (поле), непрерывно направляя развитие, сам в то же время является непрерывной функцией от пути, пройденного субстратом его воздействия. Мы имеем здесь «закон Дриша в дифференциальной форме» (Белоусов, 1979), т. е. направляющее целое рассматривается уже не как конечная цель, а как свойство последовательных стадий, преобразуемое по законам причинности [Отчасти такой ход событий осознавался уже Дришем (1915), указывавшим, что выполнение каждого этапа развития ведет к «изменению очередной задачи энтелехии». По существу речь здесь идет об изменении свойств самого фактора целостности]. Развитие предстает как лавинообразный процесс с положительной обратной связью между зачатком и его полем при целенаправленном поведении зачатка по отношению к каждому новому установившемуся состоянию поля. Хотя доказательства теории относятся скорее к частным морфогенезам и касаются в основном пространственной стороны изменений как наиболее доступной для изучения, они представляются очень вескими. На многих примерах показано (Гурвич, 1944), что детерминация целого зачатка осуществляется при неопределенном состоянии слагающих его элементов. Последние лишь статистически детерминированы как совокупность («нормированы» по Гурвичу), подчиняясь полю целого. К числу хорошо известных примеров относится случайный характер пространственно-временного распределения отдельных митозов по отношению к осям симметрии в таких зачатках, как развивающийся корешок лука или сетчатка глаза; общий результат этих случайных событий остается упорядоченным. Реальность поля наглядно доказывается явлением «динамической преформации» зачатка, когда ориентировка клеток его стенки определяется не фактической конфигурацией последней, а силовой поверхностью вне ее, соответствующей ее проспективным очертаниям, приобретаемым на ближайшей следующей стадии.
Хотя построение общей теории онтогенеза — дело будущего, она, несомненно, не сможет обойтись без системных принципов, положенных в ее основу Гурвичем. В пользу этого говорят уже простейшие соображения. 1. Развивающийся организм есть целостная динамическая система, исключающая однозначное состояние составляющих ее процессов или лежащих в их основе начальных элементов. 2. В каждый момент развития не существует иной целостности, чем та, которая свойственна зародышу на этой стадии. Поэтому идея преобразуемого целого становится неизбежной.
МНОЖЕСТВЕННОСТЬ НАЧАЛЬНЫХ СОСТОЯНИЙ КАК ОСНОВА УСТОЙЧИВОСТИ НОРМЫ
В устойчивой системе сохранение ее параметров основано на непрерывном изменении ее взаимодействующих частей. Тем самым регуляция этих параметров при каждом возмущении и, следовательно, процесс их достижения из начального неравновесного состояния, составляющий телеономическую модель индивидуального развития, осуществляются одинаково эффективно при множестве вариантов исходных значений элементов системы. Эту независимость конечных свойств динамической системы от ее начального состояния называют эквифинальностью в широком значении термина (Bertalanffy, 1969) [Эквифинальность понималась Дришем (1915) как способность особей одного вида к различным путям регенерации целого организма при одном и том же экспериментальном повреждении, т. е. речь шла о явлении, близком к тому, которое в сравнительной эмбриологии именуется «окольным развитием» (сходство начала и конца типичного развития у разных форм при различии путей в промежутке). В значении независимости итога процесса от начального состояния это понятие было использовано Л. Берталанфи (Bertalanffy, 1949, 1969), который считает такое поведение свойством лишь открытых систем. Это представляется верным лишь в том смысле, что для открытой системы теоретически возможно сохранение любого параметра, тогда как в закрытой возмущение извне ведет к определенным необратимым изменениям. Но на деле для открытых систем способность к регуляции распространяется каждый раз не на все их свойства; например, развитие морских ежей из изолированных бластомеров или регенерация асцидий из оперированных особей ведет к формированию хотя и нормальных, но уменьшенных организмов (Дриш, 1915). С другой стороны, эквифинальность закрытых систем в отношении максимума энтропии всегда является абсолютной]; но для наших задач удобнее называть ее принципом множественности начальных состояний. В развитии организмов эта закономерность имеет множество очевидных проявлении; особенно наглядны случаи, когда для вида возможны разные способы размножения (например, половое и вегетативное у асцидий, кишечнополостных и т. д.), при которых начальные стадии развития не имеют между собой ничего общего, хотя результат его тождествен. Сюда же относятся и все экспериментальные факты по онтогенетической регуляции, в частности случаи регенерации целого организма из фрагментов специализированных тканей (плоские черви, немертины и т. д.) или явления самосборки зародыша (например, гаструлы морских ежей) из изолированных клеток с последующим восстановлением нормального развития (Светлов, 1964, 1972). Вообще, не только реализация целого организма, но и выполнение любого морфогенетического акта в нормальном развитии основаны на известной независимости его результата от начальных условий. Например, нормальная индукция выполняется при значительных колебаниях в массе и времени взаимодействия членов индукционной системы, вариациях в чувствительности реактора, концентрации и составе активирующих веществ и т. д. (Шмальгаузен, 1964, 1982). Иными словами, все развитие построено на относительно устойчивых актах с «множественным обеспечением», если понимать под этим термином любые, а не только качественные различия в осуществлении одного и того же морфогенеза. В общей системе развития эти относительно устойчивые события составляют промежуточный иерархический уровень (или уровни) между однозначно детерминированным целым (взрослой нормой) и лежащими в основании системы статистически обусловленными элементарными процессами, например, клеточными делениями в «регуляционных» онтогенезах. В этом и состоит принцип нормировки Гурвича: детерминация и формирование отдельного зачатка не связаны с жесткой фиксацией начального состояния его элементов.
Принцип множественности начальных состояний имеет далеко идущие следствия для теории нормального развития, относящиеся уже к той ее области, которая исторически выделилась в учение о наследственности. Двигаясь в глубь развития вплоть до зиготы, мы должны будем заключить, что устойчивость реализации взрослой нормы (т. е. ее наследуемость) не может быть сведена к фиксированной совокупности состояний каких-либо клеточных единиц, в том числе и единиц хромосомного аппарата (генома) зиготы или яйцеклетки. Эта совокупность должна быть детерминирована лишь статистически (нормирована), т. е. должна сохранять неопределенность в пределах, допускающих нормальное (эквифинальное) завершение развития. Именно это и утверждается теорией стабилизирующего отбора. Согласно Шмальгаузену (1982, с. 84, 174), устойчивость организации не есть свойство элементов наследственной субстанции, а выражение взаимодействия частей, участвующих в развитии. Организм «устойчивее своего генотипа» (если понимать последний как набор определенных состояний хромосомных единиц). Этот один из важнейших выводов теории, основанный на обобщении эмпирических данных, является в то же время дедукцией из положения, что свойства системы несводимы к свойствам ее элементов. Адаптивная норма детерминируется лишь целостной видоспецифичной структурой зародышевой клетки, которой соответствует множество взаимозаменяемых вариантов генома, способных реализовать данную норму в типичных условиях развития. Для этой совокупности вариантов можно ввести понятие «генотипической нормы».
Какие факты подтверждает рассмотренный вывод? Прежде всего, установленная С. С. Четвериковым (1926) генетическая гетерогенность («насыщенность мутациями») природных амфимиктических популяций, скрытая под покровом адаптивной нормы и выявляемая инбридингом. Непрерывное перераспределение элементов индивидуальных геномов в ходе рекомбинации в поколениях не меняет исхода их фенотипической реализации в основной массе. Однако принцип множественности начальных состояний не содержит запретов, которые ограничивали бы вариабельность начальных факторов развития разрешающей способностью гибридологического (менделевского) анализа. Мы вправе предположить, что и в сообществах, представляющихся на этом уровне однородными (в чистых линиях самоопылителей и клональных популяциях), в действительности всегда существует скрытое генетическое разнообразие. В том, что это так на самом деле, убеждают многочисленные опыты по выращиванию в экстремальных условиях жестко отселектированных сортов автогамных растений (злаков, бобовых) и партеногенетически размножающихся насекомых (тли). При этом всегда обнаруживается разнообразие индивидуальных физиологических реакций, среди которых наиболее жизнеспособные варианты поддаются закреплению отбором, что сопровождается появлением у них устойчивых морфологических особенностей (Самохвалова, 1951, 1954; Шапошников, 1961, 1966; Агаев, 1978). Хотя эти опыты толкуются по-разному, кажется совершенно очевидным, что речь идет о генетической изменчивости, получившей фенотипическое выражение вследствие выхода условий среды за рамки, допускающие нормальное (эквифинальное) развитие [Эти факты противоречат обычному мнению о невозможности отбора в чистых линиях, основанному на классических опытах Иогансена (1933) с фасолью. Учение о генетической ассимиляции неустойчивых признаков позволяет теперь дать этим опытам иную интерпретацию. Для того чтобы лабильная реакция могла быть закреплена отбором, недостаточно одной лишь скрытой гетерогенности ее носителей. Необходимо также: а) чтобы последние осуществляли эту реакцию в одних и тех же уклоняющихся условиях и б) чтобы отбор вначале велся в этих же самых условиях. Оба эти требования в опытах Иогансена не соблюдались, и индивидуальные причины одинаковых модификаций размеров зерен не контролировались. Приблизительным подобием такого опыта была бы попытка закрепить какой-либо термоморфоз у дрозофилы с помощью его проявлений, полученных при противоположных уклонениях температуры развития, и ведя отбор среди непосредственных потомков, выращенных при нормальных условиях]. Результаты этих опытов принципиально ничем не отличаются от результатов, полученных при генетической ассимиляции (стабилизации) индивидуальных структурных и физиологических морфозов у ксеногамных организмов, таких, как дрозофила (Камшилов, 1941; Waddington, 1957). Из этого можно заключить, что при любом способе размножения адаптивная норма действительно реализуется на основе множественности допустимых состояний генома зародышевой клетки (или вегетативного зачатка).
Эта вырожденность соответствия между вариациями генетической основы и результатом нормального развития составляет, как уже сказано, один из важнейших принципов теории стабилизирующего отбора. По ее представлениям, устойчивость нормы выражается в создании отбором регуляторного эпигенетического механизма, способного в широких пределах нивелировать (канализировать) вариации генетических факторов и условий среды. Таким образом, формируя адаптивную норму, отбор неизбежно должен повышать и допустимую генетическую вариабельность в ее основании, как это и наблюдается в действительности. Итог нормального развития не сводим к фиксированной сумме начальных причин. Напротив, для генетической теории эволюции, пытающейся описывать фенотипы в терминах конкретных генов, логически ожидаемым результатом отбора является создание в популяциях максимальной однородности по всем отбираемым аллелям или их сочетаниям. Несоответствие этого ожидания реальной генетической структуре нормы вынуждает прибегать к различным дополнительным гипотезам (о балансе отбора и мутационного процесса, разнонаправленном или частотно-зависимом отборе и т. д.), само обилие которых (Kojima, 1971; Солбриг, Солбриг, 1982; Айала, 1981) говорит о непреодолимости возникающих затруднений.
ЭПИГЕНЕТИЧЕСКАЯ СИСТЕМА КАК ВЫРАЖЕНИЕ СВЯЗИ ГЕНОТИПА И ФЕНОТИПА
Адаптивная норма не исчерпывает возможностей развития индивидуальной зародышевой клетки. Вокруг нормального фенотипа лежит область разнообразных неустойчивых уклонений (морфозов), возникающих при нарушениях условий развития. Совокупность таких реакций, реализуемых на основе гетерогенной совокупности зигот (способных в обычных условиях к нормальному формообразованию), образует «мобилизационный резерв» популяции, т. е. скрытую изменчивость, составляющую, по теории Шмальгаузена (1941, 1968б), потенциальный материал эволюционных преобразований (Шишкин, 1981, 1984а, б). Но наряду с этими уклонениями существует на первый взгляд и другой их источник — мутационные вариации генома зародышевой клетки, нарушающие развитие адаптивной нормы даже в обычных для нее условиях. Именно эта категория аберраций и рассматривается большинством эволюционистов как материал естественного отбора. Возникает вопрос: действительно ли речь здесь идет о двух разных категориях уклонений и как они соотносятся с нормальным развитием? В чем состоит общность функционирования «нормальных» и «мутантных» геномов, позволяющая рассматривать их как варианты единого видового генотипа? Ведь если единичные мутационные нарушения не выводят развитие за пределы видоспецифической области аномалий (Дубинин, 1966б, с. 240; Майр, 1968), то это с тем большей очевидностью относится к нарушениям, вызванным внешними факторами. Напрашивается мысль, что ограничения в обоих случаях одни и те же. В самом деле, параллелизм фенопроявления мутационных и модификационных изменений, наблюдаемый как в природе, так и экспериментально, а также параллелизм между наследственными (устойчивыми) и модификационными признаками у близких рас и видов составляет хорошо известный факт, положенный в основу ряда исторически связанных, хотя и глубоко различных эволюционных теорий (неоламаркистские концепции, идея «органического отбора» Моргана—Болдуина и теория стабилизирующего отбора). Эта общность дает нам право полагать, что вся присущая виду совокупность возможных путей онтогенетической реализации есть проявление устойчивых свойств целостной системы развития и что эволюционный процесс ведет к изменению структуры всей этой системы.
Если каждому виду действительно свойствен ограниченный набор вариантов онтогенетического осуществления, то очевидно, что он составляет специфическое «пространство возможностей», характеризующее поведение данной системы. Для отдельной зиготы это пространство дефинитивных состояний может быть выражено на плоскости полем (разорванным на дискретные участки), а пути его реализации — пучком расходящихся траекторий, из которых в типичном случае лишь одна (с ее конечными ответвлениями) соответствует адаптивной норме, понимаемой как более или менее узко ограниченный участок поля (рис. 1, а, б). Для всех зародышевых клеток вида указанное пространство является одинаковым (эквифинальным), и индивидуальные различия между ними (и прежде всего их геномами) заключаются лишь в относительной вероятности осуществления различных траекторий развития при данных внешних условиях. Все те варианты генома, для которых нормальный фенотип составляет наиболее вероятный (устойчивый) итог развития (рис. 1, б, n1–n3), могут быть выделены как генотипическая норма; остальные же составляют то, что обычно именуется мутациями, хотя на самом деле все геномы в популяции являются мутантами по отношению друг к другу. Любое возмущение системы развития, не разрушающее ее (т. е. позволяющее закончить развитие), может в таком случае лишь изменить выбор конкретной траектории, но не в состоянии дать результат, лежащий за пределами видоспецифичного пространства возможностей. Иначе говоря, реакция системы на возмущение будет всегда в конечном счете отряжать специфику ее самой независимо от того, имело ли место внешнее воздействие на ход развития или же нарушение структуры самой зародышевой клетки.
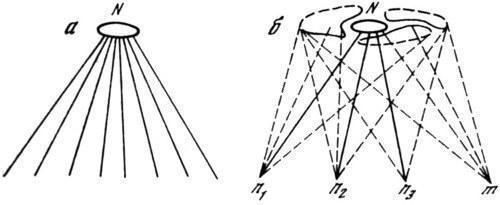
Рис. 1. Соотношения между зиготами и типами развития
а — эквифинальность типичного развития нормальных гетерогенных зигот; б — ограниченность видоспецифичного пространства возможностей развития при любых вариациях зародышевого генома. Сплошные линии — стабильный путь, прерывистые — лабильные пути развития; п1–n3 — нормальные зиготы, m— аномальная, N — адаптивная норма
Подобная теория системы развития, описывающая соотношения между индивидуальной структурой зиготы, условиями развития и конечным результатом, далеко еще не создана. И все же главные ее принципы ясны уже давно. Можно утверждать, что основу для нее заложило представление Р. Гольдшмидта (Goldschmidt, 1938, 1940) о сводимости всех фенотипических уклонений независимо от их начальных причин к количественным сдвигам внутри системы развития. Другим важным элементом является учение К. Уоддингтона (Waddington, 1957) об эпигенетическом ландшафте как структуре этой системы и о генетической ассимиляции как способе его перестройки. Мы попытаемся связать эти взгляды в одну непротиворечивую концепцию и проследить наиболее очевидные выводы из них, согласующиеся с экспериментальными данными генетики. Понятие видоспецифической системы развития используется нами вслед за указанными авторами наряду с такими синонимами, как «реактивная система» или «эпигенетическая система». Сходное значение имеет и общеупотребляемое понятие «нормы реакции» (Johannsen, 1926), обозначающее возможности эпигенетической реализации индивидуального генотипа.
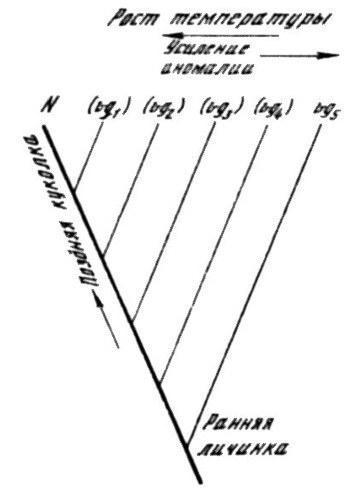
Рис. 2. Зависимость аллельных изменений фенотипа от количественных градаций фактора, управляющего переключением путей развития (на примере крыловой мутации vestigial у дрозофилы). Усиление теплового воздействия на личиночное развитие ведет к изменению фенотипа мутантов в сторону более слабых аллелей, вплоть до совпадения с нормой: момент уклонения развития от нормального пути смещается на все более поздние стадии
N — норма, vg1–vg5 — фенотипы аллельной серии
Представления Гольдшмидта о системных свойствах индивидуального развития исходят из уже упомянутого факта параллелизма мутационных и модификационных изменений (впервые экспериментально показанного с генетическим контролем Н. В. Тимофеевым-Ресовским в 1926 г.). Как следует из опытов с дрозофилой и другими организмами, эффект практически любой мутации, включая и его плейотропные проявления, может быть получен в виде морфоза (фенокопии) с помощью шоковых воздействий на тот или иной из чувствительных периодов развития. Отсюда Гольдшмидтом был сделан вывод, что оба типа явлений имеют одну и ту же физиологическую основу — неспецифические нарушения нормальной координации событий развития, а именно рассогласования скоростей морфогенетических реакций и изменения количеств, концентраций и времени взаимодействия реагирующих веществ. Возможности таких нарушений, допускающих завершение развития, имеют ограничения в рамках системы, а тем самым ограничен и набор осуществимых для нее фенотипических уклонений. Мутации в данной системе способны вызвать лишь те аномалии, которые могут быть получены и внешними воздействиями, приводящими к аналогичным количественным сдвигам в ней. Мутационные эффекты, включая и все их плейотропные проявления, являются не свойством гена, а эмбриологическим следствием времени, места и типа первичного нарушения развития, вызванного мутацией (Гольдшмидт, 1933; Goldschmidt, 1938, 1940, 1955; Камшилов, 1940; Шмальгаузен, 1982). Характер аномалий в итоге отражает целостные свойства самой системы, а не специфику действия того или иного хромосомного локуса.
Все это обосновывается многочисленными данными по феногенетическому анализу мутаций, например, таких, как Bar или vestigial у Drosophila melanogaster. Фенотипы всего ряда аллелей (вплоть до нормы) для каждой из них могут быть получены градацией температурных воздействий на личинку. Очевидно, что специфические аллельные изменения фенотипа являются здесь реакцией системы развития на чисто количественные изменения какого-то фактора, чувствительного к температуре среды. При этом чем меньше уклонение фактора от нормы (оно нейтрализуется у названных мутаций повышением температуры), тем позднее нарушается развитие (рис. 2). Так, при максимальном выражении эффекта vestigial дефект проявляется уже на стадии имагинального диска крыла, и оно не развивается дальше основания; при минимуме же редукции крыло формируется полностью, и лишь затем его пластинка частично лизируется у куколки (Goldschmidt, 1938, 1955). В ряде случаев природа количественного фактора, детерминирующего характер качественной фенотипической аномалии, может быть определена несколько более конкретно. Например, им может быть скорость роста и сегментации зачатка, как это показано для мутации aristopedia (у дрозофилы), связанной с превращением перистого отдела антенны (аристы) в структуру, подобную лапке конечности. У мутанта скорость роста имагинального диска повышена до уровня, свойственного в норме зачатку конечности; но при задержке роста с помощью колхицина развитие остается в рамках нормального пути (Балкашина, 1928; Goldschmidt, 1938, 1955). У мышей действие мутации Dh, вызывающей полидактилию задних конечностей, основано на замедлении гибели клеток апикального эктодермального гребня, вследствие чего пролонгируется его индуктивное действие на почку конечности. Напротив, мутация Os, ускоряющая отмирание гребня и сокращающая срок индукции, ведет к олигодактилии или даже редукции самой почки (Конюхов, Нончев, 1981). Примеры подобного рода заставляют многих исследователей признавать неспецифическое и непрямое воздействие генов на выбор реализуемого фенотипа (Wolpert, 1976; Alberch, 1982).
Если характер аномалии действительно не определяется прямо спецификой начального нарушения, то очевидно, что это должно обнаруживаться и при разных способах внешнего воздействия на развитие. Эксперименты с фенокопиями подтверждают это. Многие типы шоков, направленные на один и тот же чувствительный период развития, дают одинаковый результат, и, наоборот, один и тот же шок может вызвать качественно разные аномалии в зависимости от затронутой им фазы развития, а также его силы и продолжительности (Goldshmidt, 1955). С другой стороны, мы должны ожидать, что и разные по своей природе генетические нарушения будут приводить к тождественным аномалиям. Это действительно имеет место. Весь опыт экспериментальной генетики говорит о том, что уникальных по своему эффекту мутаций, по-видимому, не существует (Тимофеев-Ресовский, Иванов, 1966), и это означает лишь иную формулировку тезиса Гольдшмидта о неспецифичности воздействия мутаций на систему развития. Соответственно исследователями выделяются более или менее обширные «гетерогенные группы генов», имеющих различную локализацию, но вызывающих сходные или идентичные аномалии при мутировании. Характерный пример — группа minute у дрозофилы (укорочение торакальных щетинок), охватывающая около 60 локусов в трех хромосомах (Тимофеев-Ресовский, Иванов, 1966). Точно так же фенотип bithorax (удвоение среднегруди) может быть получен мутацией одноименного локуса в III хромосоме, комбинацией мутаций в трех разных хромосомах и, наконец, на основе «нормального» генома — с помощью шокового воздействия на личинку (Waddington, 1966). То же самое обнаруживается и при анализе природных фенотипических уклонений, где одни и те же их типы оказываются связанными с мутациями в разных хромосомах или просто с внешними воздействиями (например, фенотип Abnormal abdomen у дрозофилы; Голубовский и др., 1974). Наконец, хорошо известно, что одинаковые нормальные или аберрантные признаки у близких видов или географических рас одного вида очень часто реализуются на разной генетической основе; к таким обычным примерам относятся параллели в окраске лепестков у видов хлопчатника (Харланд, 1937) или крыльев у бабочек, в частности, желтая окраска у германской и итальянской форм Callimorpha dominula или белая — у английской и канадской форм Biston betularia (Goldschmidt, 1940; Шеппард, 1970). При генетическом анализе эти аналоги дают разные типы расщепления. В случае разных видов, где такой анализ обычно затруднен, подобные примеры принято связывать с параллельными, или гомологичными, мутациями; но на самом деле мы должны подразумевать здесь не наличие «того же гена», а сходство эпигенетических систем, допускающих осуществление одного и того же пути развития при разной генетической структуре (Goldschmidt, 1945).
Все эти факты ясно указывают на неопределенный в целом характер соответствия между первичными элементарными нарушениями процесса развития и его тем или иным конкретным итогом. Совокупность таких итогов сохраняет в пределах вида устойчивость по отношению к вариациям условий развития, т. е. ни один фенотипический признак не детерминируется фиксированной комбинацией состояний хромосомных локусов и внешних факторов. Этот контраст (асимметрия) между множественностью возможных условий развития и ограниченностью спектра его возможных исходов показывает, что здесь сопоставляются события, относящиеся к разным иерархическим уровням одной целостной динамической системы, обладающей устойчивым поведением. По существу феногенетический анализ мутантных аномалий вскрывает то же самое системное свойство развития, с которым столкнулась ранее экспериментальная эмбриология, — принципиальную невозможность сведения итога развития к определенной сумме начальных причин. Но вдобавок становится очевидным, что это касается не только нормального хода онтогенетического процесса, но и всего пространства его аберраций.
Все это, разумеется, не означает отрицания специфики первичной функции геномных локусов. Речь идет лишь о том, что ее изменения сами по себе не детерминируют признаков фенотипа, а выражаются в количественных нарушениях определенных параметров системы развития, на которые она реагирует качественным образом, изменяя выбор реализуемого в ней фенотипического результата. Тем более не имеется в виду идентичности первичных продуктов различных элементарных нарушений, вызывающих одинаковые аберрации. Различен должен быть и механизм их действия. При геномных изменениях — это те или иные цепи реакций, ведущие к нерегулируемым сдвигам в определенной фазе развития; внешние же стимулы нарушают эту фазу более прямым путем (Goldschmidt, 1955).
Изложенные представления подразумевают, что параметры морфогенетических процессов, составляющих развитие, характеризуются в каждой эпигенетической системе определенным рядом пороговых значений, в зависимости от достижения которых происходит выбор той или иной траектории развития. Необходимо уточнить, как этот выбор осуществляется, и попытаться понять, в чем состоит различие между нормальным и аберрантными путями развития в пределах одной системы. Ответ на эти вопросы в значительной мере проясняет предложенная Уоддингтоном (1947, 1960, 1970а; Waddington, 1957, 1966) модель эпигенетического ландшафта, описывающая общие свойства системы развития в виде серии ветвящихся наклонных долин, дивергирующих из общей начальной точки (рис. 3, 7). Эта модель имеет двоякий смысл. Во-первых, она отражает обычную интерпретацию развития как иерархию этапов все более специфической дифференциации зачатков, ведущей к последовательному ограничению их дальнейших формативных возможностей. При этом система траекторий, или долин, по которым «движутся» отдельные процессы, символизирует ограниченность и определенную дискретность путей дифференциации, возможных для дериватов данного зародыша или зачатка. Это свойство развития составляет хорошо известный факт, демонстрируемый, например, поведением эмбриональных тканей в эксплантатах (Светлов, 1964) Другой и более существенный смысл модели Уоддингтона состоит в том, что она позволяет изобразить путь нормального развития отдельного зачатка (и в пределе — всего организма) на фоне всего поля потенциальных возможностей его развития в пределах эпигенетической системы, свойственной данному виду. В этом случае нормальному пути соответствует глубокая долина, или креод (буквально — «необходимый путь»), а альтернативным возможностям — ее более пологие ответвления. Уклонение на любое из них связано с преодолением более или менее высокого порога (отделяющего эту долину от дна креода) и означает нарушение нормального хода развития. Но поскольку точки ответвлений соответствуют понижениям стенок креода (рис. 3), то они обозначают моменты относительной неустойчивости в выборе детерминации, т. е. чувствительные периоды, с воздействием на которые связано получение экспериментальных аберраций.
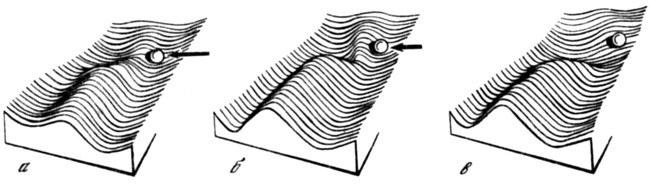
Рис. 3. Соотношения между строением эпигенетического ландшафта и характером повреждающего воздействия
а — уклонение развития на боковую долину за счет сильного внешнего воздействия (длинная стрелка); в — такое же уклонение под действием сильной мутации, вызывающей нарушение креода; б — промежуточное состояние (по Уоддингтону, 1957, интерпретация изменена)
Устойчивое (канализированное) развитие, или движение по креоду, ведущее к адаптивной норме, обеспечивается его регуляцией. Последняя выражается в том, что процесс, будучи отклонен тем или иным воздействием, снова «скатывается» в русло долины, если смещение не вышло за пределы ее склонов. Поскольку с течением нормального развития способность его к регуляции обычно падает, то это означает постепенное выполаживание креода. При наличии нескольких адаптивных норм система развития имеет несколько альтернативных креодов, выбор среди которых контролируется или факторами среды (при модификационном полиморфизме), или закономерным рекомбинированием хромосомного аппарата (например, детерминация пола). В отличие от нормы аберрантные пути развития, представленные выположенными долинами, имеют ограниченные возможности регуляции и их итог относительно лабилен (что является общим правилом для мутаций и неадаптивных морфозов; Шмальгаузен, 1968б, Уоддингтон, 1944, 1970а). Другими словами, дискретность этих путей по отношению к норме не означает их устойчивости. В свою очередь и сама эта дискретность не абсолютна и проявляется тем слабее, чем позже в развитии наступает необратимое уклонение. Например, она минимальна или отсутствует у слабых выражений таких мутаций у дрозофилы, как eyeless или vestigial (Goldschmidt, 1938; Рапопорт, 1943).
Таким образом, эпигенетический ландшафт характеризует видоспецифичное пространство возможностей развития, охватывающее области устойчивого течения процесса (креоды), области наиболее вероятных аберраций (боковые долины) и зоны с минимальной вероятностью осуществления (водоразделы между долинами, которые траектория развития всегда стремится покинуть). Эта структура отражает свойства целостной динамической системы, показывая, что ее реакция на возмущения зависит от того, какая точка ее пространственно-временного протяжения подвергалась воздействию. Чем ближе она к области креода, тем более вероятно, что весьма различные по своей природе воздействия будут одинаково забуферены и, наоборот, в зонах неустойчивости сходные причины могут иметь глубоко различные последствия. В более общей форме ход всего развития как многокомпонентной системы может быть описан движением точки в многомерном (фазовом) пространстве, где ее координаты в каждый момент времени соответствуют мерам отдельных взаимодействующих элементарных компонентов (Waddington, 1957). Креоды соответствуют наиболее устойчивым траекториям, способным притягивать к себе близлежащие точки; вся же совокупность возможных траекторий в этом пространстве составляет «фазовый портрет» системы (Белоусов, 1979; Белоусов, Чернавский, 1977), наглядным отображением которого и является трехмерная модель эпигенетического ландшафта. Демонстрируемая ею относительно простая упорядоченность системы развития есть «свойство высшего порядка» по отношению к функциям элементарных геномных единиц (Waddington, 1957, с. 34) и основывается на взаимодействии всей их совокупности.
Рассмотрим теперь характер реакций эпигенетической системы на элементарные возмущения. Очевидно, что такие воздействия не могут преобразовывать саму систему, а лишь меняют ее состояние. Они, либо изменяют выбор траектории в пределах ее ландшафта, либо поддаются регуляции и вообще не меняют исхода развития. В обоих случаях конечный результат определяется свойствами самой системы.
В случае воздействий, интенсивность которых превышает возможности регуляции канализированного развития, их эффект, очевидно, может иметь две количественных интерпретации. Либо уровень какого-то критического фактора, способного отклонить развитие в боковую долину, перерастает порог, допустимый в данной временной точке для нормального хода процесса, либо исчезает сам этот порог, отделяющий креод от данной долины (т. е. повышается чувствительность к переключающему фактору). Наконец, возможны оба типа изменений вместе. Очевидно, в этом и состоит суть тех количественных сдвигов, к которым, как показал Гольдшмидт, неизбежно должны сводиться последствия всего разнообразия элементарных воздействий на систему развития.
Исходя из сказанного, можно описать возможные результаты воздействия отдельного повреждающего фактора на индивидуальные циклы развития в нормальной гетерогенной популяции. Все зиготы одного вида принадлежат вариантам одной и той же эпигенетической системы; их генетические различия при одних и тех же условиях развития обусловливают индивидуальные детали моделировки ландшафта (разную высоту порогов между отдельными долинами, различия в степени выраженности последних). Для нормальных зигот эти различия минимальны, т. е. путь, ведущий к нормальному фенотипу, является для них наиболее устойчивым (канализированным) и обнаруживает в этом отношении лишь частные вариации, выражаемые как локальные различия в высоте защитных порогов креода.
Для элементарных воздействий, способных вызвать здесь уклонения развития от нормы, возможны три основных ситуации (Шишкин, 1984а, б). 1. Резкое изменение генома, приводящее к столь сильному снижению защитного порога на определенном участке креода, что независимо от вариаций его прежней высоты развитие неизменно уклоняется на один и тот же боковой путь (рис. 3, в). Это — идеальная мутация, наиболее удобная для генетического анализа, т. е. такое локусное изменение, которое при введении в. любой вариант генома вызывает с максимальной вероятностью определенную аномалию развития. Однако на деле результат все же должен оказаться не вполне устойчивым, поскольку сглаженность аберративных долин ландшафта сама по себе исключает эффективную регуляцию ими онтогенетических траекторий. И действительно, на практике выражение даже сильных «сырых» мутаций остается изменчивым (Уоддингтон, 1970а). 2. Противоположная ситуация — предельное сильное внешнее воздействие на ход развития, преодолевающее любой порог его устойчивости в данной временной точке и приводящее в данном цикле развития к тому же результату, что и сильная мутация (рис. 3, а). 3. Между этими двумя крайними случаями лежит огромная область промежуточных состояний, когда характер и сама возможность уклонения зависят от конкретного соотношения между особенностями эпигенетического ландшафта и условиями развития (рис. 3, б). Действие одной и той же мутации будет либо лежать ниже порога нарушения, либо преодолевать этот порог в различных точках креода — в зависимости от индивидуальных особенностей ландшафта (определяемых исходной конституцией генома) и колебаний факторов среды. И наоборот, одни и те же уклонения должны возникать при различных комбинациях внешних и внутренних условий развития. В этих случаях говорят о мутациях с неустойчивым выражением и проявлением, т. е. не показывающих при анализе правильного менделевского наследования.
Эта последняя, наиболее типичная ситуация соответствует реальной картине неопределенной изменчивости, наблюдаемой в природных популяциях. Общеизвестен факт отсутствия или редкости в них той категории аберраций, которая может быть охарактеризована как доминантные мутации с хорошим проявлением (Гершензон, 1941). При этом даже крупные однотипные аберрации при анализе оказываются связанными с разными хромосомами или частью индуцированными извне (например, Abnormal abdomen у дрозофилы; Голубовский и др., 1974). При наличии достаточно больших выборок таких фенотипов они показывают самые различные градации по устойчивости наследования — от соотношений, близких к менделевским, до полной потери проявления (например, фенотип «пятнистые глаза» у дрозофилы; Дубинин и др., 1937). Поэтому внутри таких групп изореагентов авторы часто вообще не решаются провести границу между наследственными (мутационными) и ненаследственными (модификационными) изменениями (Балкашина, Ромашов, 1935) или же ищут ее между линиями с минимальным наследованием аберрации и линиями с полным его отсутствием. Вполне очевидно, что речь здесь идет о границе, которой нет в природе. Все фенотипы одного класса представляют собой варианты реализации одной и той же онтогенетической траектории, различающиеся по степени устойчивости в силу того, что они обусловлены самыми различными сочетаниями индивидуальной генетической конституции и факторов среды. Понятия «мутации» и «модификации» на деле совершенно несопоставимы, так как первое относится к сравнению особей, а второе — к сравнению возможностей развития одной и той же особи. Все одинаковые фенотипы (как и любые другие) всегда генетически не идентичны и потому могут рассматриваться как скрытые мутанты по отношению друг к другу безотносительно к результатам гибридного анализа. С другой стороны, любой фенотип, оцениваемый на основе такого анализа как мутантный, представляет собой лишь одну из возможностей развития в пределах эпигенетической системы данной зиготы, т. е. одну из альтернатив (модификаций) по отношению к нормальной для вида траектории. Последнее особенно очевидно для тех случаев, когда возвращение на эту траекторию практически легко осуществимо путем изменения условий развития (например, мутации pennant, vestigial, Abnormal abdomen и др. у дрозофилы; Шмальгаузен, 1968).
Итак, гетерогенность однотипных аномалий развития, предсказываемая моделью эпигенетического ландшафта, подтверждается реальной картиной их наследования в природных популяциях. Точно так же подтверждается и другой вывод из этой модели — что одни и те же внешние возмущения будут, как правило, вызывать разнонаправленные отклонения от нормального развития в соответствии с индивидуальной конституцией затрагиваемых ими зигот. Усиление изменчивости при нарушении нормальных условий, отмеченное еще Дарвином, составляет хорошо известный факт, сравниваемый с разложением луча света при прохождении через призму (Лобашев, 1947). Переход популяции под действием экстремальных условий от фенотипического единообразия к проявлению разнонаправленных вариаций был обозначен как вскрытие «мобилизационного резерва» изменчивости (Шмальгаузен, 1941; Гершензон, 1941). Это явление означает, что канализирующие механизмы развития, забуферивающие индивидуальные генетические отличия особей, оказываются нарушенными при достижении некоторого порога внешних воздействий, в результате чего эти отличия проявляются в виде фенотипических аберраций. Все нормальные зиготы в популяции различаются как по ширине интервала условий, допускающих канализированное развитие, так и по характеру морфозов, осуществляемых ими в одних и тех же запредельных условиях (рис. 4, 5).
На основе представления об эпигенетическом ландшафте возможны и другие предсказания, доступные экспериментальной проверке. Если нарушение нормального развития органа А, обусловленное мутацией (или любым повреждающим воздействием), сводится к изменению меры какого-то фактора, дестабилизирующего траекторию А, то можно ожидать, что при искусственном переключении развития данного зачатка в направлении органа Б его новая траектория будет независима от указанного фактора, т. е. окажется «вне досягаемости» мутаций, воздействующих только на орган А. И наоборот, мутации, задевающие развитие органа Б, должны действовать и на любой другой зачаток, детерминированный в направлении Б, т. е. чувствительность зачатков к тем или иным нарушениям должна определяться не столько их нормальной детерминацией, сколько выбором фактического пути их развития. Эти закономерности действительно были установлены И. А. Рапопортом (1941), показавшим, что у дрозофилы эффект доминантной мутации Met, ведущей к поглощению презумптивного материала крыла мезотораксом, не подвергается каким-либо изменениям при введении в геном различных крыловых мутаций. С другой стороны, мутации, в норме влияющие на строение спинных щетинок, но не задевающие крыло, оказывают такое же действие и на гигантский мезоторакс, измененный мутацией Met.
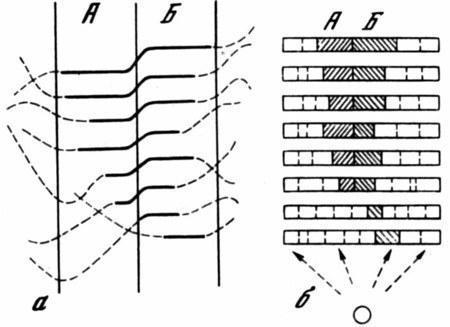
Рис. 4. Индивидуальные внутрипопуляционные различия в устойчивости нормального формообразования
а — кривые зависимости фенотипической реализации отдельных геномов от внешних условий сплошные участки соответствуют норме (с модификациями А и Б), пунктирные — неадаптивным морфозам; б — модификационные спектры, контролируемые этими геномами в одном и том же интервале условий, выходящем за пределы нормальных. Участки спектров, соответствующие модификациям А и Б заштрихованы
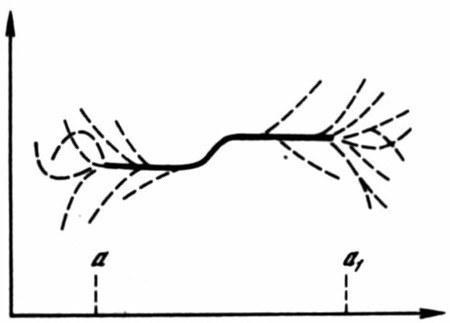
Рис. .5 Зависимость формообразования от условий развития
Сплошная линия соответствует общей для популяции адаптивной норме (с двумя модификациями); пунктиры соответствуют индивидуальным морфозам. По горизонтали — изменение среды, по вертикали — результат развития: а–а1 — интервал условий, допускающих нормальное развитие
Из рассмотренных представлений следует еще один вывод — что дискретность изменений фенотипа, вызываемая мутациями одного хромосомного локуса, выражает свойства целостной системы развития, а не непосредственно самих этих мутаций. Еще ранними работами Гольдшмидта (1916–1917 гг.) по механизму определения пола и С. Райта (Wright, 1916) по феногенезу мутаций альбинизма было показано, что в основе аллельных изменений лежат градации одного и того же фактора (например, количества вещества или скорости реакции), действующие с пороговым эффектом. С точки зрения модели эпигенетического ландшафта эта дискретность реагирования (убывающая в поздних стадиях развития) является неизбежным следствием того, что устойчивость канализированной траектории к колебаниям уровня тех или иных морфогенетических факторов снижается во времени прерывисто, образуя перепады в точках «чувствительных периодов», соответствующих ответвлениям аберративных долин (рис. 6, а). Чем больше уровень данного фактора выходит под действием мутаций за регулируемые пределы, тем более ранняя из этих критических фаз оказывается задетой, т. е. реализуется траектория, все более глубоко дивергентная по отношению к нормальной. Колебания фактора между двумя пороговыми уровнями (рис 6, б, уровни В и С), определяющими предельные возможности осуществления данного уклонения, не получают фенотипического выражения, и все вызывающие их однолокусные мутации будут расцениваться как один и тот же аллель. Выход фактора за один из порогов приведет к дискретному изменению фенотипа, т. е. переключению развития на более раннее или более позднее ответвление нормальной траектории.
Едва ли нужно пояснять, что концепция эпигенетической системы объективно лежит в основе теории стабилизирующего отбора. Представление о развитии как системе с ограниченным набором наиболее вероятных траекторий (фенотипических состояний) позволило разрушить непроходимую грань между мутационными и экзогенными изменениями, существовавшую для классической генетики (Dobzhansky, 1947, с. 209), сведя и те, и другие к дискретным реакциям целостной системы на количественные изменения переключающих эпигенетических факторов (Шмальгаузен, 1982, с. 82, 89, 103, 170–173). Установление этой закономерности открыло путь к пониманию того, что устойчивость наследования представляет собой не свойство отдельных хромосомных генов, а результат создания отбором новой организации генотипа, обеспечивающей канализированное развитие прежде лабильного (ненаследственного) признака. Наследственность превратилась тем самым в целостное и исторически обусловленное свойство, определяемое как устойчивость результата эпигенетических взаимодействий (Шмальгаузен, 1982; Waddington, 1957), а естественный отбор из сортировщика независимых от него наследственных единиц превратился в механизм создания наследуемых изменений.
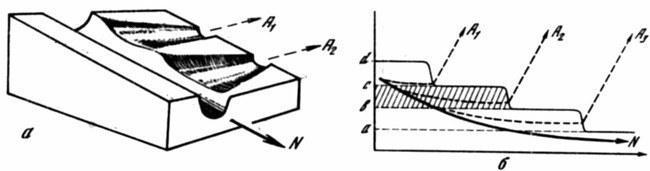
Рис. 6. Дискретность аберраций как выражение ступенчатого снижения помехоустойчивости канализированного развития
а — участок эпигенетического ландшафта, показывающий перепады высоты стенок (порогов устойчивости) креода в местах ответвления аберративных долин; б — зависимость характера аберрации от соотношения между уровнем повреждающего морфогенетического фактора и устойчивостью креода. В промежутке между двумя порогами устойчивости (заштрихован) колебания меры фактора не меняют типа развития; а, b, с, d — пороговые уровни устойчивости последовательных отрезков креода, A1–A3 — аберрантные пути развития, N — нормальный ход изменений уровня фактора и типичный путь развития. По вертикали — уровень фактора, по горизонтали — время
Представления Шмальгаузена (1968а, 1982) об историческом становлении новых адаптивных признаков легко интерпретируются как описание преобразований видоспецифичного эпигенетического ландшафта. Весь процесс, согласно этим взглядам, начинается каждый раз с лабилизации развития прежней нормы (рис. 7, а, б; N) и отбора одной из возникающих при этом элементарных неадаптивных реакций. Это означает, что в новых условиях индивидуальные варианты ландшафта реализуют различные аберрантные траектории, из которых одна ведет к наиболее жизнеспособному фенотипическому уклонению (рис. 7, б, N1). По мере отбора в его пользу прежняя нормальная траектория все более теряет устойчивость безотносительно к условиям развития, т. е. ее долина сглаживается, что означает стирание различий в стабильности между прежней нормой и отбираемым уклонением (рис. 7, б, N, N1). Постепенная селективная стабилизация новой адаптивной траектории превращает ее в креод (рис. 7, в, г, N1); вместе с этим вся зона ландшафта вокруг нее превращается из области маловероятных событий в область наиболее обычных уклонений, т. е. она расчленяется новыми долинами. Таким образом, постепенно изменяется весь рисунок ландшафта (рис. 7, а–г). Другими словами, происходит изменение аберративного пространства эпигенетической системы.
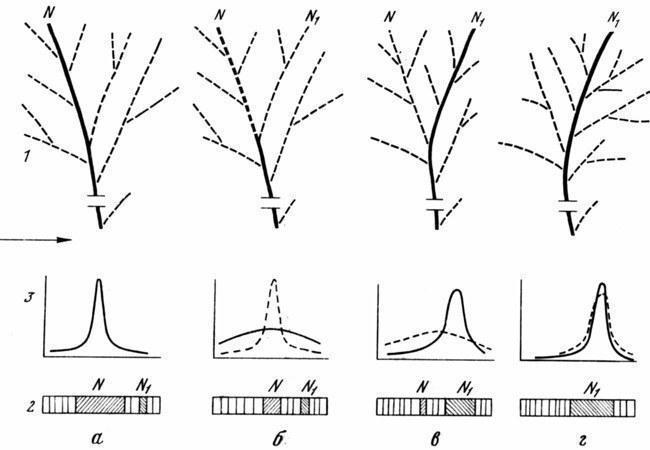
Рис. 7. Перестройка системы развития в ходе становления новой элементарной адаптации
1 — изменения эпигенетического ландшафта, 2 — типичные изменения модификационных спектров, 3 — изменения дисперсии преобразуемого адаптивного признака в популяции: а — канализированное развитие фенотипа N (соответствующего главной полосе в спектрах и пику вариационной кривой); б — дестабилизация развития, ведущая к снижению частоты фенотипа N и уменьшению его роли в спектрах; в, г — две последовательные стадии канализации онтогенетической траектории N1 с развитием вокруг нее новой сети аберративиых долин. Фенотип N1 из неустойчивого морфоза превращается в новую адаптивную норму; прежняя норма становится морфозом или исчезает. Жирными линиями выделены креоды; заштрихованы типы развития, соответствующие последовательным нормам. По вертикали — частота встречаемости признака, по горизонтали — изменение признака
Итак, на любом этапе своих эволюционных изменений система обнаруживает специфический набор возможностей развития, свойственный ей лишь в данный период ее истории. Поэтому утверждение, что материал эволюции составляют случайные геновариации, является, по выражению Уоддингтона, «пустым» (Waddington, 1957, р. 188). Сколько бы случайными ни были нуклеотидные изменения хромосомной ДНК, их возможные воздействия на фенотип всегда ограничены исторически сложившейся структурой (ландшафтом) эпигенетической системы. Изменение этой структуры выражается на популяционном уровне как изменение характера неопределенной изменчивости (Камшилов, 1967). Поэтому указание Дарвина (1952) о том, что изменчивость не вызывается отбором, должно быть правильно понято. Независимым от отбора является лишь само ее наличие, но не характер ее фенотипического выражения.
ЭПИГЕНЕТИЧЕСКАЯ СИСТЕМА И ПОНЯТИЕ МЕНДЕЛЕВСКОГО ФАКТОРА
Изложенные представления неизбежно подводят нас к проблеме онтогенетической интерпретации менделевского наследования. Понятие менделевского фактора означает наличие двух устойчивых альтернативных (аллельных) состояний признака, каждое из которых наследуется в соответствующей «чистой линии» однозначно, а у гибридов (второго поколения) — в определенных числовых соотношениях. С эпигенетической точки зрения это означает, что развитие признака у сравниваемых групп особей канализировано в двух различных направлениях, причем помехоустойчивость каждого из них основана на взаимодействии всех элементов генотипа и выражается в нечувствительности к эффекту рекомбинаций, возникающих при скрещивании внутри данной группы (линии). При межлинейном скрещивании оба направления (креода) объединяются в одном эпигенетическом ландшафте как стабилизированные разветвления одной траектории, и исход развития в принципе может колебаться между двумя крайними ситуациями. При одной из них реализуется та или иная промежуточная аберрантная траектория, что нередко дает не вполне устойчивый результат, который в практике менделевского анализа описывается как отсутствие единообразия гибридов первого поколения (Филипченко, 1924). Во втором случае (рис. 8) развитие направляется к одной из двух имеющихся областей притяжения (креодов), т. е. имеет место выраженное доминирование. При любом варианте выбор пути развития зависит от неспецифического сдвига в фазе неустойчивой детерминации (точке разветвления креодов), связанного в конечном счете с функцией той или иной пары гомологичных геномных единиц. Последняя отождествляется моргановской хромосомной генетикой с аллельными состояниями менделевского гена. Как уже отмечено, феногенетический анализ аллельных серий фенотипов показывает, что их дискретность отражает не свойства самого детерминирующего фактора, а пороговую реакцию системы развития на его количественные изменения. С этой точки зрения действие пары локусов, ответственных при данном состоянии системы за переключения пути развития, может контролировать в ней три градации фактора — две крайних (родительских) — АА и аа и одну промежуточную (гибридную) — Аа, обозначаемые генетикой соответственно как гомо- и гетерозиготное состояния. Между ними лежат либо два критических порога, либо в случае доминирования только один (между уровнями аа и Аа — рис. 8). При этом кажется очевидным, что для упорядоченного осуществления каждого из родительских фенотипов не требуется совершенно одинаковой интенсивности действия «изоаллельных» локусов-переключателей (или тем более идентичности их нуклеотидной структуры). Достаточно лишь, чтобы суммарный эффект любой индивидуальной их пары, взятой из линии аа, не превышал порога, допускающего развитие в направлении соответствующего фенотипа (рис. 8). Как в общем случае, так и при доминировании это — порог между градациями аа и Аа. При скрещивании гибридов первого поколения в соответствии с законами расхождения хромосом в мейозе возникают, как известно, зиготы с локусными комбинациями всех трех типов в количественных соотношениях 1:2:1, что означает реализацию в тех же пропорциях трех соответствующих мер переключающего фактора и определяемых ими вариантов развития, т. е. происходит менделевское расщепление по фенотипам. При наличии в системе лишь одного порога переключения (доминировании) возникают лишь два исходных стабилизированных варианта в соотношении 3:1.
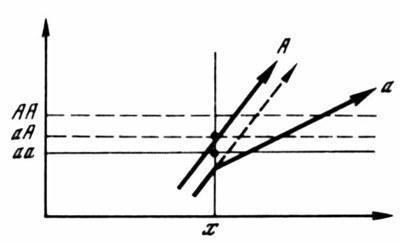
Рис. 8. Онтогенетический механизм менделевского наследования
Выбор пути развития у второго поколения гибридов стабилизированный линии с фенотипами А и а. Если к критическому моменту развития Х уровень переключающего морфогенного фактора не превышает порогового значения аа, то развитие отклоняется от креода А на путь, соответствующий рецессивному фенотипу а. В случае превышения этого порога развитие продолжается в направлении более устойчивого (доминантного) фенотипа А. По горизонтали — время, по вертикали — мера переключающего фактора и направление развития. Аа, АА — надпороговые значения фактора, реализуемые у гибридов.
В первой критической фазе два реализуемых варианта развития дают соотношение 3:1 в зависимости от того, превышается или нет пороговое значение (аа) фактора I. Во второй критической фазе, где выбор определяется уровнем фактора II (с пороговым значением вв), каждый из вариантов дифференцируется на два новых — также в соотношении 3:1. В итоге возникают четыре класса фенотипов в отношении 9:3:3:1. АА, Аа, аа, ВВ, Bв, вв — уровни переключающих факторов, определяемые рекомбинацией двух пар локусов; АВ.., Ав.., аВ.., aв.. — классы осуществляемых фенотипов
Любая упорядоченность биологических явлений должна рассматриваться как продукт естественного отбора (Майр, 1981), и в отношении правил менделевского наследования эта связь совершенно очевидна. Упорядоченность результата развития основана здесь на наличии в эпигенетической системе родителей соответствующих креодов, которые могут возникать лишь в результате стабилизирующего действия отбора. Геномные мутации, как и другие единичные воздействия на систему, могут, как показано, лишь нарушать креоды, но сами по себе не создают помехоустойчивых путей развития. В соответствии с этим выражение «сырых» мутационных аномалий, как природных, так и экспериментальных, весьма изменчиво и в целом плохо подчиняется менделевским правилам (Дубинин и др., 1937; Гершензон, 1941; Камшилов, 1940; Шмальгаузен, 19686), что и привело к возникновению понятий генотипической среды гена, а также экспрессивности и пенетрантности мутантного признака. Лишь эффекты наиболее крупных геномных нарушений показывают относительно малую зависимость от вариаций генотипа, т. е. такие мутации достаточно однозначно детерминируют определенный тип аномалии; но это соответствие все же никогда не бывает вполне стабильным [Последнее, в частности, касается крупных генетических аномалии у человека, которые, даже будучи доминантными, могут иногда не проявляться в гомозиготе и показывают в целом изменчивость выражения (например, полидактилия; Гершкович, 1968)].
Обусловленность правильного наследования предварительной стабилизацией скрещиваемых фенотипов очевидна уже из того факта, что законы Менделя были установлены именно на жестко отселектированных (стабилизированных) линиях носителей альтернативных признаков. О специфичности условий, при которых выполняются эти законы, особенно наглядно говорит содержание второго из них — закона независимой рекомбинации, указывающего, что при наследовании каждая пара аллельных факторов «ведет себя как единственная» (Филипченко, 1924), т. е. всегда дает во втором поколении гибридов расщепление на два родительских фенотипа на фоне любых вариантов рекомбинации всех остальных факторов. С точки зрения хромосомной генетики признание универсальности этого правила было бы равносильно неприемлемому для нее утверждению, что характер выражения гена не зависит от вариаций генотипической среды. Очевидно, что такая независимость возможна лишь при условии, что влияние рекомбинации на ход развития снивелировано предшествующим отбором, так что при любом ее варианте сохраняется лишь выбор из двух возможностей, определяемый в какой-то критический момент развития состоянием только одной пары локусов. В соответствии с этим действительно признается, что необходимой предпосылкой генетического анализа, опирающегося на менделевские правила наследования, является предварительный отбор линий по анализируемым признакам (Лобашев, 1966); и, в частности, с этого начинается анализ мутантных фенотипов. В представлениях генетики результатом такого отбора является создание чистой линии носителей мутантного аллеля, т. е. гомозигот по тому локусу, с которым отождествляется данный аномальный признак. С эпигенетической же точки зрения эта процедура означает стабилизацию прежде неустойчивой траектории развития признака, основанную на реорганизации всего генотипа данной линии (Waddington, 1957). Возникает вопрос: в чем же состоит «решающая» роль одной локусной пары в определении такого признака, выявляемая анализирующим скрещиванием? Прежде всего, она характеризует только его упорядоченное развитие, созданное селекцией, а не исходное состояние, при котором он был лишь неустойчивой аномалией. Во-вторых, эта определяющая роль относится не к однозначному канализированному развитию признака, наблюдаемому в родительских линиях, а к гибридному варианту, где в одной эпигенетической системе совмещены две разные возможности такого развития. Сущность онтогенетического действия «детерминирующей» локусной пары в системе гибрида заключается в том, что на временном отрезке, соответствующем выбору между двумя траекториями, ее эффект выходит за рамки регуляции одного из альтернативных креодов, но остается в пределах нормы для второго, так что все равно осуществляется один из стабилизированных вариантов развития. От переключателя здесь зависит лишь выбор пути развития, но не само его осуществление, определяемое всем генотипом. Что же касается гомозиготности родительской линии по одному из членов «детерминирующей» пары, то, как уже сказано, это понятие означает не отсутствие вариабельности локуса, а ограниченность ее теми пределами, при которых действие его двойной дозы не нарушает устойчивого развития признака, маркирующего данную линию (рис. 8). Подобные пределы вариабельности, ограничивающие возможности канализированного развития, существуют в данной системе для любой пары локусов, т. е. все они являются в этом смысле «детерминантами» признака.
Таким образом, понятие менделевского гена подразумевает определенное состояние видовой эпигенетической системы, при котором в ней благодаря отбору становятся возможными два устойчивых варианта развития признака, выбираемых в зависимости от сочетания состояний определенной пары хромосомных локусов. Поэтому законы их рекомбинации, определяемые поведением хромосом при мейозе и оплодотворении (расхождение и независимое комбинирование гомологов) получают при такой организации системы однозначное выражение в наследовании альтернативных признаков, и эта однозначность составляет сущность законов Менделя, т. е. менделевский ген — это показатель определенной упорядоченности эпигенетической системы, а не сам хромосомный локус. Последний получает свойства переключателя аллельных признаков лишь постольку, поскольку в системе развития гибридов существует сама возможность переключения, выражающаяся в наличии в ней двух стабилизированных креодов. Только в этих условиях приобретают смысл понятия гомо- и гетерозиготности, ибо они обозначают те градации эпигенетического эффекта локусной пары, которые в данной системе определяют выбор дискретных путей развития; положение же порогов между этими градациями, как и наличие самих включаемых ими путей, характеризуют систему, а не участок генома. При отсутствии в системе второго креода (т. е. при существовании у признака только одной нормы) любое геномное изменение (рекомбинация, мутация) может иметь лишь два эффекта — либо оно не задевает нормального развития, либо приводит к неустойчивым аберрациям, не показывающим правильного наследования (как это и наблюдается в отношении природной неопределенной изменчивости). Это свойство аберраций означает, что для однокреодной системы понятие аллеля вообще теряет содержание, так как в ней просто нет альтернативной возможности устойчивого развития, поддающейся включению при каком-либо локусном изменении.
Из сказанного следует, что менделевский фактор — это не материальная частица, а отношение между двумя устойчивыми альтернативными состояниями эпигенетической системы («чистыми линиями»), выявляемое в гибридном анализе. Это отношение не существует вне сравнения указанных состояний. Число менделевских генов, которым определяется анализируемый признак в данной системе скрещиваний, означает не что иное, как число двоичных выборов между последовательными ветвлениями канализированных траекторий, которое должно быть сделано в системе развития гибрида, чтобы получить в итоге один из родительских фенотипов (рис. 9). Легко видеть, что если каждый такой выбор зависит в данной системе от одной пары локусов, дающей соотношение 3:1 при реализации двух соответствующих траекторий (креодов), то итоговое число возможных результатов развития (т. е. различающихся фенотипов) будет составлять 2n, а их количественное соотношение (3+1)n, где п — число последовательных критических фаз, соответствующих развилкам между дочерними креодами, или, что то же самое, число «детерминирующих генов» (рис. 9). Эти соотношения и составляют суть менделевского наследования.
Рис. 9. Дигибридное расщепление как результат двух последовательных выборов между дихотомирующими креодами в эпигенетической системе гибридов второго поколения
Все это делает очевидным, что понятие менделевского аллеля и геномной мутации обозначают совершенно различные явления, относящиеся к разным иерархическим уровням системы. Менделевский ген с его аллельными состояниями характеризует такой способ организации системы, при котором широкий спектр вариаций генома как целого не выводит развитие за пределы ограниченного набора дискретных устойчивых путей; выбор среди них зависит каждый раз лишь от суммарного уровня активности одной определенной пары гомологических локусов. Напротив, геномная мутация означает элементарное возмущение в системе, которое может лишь нарушить ее упорядоченность, но не в состоянии создать новой. Отождествлять менделевский ген с участком хромосомы, а аллельное изменение с мутацией этого участка — значит наделять элемент системы свойствами самой системы.
Это отождествление, принятое, хотя и не без колебаний, моргановской хромосомной генетикой (Морган, 1937), заставляет нас вернуться к вопросу о возможности изменения гена под действием отбора. Такая возможность, как будто бы вытекавшая из ряда опытов (Castle, 1916), энергично отвергалась ортодоксальным моргановским учением, видевшем в гене локус, способный лишь спонтанно мутировать. Но даже если иметь в виду собственно локус, то в сегодняшних представлениях — это полинуклеотидная матрица, чувствительность которой к отбору признается в известной степени даже нейтралистской теорией молекулярной эволюции. Тем более нет сомнений в отношении селективной обусловленности менделевских факторов (которые и выявляются методами классической генетики), ибо, как показано выше, они представляют собой просто символы эпигенетических соотношений, созданных отбором. Проще говоря, менделевский ген есть продукт отбора (Шишкин, 1984в).
Наиболее ясным экспериментальным доказательством этого являются опыты К. Уоддингтона по генетической ассимиляции (стабилизации) лабильных признаков (Waddington, 1957). Здесь отбирались крыловые морфозы у дрозофилы, такие, как dumpy или bithorax, полученные путем теплового воздействия на личинку, после чего потомство их носителей снова подвергалось тепловому шоку на той же стадии и отбору по той же дефинитивной аномалии. После нескольких поколений отбора указанные фенотипы стали появляться без действия шока, и из них были получены чистые линии. Таким образом, признаки стали устойчивыми (наследственными). При скрещивании их носителей с исходной нормальной культурой они выщеплялись с той или иной степенью пенетрантности (высокой у bithorax), что позволило рассматривать их как проявления соответствующих мутантных аллелей. Поскольку в исходной линии этих фенотипов заведомо не было, то возникает вопрос: откуда появился новый ген? Возможности ответа с традиционных позиций здесь весьма ограниченны. Во-первых, допускается, что полученные стойкие фенотипы суть эффекты скрытых мутаций, лежащих при обычных условиях ниже порога проявления; отбор же объединил их в полиген с более сильным эффектом. Это наиболее обычное объяснение (Шеппард, 1970; Майр, 1974; Рьюз, 1977), вынужденное апеллировать к допущениям, лежащим за пределами анализа; однако и оно признает, что детерминирующий комплексный фактор есть продукт отбора. Другая возможность — это допустить существование гипотетического эффекта Болдуина, т. е. появление в ходе каждого эксперимента мутаций, совпадающих по выражению с исходным морфозом; но тогда сама неизменность их появления в процессе отбора вырастает в очередную загадку [Интерпретация этих опытов самим Уоддингтоном вскрывает дуализм его представлений, отражающих стремление совместить эпигенетическое объяснение устойчивости фенотипа с идеей дискретной наследственности. Как эмбриолог-феногенетик Уоддингтон сознает, что получение наследственного признака, вызываемое генетической ассимиляцией, означает канализацию его развития, обусловленную всем генотипом и не зависящую от определенного локуса, т. е. поиски последнего заведомо неправомерны. Но, как генетик, он тем не менее задается вопросом: откуда возник ген, определяющий включение ассимилированного признака? При этом понятие переключателя креодов у гибридов бессознательно подменяется понятием гена, детерминирующего признак в ассимилированной линии. В рассматриваемых случаях (фенотипы dumpy и bithorax) Уоддингтон, естественно, не находит удовлетворительного «генетического» ответа и вынужден допускать здесь появление случайных мутаций в направлении отбора (эффект Болдуина) вопреки собственному определению механизма генетической ассимиляции (Waddington, 1957, с. 166, 176, 180–182)].
Напротив, эпигенетическое объяснение эксперимента не встречает никаких трудностей, так как результаты здесь прямо соответствуют ожиданиям. Менделевский ген в общем случае обнаруживается лишь там, где есть два устойчивых альтернативных состояния признака; эти два состояния означают наличие в системе развития гибридов двух креодов; креоды же создаются стабилизирующим отбором. Стабилизация первично неустойчивого морфоза в рассмотренных опытах привела к созданию линии, в которой путь его развития приобрел свойства креода. Совмещение последнего в одной системе с прежним креодом, характеризующим нормальную линию (гибридный анализ), привело к осуществлению двух альтернативных вариантов признака, т. е. к менделевскому расщеплению. Отсутствие полного проявления ассимилированного признака указывает лишь на недостаточную стабилизированность его траектории.
Таким образом, возможность строго упорядоченных эпигенетических переключении, составляющая сущность менделевского гена, возникла в результате отбора. Речь здесь идет в первую очередь не о создании отбором нового состояния локуса, а о приобретении этим локусом свойств переключателя.
Из всего сказанного легко понять, какой смысл должен вкладываться концепцией эпигенетической системы в процедуру менделевского анализа. Последний раскрывает нам не «генетическую основу» исследуемых признаков, а структуру целостной видоспецифичной системы развития, которую мы «проявляем» шаг за шагом с помощью стабилизации различных альтернативных траекторий, иерархически строящих эту систему и характеризующих ее потенциальные возможности. В зависимости от условий отбора одни и те же признаки могут быть стабилизированы различными путями, в результате чего выбор между их траекториями у гибридов будет зависеть каждый раз от разных локусов (что воспринимается исследователями как обусловленность данного признака в этих случаях неидентичными генами). Другими словами, подлинными инвариантами, определяющими выбор в признаковом пространстве данной системы, являются не состояния хромосомных локусов, а иерархические последовательности развилок (чувствительных точек) онтогенетических траекторий, упорядоченное переключение которых можно организовать с помощью отбора разными способами. Совокупность этих разветвлений и составляет ландшафт системы.
Инвариантность общего рисунка траекторий системы по отношению к их потенциально возможным «пусковым факторам» особенно наглядно выявляется в экспериментах по детерминации пола. У большинства ксеногамных организмов альтернативные комбинации половых признаков имеют простой хромосомный механизм переключения, т. е. наследуются как гетерозигота и рецессивная гомозигота по одному менделевскому фактору. При этом определение пола (например, у рыб и амфибий) в принципе можно полностью извратить с помощью гормональных воздействий или межрасового скрещивания таким образом, что зигота с генетической конституцией самца развивается в фенотипическую самку с нормальной репродуктивной способностью и наоборот; иногда удается с помощью отбора перенести механизм определения пола с половых хромосом на аутосомы; наконец, в ряде случаев у разных природных рас одного вида детерминация пола может осуществляться на основе противоположных вариантов гетерогаметности, т. е. мужского — XY и женского — WZ (Астауров, 1966). Все это ясно показывает, что способность гомологичных геномных элементов переключать развитие есть выражение структуры самой системы, допускающей такие переключения, и что в роли диспетчеров эти гомологи могут меняться ролями, а также могут быть заменены множеством других агентов с тем же неспецифическим эффектом, определяющим выбор одной из наличных канализированных возможностей. Подлинная причина, определяющая возможности выбора в такой системе,— это организация соответствующего ей целостного генотипа.
Рассмотренные взгляды на соотношение хромосомных единиц, менделевских генов и признаков фенотипа находятся не в столь уж резком несоответствии с теоретическими представлениями хромосомной генетики, как это может показаться. Несоответствия следует скорее искать внутри самих этих представлений, которые объединяют редукционистское истолкование числовых соотношений наследования в группах особей («признаки определяются дискретными генами») и вполне системное понимание роли геномных единиц в определении итога развития отдельной особи («каждый признак определяется всем геномом»). Основой первого из этих положений считается менделевский анализ; основу второго составляют данные экспериментальной генетики, показывающие, что один и тот же признак может нарушаться множеством разнородных мутаций и, следовательно, его нормальное развитие зависит от функции множества локусов (Морган, 1924, 1937а, б, в; Уоддингтон, 1947). Это последнее представление получило свой законченный вид в теории генного баланса (Bridges, 1922; Морган, 1937д), согласно которой каждый признак есть выражение равновесия, создаваемого в развитии действием всех генов, а изменение в любом из них (мутация) приводит к новому равновесию, дающему иной конечный продукт. Поэтому ген не создает признака, а лишь склоняет общий баланс взаимодействия к тому или иному фенотипическому исходу. Таким образом, теория признает, что конечный эффект отдельной хромосомной единицы выражает не ее собственные свойства, а свойства всей системы взаимодействий, в рамках которой данная единица функционирует. Все это по существу очень близко к признанию идеи о неспецифическом влиянии гена на итог развития, сформулированной позднее Гольдшмидтом.
Идея генного баланса составляет основу представлений хромосомной генетики об онтогенетическом («физиологическом») действии гена (Морган, 1937д; Лобашев, 1963; Майр, 1968, 1974; Дубинин, 1976; и др.). Соответственно представления об однозначной связи между генами и признаками формально отвергаются генетической теорией; их характеризуют как ошибку Вейсмана (Дубинин, 1966а, с. 238; Морган, 1937а, б, в), или как «генетику горохового мешка», отражающую позиции раннего менделизма (Майр, 1968, с. 216), или, наконец, как некомпетентное мнение, ошибочно приписываемое генетике (Тимофеев-Ресовский, Иванов, 1966, с. 116). Роль отдельных генов сводится лишь к смещению путей развития (Мюнтцинг, 1967, с. 73).
Однако принятие такой позиции обязывает ответить на вопрос: как в таком случае должны интерпретироваться результаты менделевского анализа, составляющего основой познавательный инструмент классической генетики? Последний, как известно, построен именно на однозначном проецировании генов на признаки (Шмальгаузен, 1982; Столетов, 1967; Майр, 1968), и это позволяет понять, почему принцип определения признака всем генотипом иногда называют чисто теоретической декларацией генетики, не меняющей ее редукционистских представлений (Светлов, 1964). В самом деле, можно ли согласовать оба подхода? Если, например, различия двух константных (чистолинейных) фенотипов сводятся по результатам анализа к различию в одном гене, понимаемом как хромосомный локус, то перевести этот вывод на язык балансовой гипотезы можно только одним способом — признать, что геномы всех особей обеих линий абсолютно идентичны, за исключением единственного участка, который сдвигает генное равновесие к одному из двух вариантов. Но это предположение неприемлемо, так как в действительности геномы любой лабораторной линии или природной расы всегда вариабельны по многим локусам (ср.: Камшилов, 1939; Дубинин, 1966а). На практике этих противоречий обычно даже не замечают, т. е. в истолковании менделевского анализа не усматривают особой проблемы. Указывается, правда, что его конкретный результат не следует понимать как выявление всей генетической основы нормального признака, которая всегда очень сложна. Анализ каждого простого расщепления вскрывает лишь отдельный ее элемент; полное же представление о ней дает исследование всех возможных мутационных аномалий признака. Но очевидно, что такое толкование несовместимо с балансовой гипотезой, поскольку в ее представлениях любой признак особи имеет всегда одну и ту же генетическую основу, а именно весь геном. Таким образом, очевидно два вывода. 1. Хромосомная генетика не содержит объяснения менделевского анализа, согласуемого с балансовой теорией. 2. Сведение монофакторного различия между двумя чистыми линиями к изменению в одном хромосомном локусе не отражает фактических различий их генотипов.
Итак, вопрос состоит в том, можно ли вообще избежать совмещения в генетической теории двух взаимоисключающих принципов: «ген (группа генов) — признак» и «геном (или генотип) — признак»? Для этого логически есть только одна возможность: признать, что ген балансовой гипотезы (хромосомный локус) и ген менделевского анализа — это не одно и то же. Другими словами, менделевский фактор должен выражать такое различие между двумя вариантами генотипа, которое принципиально несводимо к свойствам или состояниям их отдельных элементов. Именно к этому выводу мы и пришли выше, опираясь на теорию эпигенетической системы и отвечающую ей реальную феноменологию наследования. Правильное менделевское расщепление выражает взаимоотношение двух состояний системы, соответствующих двум альтернативам канализированного развития. Выбор между ними определяется у гибридов количественным уровнем активности одной гомологичной пары локусов, которая играет в этом случае роль «дифференциатора». Эта роль, выражаемая символом менделевского гена, обеспечивается исключительно свойством гибридных генотипов осуществлять два устойчивых варианта развития; при потере этого свойства исчезает и сама упорядоченность наследования, позволяющая выделять его контролирующий фактор.
Это представление о сущности гена в основе своей не является для генетики новым. Примерно таким же оно было и у автора термина «ген» Иогансена (Johannsen, 1926), который понимал под ним не частицу наследственного вещества, а единицу различия двух генотипов (Baur, 1922). Достижения хромосомной теории наследственности в глазах многих исследователей, особенно феногенетиков, не отменяли основы такого представления о гене. Последний, даже будучи отнесен к хромосоме, мыслился ими как причина различия в развитии, не существующая вне свойств целостного генотипа (Промптов, 1934; Камшилов, 1934); допущение о самостоятельном действии гена приравнивалось к возможности «изолированного существования дырки от бублика» (Промптов, 1934). Согласно Камшилову (1935, с. 141), ген — это не локус, а отличие двух целостных генотипов, «выявляемое в своеобразном типе развития». Даже для Моргана признание хромосомной локализации гена было сопряжено с некоторыми колебаниями, и одно время им допускалось, что речь идет лишь об абстрактном свойстве, каким-то образом связанном с данным участком хромосомы (Морган, 1937г). В наше время с развитием биохимических методов генетического анализа, где локус выступает как единица матричного синтеза, его нетождественность менделевскому гену вновь становится для генетиков все более очевидной (Грант, 1980, с. 290; Голубовский, 1982).
Признание системной обусловленности «генных» свойств локуса отчасти нашло выражение и в гипотезе Фишера (Fisher, 1930) об эволюции доминантности. Согласно последней, способность локуса детерминировать признак в присутствии своего неидентичного по свойствам гомолога (т. е. в гетерозиготе) есть результат его взаимодействия с системой модификаторов, созданной отбором. Тем самым признается, что по крайней мере классическая картина менделевского расщепления по фенотипам (3:1) не является свойством гомологичных хромосомных единиц и возникает лишь как продукт реорганизации отбором всего генотипа. Отсюда остается лишь шаг до распространения этого вывода на любое упорядоченное соответствие между локусом и признаком, выявляемое гибридным анализом.
С другой стороны, первый шаг к осознанию того факта, что в основе выбора между аллельными состояниями лежат какие-то количественные различия, был сделан еще ранними менделистами, получив выражение в известной гипотезе «присутствия—отсутствия» Бэтсона—Пеннета, выдвинутой для объяснения доминирования (доминантный признак определяется наличием фактора в гаметах, рецессивный — его отсутствием). Критика со стороны моргановской школы, указавшей на несовместимость этого взгляда с явлением множественного аллелизма и представлением о линейном расположении генов в хромосомах, привела к новой формулировке гипотезы, в которой еще более определенно говорилось о количественной природе аллельного различия (рецессивный фактор есть нечто утративший по сравнению с доминантным; Пеннет, 1930). И наконец, Гольдшмидт, независимо обосновавший идею о «количестве гена» как основе аллельных изменений, развил ее в представлении о пороговых уровнях морфогенетического эффекта, определяющих выбор фенотипа в критических точках развития (Goldschmidt, 1927, 1938, 1940).
Как показано выше, наличие у хромосомной единицы свойств менделевского фактора означает, что данная структура генотипа допускает несколько устойчивых путей развития признака. При отсутствии такого выбора, т. е. возможности переключения стабилизированных траекторий, не может быть и локуса-переключателя, т. е. гена, «определяющего» признак. Следовательно, в общем случае роль отдельного локуса в детерминации свойств фенотипа остается неопределенной и может быть описана лишь как элемент в сложной системе функционирования всего генотипа в процессе развития.
Но если отдельный локус не имеет самостоятельного выражения в фенотипе и последний в каждом своем признаке определяется всей зародышевой плазмой (генотипом), то в чем тогда заключается принцип корпускулярной (дискретной) наследственности, лежащей в основе хромосомной теории? Актуальность этого вопроса стала очевидной для Моргана еще в 1918 г., до появления балансовой гипотезы, и его ответ заслуживает внимания. Он заключает, что если указанные соотношения между признаками и зародышевой плазмой действительно имеют место, то последняя все же в любом случае остается построенной из элементов, независимых в отношении мутирования, кроссинговера, а также расхождения гомологов и комбинирования их пар в процессе созревания половых клеток (Морган, 1924, с. 232, 235). «В этом, и только в этом смысле мы вправе говорить о корпускулярном строении плазмы и корпускулярной наследственности» (Там же, с. 235). Эти признания поразительны. Из них следует, что моргановская теория наследственности описывает лишь закономерности перераспределения хромосомных элементов, но при этом не знает, каким образом они могут быть связаны с наследованием признаков! И тем самым вновь косвенно признается, что менделевские правила наследования признаков описывают нечто совсем иное, нежели элементарные свойства хромосомных единиц.
Отсутствие метода для интерпретации итога развития в терминах дискретных хромосомных генов, неявно признанное Морганом, не является следствием неполноты знаний, как часто полагают. Оно носит принципиальный характер. Данные эмбриологии не оставляют сомнений, что развитие есть неразложимый эпигенетический процесс, основанный на взаимодействии всех его элементарных факторов и непрерывном увеличении качественного многообразия. Явления цитоплазматической прелокализации зачатков и связанных с нею фаз преформированного (мозаичного) развития составляют здесь лишь частные эпизоды, обусловленные эпигенетическими взаимодействиями в оогенезе и вновь сменяемые в ходе развития регуляционными процессами (Шпеманн, 1925; Дэвидсон, 1972; Светлов, 1978). Детерминация частей определяется в развитии лишь детерминацией целого, и признаки взрослого организма не могут иметь коррелятов в зиготе или ее геноме (Гурвич, 1944; Светлов, 1964, 1978). Как мы видели, любой итог развития — нормальный или аберрантный — всегда является эквифинальным по отношению к вариациям значений его элементарных причинных факторов. Устойчивость признаков «не свойство генов, а выражение взаимозависимостей частей в корреляционных системах развивающегося организма» (Шмальгауэен, 1982, с. 174). Эта несводимость процесса развития к преформационной модели, рассматривающей организм как сумму следствий из независимых начальных причин, давно понятая экспериментальной эмбриологией (Бляхер и др., 1935; Белоусов, 1979), формально признается также и хромосомной генетикой, видящей в этом одно из главных своих отличий от вейсмановской теории зародышевой плазмы (Морган, 1937а, б; Дубинин, 1966а).
Как уже говорилось, отсутствие специфической роли индивидуальных хромосомных генов (локусов) в определении свойств фенотипа, наиболее аргументировано было показано Гольдшмидтом, и он оказался, одним из тех немногих ведущих генетиков, кто ясно увидел, к каким последствиям это ведет для теории развития (а тем самым и наследственности). «Факты генетики, конечно, могут описываться в терминах генов, но теория зародышевой плазмы должна полностью освободиться от концепции генов как единиц» (Goldschmidt, 1938, р. 311), т. е. генотип является неразложимой основой развития фенотипа. «Зародышевая плазма как целое контролирует определенную реактивную систему, которая есть не мозаика отдельных эффектов, но единая система развития, управляемая как целое одним фактором... Для многих генетиков явно трудно мыслить в таких понятиях, поскольку большинство их настолько связаны аксиоматической верой в атомистическую генную теорию, что не в состоянии думать иначе; но эмбриологи, физиологи и, возможно, систематики не найдут трудностей в принятии этой концепции» (Goldschmidt, 1940, р. 218).
Предвидения Гольдшмидта в отношении принятия генетикой этих взглядов (к которым она, казалось бы, подошла вплотную) вполне оправдались, и причины этого понятны. Системные обобщения генетики, в которых не остается места для генов как детерминантов признаков, целиком относятся к ее теоретическим представлениям об индивидуальном развитии, т. е. той области, где она не имеет особенно больших успехов. Напротив, редукционистский подход к изучению наследственности, связанный с применением менделевского анализа, является основой всех тех огромных и очевидных практических достижений, которые позволили генетике претендовать на роль точной науки и завоевали ей небывалый авторитет в биологии XX в. Не удивительно, что в этих условиях, когда главной повседневной задачей остается установление связей между геномными единицами и признаками организма, кажется неуместным вспоминать о том, что в теории за генами не признается таких детерминирующих свойств и что их следует относить только к генотипу. Сама очевидность выявляемых дискретных связей и возможность их экспериментальной проверки обычно мешают задуматься над тем, какова в действительности их природа и при каких условиях они возникают. Свойство локуса переключать пути развития при скрещивании двух стабилизированных отбором фенотипов абсолютизируется как его постоянное свойство, а сама его функция переключателя, выявляемая лишь у гибридов, рассматривается как доказательство его особой детерминирующей роли в отношении данного признака. Одним из красноречивых свидетельств торжества этого редукционистского стиля мышления является хрестоматийное утверждение, что принцип «чистоты гамет» сам по себе обеспечивает дискретность наследования (несмешиваемость) признаков, т. е. что законы расхождения хромосомных гомологов должны всегда прямо отражаться в свойствах фенотипов, независимо от устойчивости их развития.
Необходимость увязать эти представления с системными требованиями балансовой гипотезы на практике выливается в «компромиссное» решение, т. е. концепцию генотипической среды. Участие всего генотипа в реализации признака понимается здесь как взаимодействие «основного» гена (или генов), определяющего признак, с остальными наследственными элементами, модифицирующими его специфическое действие и играющими по отношению к нему роль «шумов». Такое понимание соотношений гена и признака утвердилось в умах многих исследователей как типичное для хромосомной генетики независимо от того, отвергают ли они его (Goldschmidt, 1940) или считают приемлемым (например, Астауров, 1971, с. 218; Bertalanffy, 1969, р. 73). Вполне очевидно, что эта интерпретация целиком остается в рамках преформистских представлений о дискретной (мозаичной) детерминации признаков. По существу она мало отличается от концепции Вейсмана, в которой возможность проявления каждого детерминанта подобным же образом зависела от общего исхода борьбы наследственных зачатков. Эта, казалось бы, постоянно отвергаемая, но на деле прочно укоренившаяся вера в независимое определение признаков хромосомными генами косвенно обнаруживается даже там, где на словах подчеркивается существование у видового генотипа и контролируемой им эпигенетической системы особых целостных свойств, не нарушаемых элементарными мутациями. Ибо, когда речь заходит о сравнении таких генотипов, то их различия на самом деле оцениваются не в целостных свойствах, а лишь в количестве неидентичных генов (Dobzhansky, 1947, с. 106, 110, 337, 338; Майр, 1968, с. 432). Сущность генотипа как целого, выражаемая только через индивидуальное развитие (Камшилов, 1934), по-прежнему сводится здесь к сумме хромосомных элементов.
* * *
В историческом плане появление каждой новой области естествознания обычно начинается с ощупывания предмета исследования аналитическими методами и, как следствие этого, с преобладания редукционистского подхода к его описанию; время системных обобщений приходит позднее. Так было и в истории традиционных экспериментальных областей биологии — механики развития и генетики, из которых последняя прошла путь от типичной «механики наследственности» (Вейсман и ранние менделисты) до системных построений Гольдшмидта, Шмальгаузена и Уоддингтона. Естественно поэтому, что появление и бурное развитие нового фронта исследований — молекулярной генетики, изучающей строение и функции матричных структур клетки, снова привели к возрождению редукционистских представлений о связи гена и признака, выраженных теперь в формуле «один ген — один фермент». Это внесло новый элемент дуализма в мировоззрение классической генетики, многие представители которой, отрицая в теории прямую детерминацию генами признаков фенотипа, в то же время вынуждены допускать ее возможность в отношении первичных генных продуктов (Тимофеев-Ресовский, Иванов, 1966; Waddington, 1966); последние (прежде всего белки) рассматриваются как признаки, наиболее адекватно отражающие свою генетическую основу (Гершензон, 1974). Все это не могло способствовать укреплению системных взглядов в классической генетике, влия ние которых и без того было ограниченным. Однако и в молекулярной биологии тем временем начинают слышаться новые ноты. Давно высказанное мнение, что принцип «ген—фермент» представляет крайнее упрощение реальных соотношений (Haldane, 1954), находит теперь все больше подтверждений. Явления полиморфизма ферментов и сходство пространственной структуры их гомологов у разных организмов уже сами по себе заставляют думать, что именно эта структура играет главную роль в определении функциональных свойств белка, тогда как образующие его аминокислотные последовательности (во всяком случае в участках, лежащих вне активных центров) могут при этом варьировать. Таким образом, имеется вырожденность соответствия не только между нуклеотидным кодом ДНК и линейной структурой белка, но также и между последней и характером его функции (Уоддингтон, 1970б; Волькенштейн, 1981а, б). Более специальные исследования показывают, что весь многоступенчатый процесс, ведущий от цистрона к ферменту, т. е. транскрипция—процессинг (посттранскрипционная редукция иРНК) — трансляция и посттрансляционные изменения — является поливариантным на каждом своем этапе (Инге-Вечтомов, 1976; Михайлова, Симаров и др., 1981). Одним из ярких проявлений этой поливариантности является тот факт, что в норме с одного и того же сегмента ДНК могут считываться в разных тканях или на разных стадиях онтогенеза различающиеся транскрипты и соответственно образуются разные белки (Голубовский, 1985). В целом такая неоднозначность может быть обусловлена воздействиями самой различной природы, что создает, с одной стороны, возможность уклонения конечного продукта синтеза от нормы, а с другой — создает основу для его регуляции («фенотипической супрессии» мутаций) при ошибках кода. Такая регуляция, в частности, показана у дрожжей-сахаромицетов для посттранскрипционных этапов синтеза, где ее можно вызвать изменением температуры, рН, осмотического давления и действием других внешних агентов (стрептомицина, глицерина и т. д.). Эти процессы предотвращают потери в структуре полипептидной цепи и облегчают ее функциональную регуляцию на посттрансляционном уровне (Михайлова, Симаров и др., 1981). Последняя может экспериментально индуцироваться теми же агентами, что и посттранскрипционная регуляция, но во многих случаях она наблюдается как естественный процесс, приводящий к сохранению мутантом нормальной биохимической функции. При этом часто имеет место не исправление ошибок в аминокислотных последовательностях, а нивелирование их эффекта. Наиболее известный механизм этой регуляции, изучавшийся у различных организмов — взаимодействие продукта активности мутантного локуса с нормальными или мутантными субъединицами того же белка («межаллельная комплементация»). Оно приводит либо к восполнению утрат в первичной структуре одной полипептидной цепи за счет других, либо к восстановлению функции целого за счет конформационных изменений мутантных субъединиц, т. е. такому объединению имеющихся фрагментов, которое позволяет их комплексу приобрести нормальную пространственную структуру путем самосборки. При этом аминокислотные изменения вне активных центров не нарушают нормальной функции (Инге-Вечтомов, Сойдла, 1978).
Явления комплементации продуктов синтеза, нарушающие колинеарное соответствие между цистроном и ферментом, проливают свет на эволюционное значение посттрансляционных изменений вообще. Тот факт, что активные центры белков сохраняют относительную автономность и нередко могут функционировать независимо от их связи (например, две неидентичные субъединицы триптофансинтетазы, разделенные у кишечной палочки, но кодируемые как одно целое у нейроспоры), заставляет исследователей полагать, что многие мультифункциональные белки могли возникать путем объединения ферментов, кодируемых разными генами, и, наоборот, что мультиферментные комплексы в ряде случаев являются продуктами эпигенетического расчленения мультифункциональных белков-предшественников, как это прямо наблюдается у некоторых вирусов (Инге-Вечтомов, Сойдла, 1978). Здесь уже не приходится говорить о правиле «ген—фермент». Необходимость учитывать возможный эффект посттрансляционных модификаций становиться все более очевидной для исследователей, заставляя воздерживаться от прямого отождествления рядов изоферментов с продуктами изоаллельных мутаций (Солбриг, Солбриг, 1982, с. 256).
Все эти факты показывают, что, несмотря на большое различие в дистанции, разделя ющей локус и фенотип в классической и молекулярной генетике, их взаимоотношения в принципе сходны. Функциональную устойчивость конечного продукта синтеза невозможно свести здесь к устойчивости самих матриц (ДНК и РНК). Она основывается на регулирующих эпигенетических взаимодействиях, охватывающих всю систему синтеза и способных забуферивать определенные ошибки генетического кода, а также нарушения процессов транскрипции и трансляции. Как и в «макроонтогенезе», регуляционные возможности этой системы чувствительны к внешним факторам. Синтезированная молекула в такой же мере является продуктом цистрона, как и всех не зависящих от него элементов системы синтеза (ферментов, РНК и т. д.), и не обязательно колинеарна цистрону. Все это заставляет полагать, что и эволюционный механизм становления белков в принципе является тем же, что и у других элементов адаптивной нормы. Белковая молекула также имеет «фенотип», частично определяемый средой, и его соответствие субстрату своего действия должно быть исторически стабилизировано (Уоддингтон, 1970б). Очевидно, что первым шагом эволюционного становления нового типа белка должно быть его возникновение в качестве одной из неустойчивых посттрансляционных модификаций существующего фермента. Если последняя оказалась в новых условиях адаптивно ценной, то путь ее осуществления преобразуется отбором в сторону максимальной помехоустойчивости. Очевидно, это выражается прежде всего в постепенном спрямлении и упрощении всей последовательности этапов синтеза, т. е. установлении все большей колинеарности между новой молекулой и исходной для нее матрицей. С этой точки зрения случаи упорядоченных посттрансляционных превращений ряда нормальных белков можно истолковать как промежуточные этапы стабилизации их морфогенеза, «рекапитулирующие» ход первично неустойчивых преобразований. К таким примерам относится, в частности, образование у млекопитающих инсулина путем протеолиза гигантской молекулы-предшественника и разделения ее на две субъединицы или же образование основного белка у фага Т4, где около 900 одинаковых полипептидных цепей сначала спонтанно агрегируют, а затем отщепляют N-концевые участки (Стент, Кэлиндар, 1981). Вероятно, также и вырезание неактивных (интронных) последовательностей из матричной РНК, составляющее обычный этап белкового синтеза, представляет собой отражение преобразований, происходивших некогда на посттрансляционном уровне. Все это заставляет считать, что матрица ДНК, лежащая в основании стабильно синтезируемого белка, исторически является не причиной его возникновения, а, наоборот, следствием стабилизации его морфогенеза, первично основанного на модификации продукта, кодировавшегося другим вариантом матрицы. Если это верно, то мы должны заключить, что связывать историческое появление новых гомологичных белковых субъединиц, например b-гемоглобина, с дупликацией соответствующего гена (как это обыкновенно делают) — значит менять местами причины и следствия; онтогенетическая и историческая причинность не тождественны. К отчасти сходным выводам пришел Ю. М. Оленев (1977), указавший, что необходимость в новом продукте возникает раньше, чем удваивается соответствующий ген, и что эта проблема в эволюции решается сначала обходными путями. Возможность того, что расширение функций гена может предшествовать его дупликации, признает также Т. Р. Сойдла (1983).
Таким образом, механизм матричного синтеза может быть описан в своей основе как система, регулируемая к определенному конечному состоянию и, очевидно, преобразуемая в эволюции «сверху вниз» (от конечного продукта к матрице), т. е. в соответствии с теорией стабилизирующего отбора. Можно согласиться с Б. М. Медниковым (устное сообщение), считающим, что белок есть в сущности орган, к развитию которого приложимы все закономерности, справедливые для обычных органов.
Следует добавить еще одно соображение. Эффект биохимических мутаций может фенокопироваться (Goldschmidt, 1955), и если это правило так же универсально, как в отношении обычных фенотипов, то надо заключить, что любой продукт синтеза, получаемый на основе единичной мутации данного генотипа, может быть в принципе получен на той же основе и без мутации, с помощью подбора внешних воздействий, определенным образом нарушающих транскрипцию, трансляцию или поздний эпигенез. Обычно подобные случаи индуцированного синтеза объясняют дерепрессией оперонного механизма, содержащего необходимую матрицу, в соответствии со схемой Жакоба и Моно (Уоддинг тон, 1964; Волькенштейн, 19816); но в свете соображении, изложенных выше, допустимо полагать, что это явление имеет более общую основу — ограниченность спектра потенциальных возможностей первичного синтеза, допустимых для данного генотипа как целостной видовой системы при всех его элементарных мутационных изменениях.
ЭПИГЕНЕТИЧЕСКАЯ ТЕОРИЯ ЭВОЛЮЦИИ
Выше подчеркивалось, что вопрос о происхождении нормальной (адаптивной) организации, составляющий главную проблему эволюционной теории, сводится к объяснению того, каким образом новые свойства организмов приобретают устойчивость, становясь необратимыми. Это объяснение в общей форме дает концепция эпигенетической системы, в рамках которой элементарное эволюционное изменение означает переход индивидуального развития на одну из аберрантных траекторий с последующим преобразованием ее в канализированный путь развития (креод). Теория стабилизирующего (канализирующего) отбора, рассматривающая такой ход событий как основу всего эволюционного процесса, может быть названа эпигенетической теорией эволюции (Шишкин, 1984а). Это название оправданно, во-первых, потому, что инициирующим фактором эволюции здесь признаются нарушения хода онтогенеза. Во-вторых, оно позволяет избежать ошибочного представления, согласно которому стабилизирующий отбор есть лишь один из частных эволюционных механизмов наряду с «ведущим», «дизруптивным» и другими формами отбора, а учение Шмальгаузена—Уоддингтона составляет просто раздел синтетической теории, рассматривающий изменения в устойчивой или колеблющейся среде. Хотя отличия эпигенетической теории от традиционных взглядов и подчеркивались ее авторами, они все же никогда не обобщались, а само ее изложение осталось в большой мере засоренным чуждыми для нее понятиями. Поэтому вначале необходимо кратко охарактеризовать ее содержание.
В основе эпигенетической теории лежит представление об адаптивной норме, или типичной организации, как объекте эволюционных изменений. Под адаптивностью, или целесообразностью организации, понимается ее способность к самоподдержанию и самовоспроизведению (наследованию), т. е. ее устойчивость. Последняя, в свою очередь, выражается в способности нормального индивидуального развития релаксировать в широких приделах внешние и внутренние возмущения на пути к осуществлению данной взрослой организации. Объяснение этого свойства развития и должно составлять ключевую задачу эволюционной теории.
Эпигенетическая концепция решает эту проблему, исходя из представления, что видоспецифичное индивидуальное развитие есть целостная динамическая система с ограниченным и структурированным пространством возможных конечных состояний, среди которых нормальный (адаптивный) финал соответствует равновесию системы, а вся область потенциальных фенотипических уклонений — ее более или менее неустойчивым флюктуациям. Любое отклонение итога развития от равновесия в ответ на повреждающее воздействие всегда представляет собой системную реакцию, выбор которой определяется не спецификой повреждающего фактора (например, типом мутации), а лишь мерой, местом и временем возмущения, вносимого им в ход развития.
Вся совокупность возможных состояний системы (рис. 1, 7), или ее фазовое пространство (эпигенетический ландшафт), составляет ее целостную характеристику, определяемую общей организацией (генотипом) зародышевой клетки и неразложимую на независимые эффекты действия каких-либо элементарных факторов внутри этой клетки или ее онтогенетических производных (Гурвич, 1944; Goldschmidt, 1940; Waddington, 1957). Иначе говоря, возможные альтернативные состояния видового фенотипа — это параметры системы развития, а состояния ее отдельных взаимодействующих элементов (начиная с элементов структуры генома) — динамические переменные, характеризующие нижний иерархический уровень системы. Огромному многообразию комбинаций значений элементарных составляющих (варьирующих в каждом индивидуальном цикле развития) соответствует на верхнем уровне ограниченное пространство вариаций фенотипических параметров, т. е. последние образуют по отношению к этому многообразию инвариантную (эквифинальную) совокупность. При этом чем выше относительная ус тойчивость (вероятность осуществления) данной фенотипической аберрации, тем шире спектр возможных состояний нижнего уровня, в рамках которого она реализуется. Нормальный фенотип, соответствующий равновесию системы, по степени эквифинальности резко превосходит все остальные, т. е. в нормальных условиях он реализуется подавляющим большинством индивидуальных вариантов зигот, существующих в природных популяциях.
Эти представления о системном механизме индивидуального развития опираются на опыт экспериментальной эмбриологии, популяционной генетики и феногенетики, показавший высокую устойчивость нормального развития по отношению к вариациям морфогенетических процессов и генетической структуры зародышевой клетки, а также взаимозаменяемость различных комбинаций внешних и внутренних факторов при осуществлении одних и тех же аберрантных фенотипов. Единственной альтернативой такому пониманию развития может быть представление, согласно которому каждая особенность (или состояние) фенотипа детерминируется в онтогенезе своим особым причинным фактором или их суммой. Это — преформистская модель развития Ру—Вейсмана, несостоятельность которой была доказана экспериментальной эмбриологией.
Изложенные взгляды на природу индивидуального развития предполагают ряд неизбежных следствий.
1. Ни одно возмущение в системе развития, вызванное внутренним или внешним агентом (мутация, воздействие среды) не в состоянии изменить свойств самой системы. Оно либо релаксируется в ходе индивидуального развития, либо приводит к изменению выбора онтогенетической траектории в пределах, определяемых пространством возможностей данной системы. Таким образом, мутация не может быть приравнена к эволюционному изменению (преобразованию системы).
2. Мутация (как и вообще любое возмущение) не детерминирует определенного фенотипа, поскольку каждый из них есть целостная реакция системы, осуществляемая при разных комбинациях значений элементарных взаимодействующих причинных факторов. В свою очередь, каждое из этих значений может при тех или иных условиях соучаствовать в осуществлении любого из фенотипов, «разрешенных» данной системой. Поэтому никакое фенотипическое изменение не может быть описано в редукционистских понятиях (в частности, и в терминах аллельных состояний генов).
3. Все индивидуальные исходы развития, реализуемые данной видоспецифичной системой, основаны на более или менее неидентичных комбинациях элементарных причинных факторов (как в случае ксеногамных популяций, так и клонов). Поэтому любая группа изореагентов (циклов развития с одинаковым фенотипическим результатом) всегда представляет собой генетически неоднородную выборку.
4. Любое уклонение развития от равновесной траектории, независимо от вызвавшей его элементарной причины (мутация или внешнее воздействие), есть нарушение устойчивости данной индивидуальной системы. Это означает большую или меньшую нестабильность воспроизведения (наследования) соответствующего аберрантного фенотипа в ряду потомков данной особи и в конечном счете — его вытеснение или поглощение более устойчивой нормой (т. е. релаксацию уклонения на популяционном уровне).
Итак, подлинно эволюционное изменение — это изменение структуры системы развития (характеризуемой определенным рисунком допустимых для нее онтогенетических траекторий), но отнюдь не флюктуирование в пределах существующей структуры, к которому сводится эффект элементарных мутаций. Именно понимание этой сути проблемы привело в свое время Гольдшмидта (Goldschmidt, 1940) к идее «системных мутаций», предполагающей реорганизацию системы путем скачкообразной перестройки всего хромосомного аппарата зародышевой клетки. По существу это был неудачный ответ на правильно поставленный вопрос о механизме эволюционного преобразования пространства возможностей развития.
Эпигенетическая концепция решает эту проблему иначе. Прежде всего подчеркнем, что перестройка системы развития означает изменение ее равновесного состояния (адаптивной нормы) и тем самым — появление новой канализированной онтогенетической траектории, реализующей это состояние. Положение последней в фазовом пространстве системы, в свою очередь, определяет свойства самого этого пространства, поскольку все существующие в нем аберрантные пути развития суть ответвления канализированной траектории. В переводе на язык традиционных понятии это означает, что характер адаптивного фенотипа определяет собой спектр аберративной изменчивости.
Таким образом, сдвиг адаптивной нормы есть всегда показатель перестройки структуры (эпигенетического ландшафта) системы. При этом очевидно, что между каждыми двумя последовательными историческими состояниями этой структуры (соответствующими минимальному сдвигу нормы) должна существовать прямая преемственность. Новая траектория равновесия не появляется на пустом месте и должна соответствовать одному из аберрантных онтогенетических путей, существовавших в рамках прежней структуры (рис. 7). Его стабилизация рассматривается теорией как результат естественного отбора в пользу соответствующей фенотипической аберрации (морфоза), осуществляемой частью зигот в экстремальных условиях развития. Процесс отбора должен вести к постепенному росту надежности реализации данного уклонения в ряду поколений, т. е. к повышению его наследуемости. Этот эффект получил название «генетической ассимиляции признаков» (Waddington, 1953, 1957). Таким образом, перестройка системы развития, начинаясь каждый раз с выбора одного из фенотипических вариантов, неустойчиво реализуемых в рамках наличной организации видового генотипа, приводит в конце концов к преобразованию самой этой организации, т. е. система перестраивается в направлении от взрослого фенотипа к зиготе (Шишкин, 1981, 1984а, б). «Не изменения генотипа определяют эволюцию и ее направление. Напротив, эволюция организма определяет изменение его генотипа» (Шмальгаузен, 1940б, с. 57). В сумме эволюция рассматривается как процесс репарации устойчивости нормы, периодически расшатываемой историческими изменениями среды; эта репарация достигается каждый раз ценой селективного преобразования самой нормы вместе с реализующей ее системой развития.
Остановимся теперь подробнее на механизме установления нового равновесия системы, или, что то же самое, на стабилизирующем действии отбора. Всякое необратимое изменение внешних условий в сторону предела, ограничивающего возможности канализированного развития в рамках данной системы, ведет к реализации разнонаправленных уклонений, спектр и относительная частота которых определяются свойствами этой системы. Любое различие этих фенотипов по жизнеспособности в данных условиях дает селективное преимущество определенному их классу. Но поскольку этот фенотип, как и все другие реализуемые классы аберраций, относится к неравновесным состояниям системы, то его воспроизведение на первых стадиях селекции оказывается крайне нестабильным, т. е. принадлежащие к нему изореагенты практически повторяют в своем потомстве весь тот спектр уклонений, из которого они сами были перед этим отобраны. Отбор на этом этапе как бы черпает решетом воду. Сохранение адаптивно ценной аберрации в ряду поколений обязано здесь не столько большей эффективности ее индивидуального наследования по сравнению с другими морфозами, сколько ограниченности самого аберративного пространства данной системы, в результате чего отбираемый вариант вновь неизменно воспроизводится ею в числе остальных.
Каждый элементарный шаг отбора, охватывающий два поколения, означает преимущественное сохранение особей, сумевших воспроизвести фенотип своих ранее отобранных родителей, несмотря на комбинирование их гамет при скрещивании (у ксеногамных организмов) и различные другие генетические изменения в процессе самого гаметообразования (мейотическая рекомбинация и ошибки репликации). Поэтому история любого фенотипа, сохраненного длительным отбором, — это цепь последовательных испытаний его носителей на способность воспроизводить самих себя в условиях непрерывного изменения пространства вариаций их геномов. В результате чем большим числом поколений отбора отделены такие носители от исходной группы изореагентов, давшей им начало, тем больший размах генетических изменений данная линия оказалась способной выдержать, не меняя своего фенотипа. Это означает, что генотип вновь созданной («ассимилированной») линии перестраивается отбором в направлении все более помехоустойчивого осуществления данного фенотипа. В соответствии с этим последний начинает все эффективнее поглощать в скрещиваниях остальные аберрации, свойственные данной системе развития, т. е. реагирует на гибридизацию с ними как на регулируемые онтогенетические помехи. Сохраняемое уклонение становится, таким образом, все более однозначно наследуемым, превращаясь в новую норму.
Это преобразование нормы посредством отбора по существу представляет собой выражение общей способности системных объектов релаксировать вызванные в них возмущения, т. е. изменяться целенаправленно. Восстановление равновесия, или «поиск цели» (Эшби, 1962), осуществляется системой путем последовательной коррекции ее состояния, ведущей к затуханию исходного возмущения. Именно такая ситуация, но только связанная с качественном изменением самой системы, возникает в процессе эволюционного сдвига нормы. При переходе популяции в экстремальные условия ее система развития дестабилизируется и вместо реализации прежней нормы переходит к беспорядочным и неустойчивым индивидуальным флюктуациям. Дальнейшее выживание системы в новых условиях зависит от того, удастся ли ей стабилизироваться в каком-либо из этих изменчивых состояний. Этот поиск нового равновесия осуществляется системой посредством преимущественного сохранения (отбора) индивидуальных вариантов развития, реализующих наиболее жизнеспособную флюктуацию. Процесс отбора является здесь не чем иным, как цепью затухающих циклов коррекции с обратной связью, приводящих к стабилизации новой нормы. Каждый акт сохранения отбором носителей адаптивно ценной аберрации есть сдвиг состояния системы в сторону будущего равновесия; «шумы» при воспроизведении этого фенотипа в следующем поколении означают новое отклонение от равновесия; очередной акт просеивания снова сдвигает облик популяции в направлении будущей нормы и т. д. до тех пор, пока каждое вновь воспроизводимое поколение не станет фенотипически однородным и подобным родительскому. Адаптивно ценное изменение становится устойчивой характеристикой системы.
Все это показывает, что созидательная роль отбора, как и каждого творческого процесса, заключается в конечном счете в «запоминании случайного выбора» (Кастлер, 1967), которое выражается в данном случае в выборе одной из относительно равновероятных флюктуаций системы развития и превращении ее в стабильно осуществляемую новую норму. Последняя на всем протяжении своего становления играет роль «цели», определяющей направление коррекции свойств системы в ходе ее преобразования отбором.
Эти представления коренным образом отличаются от традиционного истолкования отбора как механизма просеивания и комбинирования «наследственных изменений», понимаемых как специфические эффекты определенных генов и их сочетаний. При таком подходе объяснение наследуемости эволюционных новшеств становится излишним, поскольку в ней видят просто имманентное свойство соответствующих генов, не зависящее от отбора. Вся процедура «создания» отбором элементарного изменения по существу приравнивается здесь к одноактному выбору его причинного фактора, за которым должно следовать автоматическое воспроизведение нового признака в поколениях. Напротив, для эпигенетической теории устойчивость воспроизведения (наследуемость) — это и есть то, что требует объяснения на основе принципа естественного отбора.
Этапы элементарного сдвига адаптивной нормы, отражающие перестройку видоспецифичной системы развития (размывание старой и стабилизацию новой равновесной траектории и связанное с этим преобразование фазового пространства системы) специально рассмотрены выше (рис. 7). Но этот процесс может быть описан и несколько иначе — с точки зрения изменений, претерпеваемых совокупностью индивидуальных онтогенетических циклов, принадлежащих данной системе (рис. 10).
Рис. 10. Преобразование индивидуальных модификационных спектров в ходе элементарного сдвига адаптивной нормы
1 — соотношение между зиготами и реализуемыми фенотипами; 2 — модификационные спектры, характеризующие серию зигот в одном и том же интервале условий. I — стабильное (эквифинальное) осуществление группой зигот исходной адаптивной нормы А в средних допустимых для нее условиях N; II — дестабилизированное развитие таких же зигот в интервале экстремальных условий N1, где среди реализуемых уклонений (b), с, d, k, т, z морфоз (b) имеет адаптивное преимущество; III — стабилизация морфоза (b) и превращение его в адаптивную модификацию В в рамках новой полиморфной нормы АВ; IV — дальнейшая стабилизация фенотипа В на фоне утраты прежней нормы или сохранения ее в качестве неустойчивого морфоза (a). F1 — F2 — Fn— Fx — смена поколений; заштрихованы осуществленные типы развития; их элиминируемые варианты показаны нисходящими стрелками. F1— F2 — вскрытие изменчивости; F2 — Fn, Fn — Fx — этапы стабилизирующего отбора
Поскольку вероятности осуществления одних и тех же путей развития для разных зигот одного вида всегда неодинаковы в силу их генетических различий, то в любом конкретном диапазоне условий, выходящих за рамки обычных, каждая из них обнаруживает свой собственный спектр реализуемых фенотипических уклонений (морфозов). С этой точки зрения популяция нормальных зигот может быть представлена как серия разнородных модификационных спектров, в которых основную часть всегда составляет адаптивный фенотип, а краевые отрезки — различные сочетания морфозов (рис. 4; 10, I). В обычных условиях все зиготы развиваются эквифинально, реализуя норму (рис. 10, I). По мере изменения среды в сторону критического порога канализированное развитие сменяется разнонаправленным, т. е. возникает все большее число морфозов в соответствии со спецификой индивидуальных спектров (рис. 10, II). При сохранении таких условий в ряду поколений начинается отбор в пользу наиболее жизнеспособного морфоза, ведущий к постепенному росту его устойчивости и дестабилизации прежней нормы (рис. 10, III). Элиминация остальных типов аберрантных реакций вначале малоэффективна, поскольку они вновь возникают в потомстве отбираемого варианта ввиду его слабой онтогенетической устойчивости. Но по мере стабилизации отбираемого фенотипа его наследование становится все более однозначным, и он (в случае ксеногамного размножения) все более поглощает в скрещиваниях остальные аберрации, остающиеся неустойчивыми. Это закрепление адаптивно цепной реакции, превращающее ее в новую норму, приводит к тому, что в модификационных спектрах последовательных поколений зигот она занимает все большее место за счет старой нормы. Таким образом, обе нормы сосуществуют на определенном этапе в индивидуальных спектрах как две адаптивные модификации, реализуемые в зависимости от колебаний условий (рис. 10, III), пока, наконец, новая полностью не возобладает. Возможность реализации прежнего нормального фенотипа в установившихся новых условиях если не исчезает совсем, то низводится до уровня аберрантной атавистической вариации (рис. 10, IV).
Таким образом, каждый элементарный шаг в селективном преобразовании нормальной организации сводится к экстремальному разнонаправленному модифицированию (дестабилизации) существующей нормы и последующему фиксированию наиболее оптимального из выявившихся вариантов индивидуального реагирования. Модифицирование и стабилизация «непрерывно кооперируются» (Шмальгаузен, 19686, с. 315). Первая фаза этого шага означает индивидуализацию фенотипического выражения отдельных геномов (рис. 10, I—II), вторая — его унификацию, т. е. создание механизма канализированного развития новой адаптации, нивелирующего частные генетические различия (рис. 10, III—IV). Первая из этих фаз есть период неустойчивости, разделяющий каждые два последовательных устойчивых состояния нормы.
Все это позволяет осознать, что, говоря о движущей функции отбора, мы подразумеваем конечный результат этого процесса, а не его механизм. Первичное нарушение прежней нормы, как мы видели, вообще связано не с отбором, а с нарушением условий развития, ведущим к проявлению скрытой изменчивости; создание же новой нормы не сводится к одному лишь выбору оптимального варианта изменения. Отбор не может непосредственно сохранять неустойчивые реакции, составляющие скрытый резерв изменчивости. Сдвиг нормы в пользу одной из них осуществляется не за счет непосредственной элиминации других (ибо вначале они возникают в поколениях вновь и вновь), а за счет стабилизации оптимальной реакции, которая становится вследствие этого все более универсальной для всей совокупности развивающихся особей. Таким образом, движущего отбора как особого процесса по отношению к стабилизирующему не существует. Это понятие описывает общий итог длинного ряда чередующихся фаз вскрытия резерва изменчивости и стабилизации определенных вариантов изменений (Шишкин, 1984а, б). Если, по Дарвину (1952, с. 139), естественный отбор есть «сохранение полезных индивидуальных изменений», то это означает не что иное, как приобретение ими онтогенетической устойчивости. Указание Шмальгаузена (1968б) о неразрывной связи движущей и стабилизирующей форм отбора (к сожалению, оставшееся неконкретизированным) нуждается в уточнении. Речь идет не только о двух сторонах, но о двух разнокачественных и разномасштабных измерениях одного процесса.
В том, что движущий отбор действительно основан на стабилизации частных аберраций нормы, убеждает широкий круг наблюдений над ходом ее преобразования в различных природных и экспериментальных условиях. Наиболее строгое подтверждение дают рассмотренные выше опыты по генетической ассимиляции структурных морфозов (Waddington, 1957), поскольку здесь не остается сомнений в предельно неустойчивом характере исходного класса реакций, формирующих новую норму. Другую важную категорию фактов составляют многочисленные эксперименты по ассимиляции физиологических морфозов, т. е. по принудительному селективному адаптированию различных групп организмов (обычно насекомых) к новым факторам среды, например, опыты М. М. Камшилова (1941) по воспитанию холодоустойчивости у дрозофилы, И. В. Кожанчикова (1941) по выращиванию жуков-листоедов на непривычном корме, подобные же опыты Г. В. Самохваловой (1951, 1954) и особенно Г. X. Шапошникова (1961, 1965) по смене хозяина у тлей и т. д. [Наблюдаемое при этом развитие новых стойких адаптации нередко приписывают «длительным модификациям» в тех случаях, когда используются партеногенетические клоны (например, в ряде опытов с тлями), поскольку их принято считать наследственно однородными. Но пестрота их индивидуального реагирования в экстремальных условиях и весь последующий ход преобразовании явно указывают на скрытую гетерогенность. Последняя возникает за счет ошибок репликации генома, которые неизбежны при любом способе формирования генеративных клеток]
Первой реакцией на резкую смену условий является, как правило, высокая смертность или снижение плодовитости и, несомненно, экстремальное физиологическое состояние выживающих особей. В ходе отбора на устойчивость этого состояния оно из сублетального превращается в одно из обычных или даже оптимальное. Совершенно очевидно, что эта толерантность к ранее вредному фактору возникает здесь как новое качество, созданное отбором и не существовавшее внутри прежней нормы в виде готовой наследственной вариации. Например, в опытах с охлаждением личинок дрозофилы в исходной популяции не было особей, устойчиво резистентных к холоду или предпочитающих умеренно пониженную температуру для развития (Камшилов, 1941, 1979), а в опытах с тлей Dysaphis anthrisci majkopica вплоть до 8–11 поколений воспитания на новом хозяине не существовало особей, предпочитающих его старому (Шапошников, 1961, 1965). Последовательность изменений, наблюдаемых в подобных случаях (в частности, в опытах с Dysaphis), хорошо соответствует теоретически ожидаемому ходу событий. Вначале фенотипически однородная популяция, попавшая в необычные условия, показывает резкое повышение изменчивости, включая дифференциальную плодовитость и жизнеспособность. Это — фаза вскрытия мобилизационного резерва изменчивости (рис. 10, I—II). Далее наблюдается все большая унификация фенотипов в пользу новой адаптивной нормы, причем сначала ряд поколений не оказывает предпочтения старому или новому варианту условий. Это — переходная фаза сосуществования обеих норм в одних и тех же индивидуальных спектрах в качестве двух адаптивных модификаций (рис. 10, III). Наконец, новая норма окончательно стабилизируется в качестве единственной адаптации (ср. рис. 10, IV). Ее обособление от носителей прежней нормы может достигать уровня репродуктивной изоляции (Шапошников, 1978).
Для описанных преобразований известно огромное количество природных аналогий, в большинстве относящихся к случаям «привыкания» различных форм насекомых к пестицидам. Генетическая природа возникающей при этом резистентности к определенному фактору может быть в разных случаях различной (Дубинин, 1966а). В итоге возникают новые расы, показывающие при скрещивании с исходными разные типы расщепления (Dobzhansky, 1947). Нет никаких оснований полагать, что процесс их становления отличается от вышеописанного и что новое свойство существовало в устойчивой форме у единичных мутантов до начала отбора.
Между тем, как уже говорилось ранее, именно это предположение составляет основу господствующих представлений о механизме движущего отбора. В нем видят лишь процесс накопления мутаций с наследуемым фенотипическим эффектом, т. е. считается, что последний не создается отбором, а лишь подхватывается им. В частности, появление устойчивой к пестицидам расы насекомых, если она отличается от исходной одним менделевским фактором, рассматривается как самоочевидный результат распространения единичной мутации резистентности (Dobzhansky, 1947, с. 190). Не касаясь пока теоретической стороны этих взглядов, остановимся на их соотношении с наблюдаемыми фактами. Во-первых, они противоречат опыту экспериментальной генетики, показывающему, что эффекты «сырых» мутаций, в особенности малых (составляющих потенциальный материал эволюции), неустойчивы по сравнению с адаптивной нормой и в той или иной степени поглощаются ею. Во-вторых, заслуживает внимания сам характер используемых доказательств. Последние обычно сводятся к демонстрации изменений частоты определенного признака в разных популяциях, расах или других группах, прослеживаемых либо в пространстве (вариации типов окраски тела, жилкования крыльев и т. п.; Zimmermann, 1933; Тимофеев-Ресовский и др., 1965), либо даже в геологическом времени (например, изменения морфотипов зубов в эволюции млекопитающих; Simpson, 1953). При этом подразумевается, что минимальные частоты признака соответствуют его существованию в виде мутаций, а более высокие — его вхождению в состав нормы. Но если между мутантным фенотипом и его адаптивным аналогом не предполагается различий в онтогенетической устойчивости, то тем самым мы заранее приравниваем акт мутирования к созданию элементарной адаптации вместо того, чтобы это доказать.
На самом деле задача состоит как раз в том, чтобы выяснить, какова была фактическая наследуемость «мутантной» аберрации до начала ее экспансии и каково вообще ее происхождение. Была ли она всегда устойчива или же представляла сначала лабильную реакцию, постепенно стабилизированную и усиленную отбором? В большинстве случаев мы этого проверить не можем, и, говоря о распространении «мутаций», исследователи подразумевают сложившиеся адаптивные экотипы (как, например, в случае меланизма у хомяков; Гершензон, 1946), внутри которых обычно выявляется наследственная пестрота (Дубинин, 1966а, с. 280). Но там, где предыстория таких форм хотя бы отчасти известна, мы не находим доказательств их изначальной устойчивости. Примером может служить эволюция индустриального меланизма у березовой пяденицы Biston betularia, представляющая, казалось бы, хрестоматийный случай «включения полезной мутации в норму». Современная меланистическая морфа этой бабочки в Англии, преобладающая в большинстве популяций, доминирует над светлой морфой и дает с ней монофакторное расщепление. Но ни одно из этих свойств не является исторически первичным. У более ранних меланистов (как полагают, гибридов), пойманных полтора века назад, окраска была светлее современной, т. е. доминирование было неполным, даже несмотря на то, что уже тогда темная окраска, очевидно, входила в полиморфную норму, давая адаптивное преимущество на темноствольных деревьях (Kettlewell, 1956). Ни доминирования, ни правильного расщепления не обнаруживается также и при скрещивании английской темной морфы Biston со светлой из Канады (где меланистов еще нет); вместо этого имеет место промежуточное наследование (Шеппард, 1970). Доминирование есть выражение устойчивости фенотипа (Шмальгаузен, 1982, 1968б), и оно, несомненно, усиливалось в ходе становления темной морфы Biston. Экстраполируя от двух известных ее последовательных состояний (в Европе) к начальному моменту ее адаптивной истории, мы можем с большим основанием считать, что исходным для нее материалом послужили слабо выраженные уклонения от светлой окраски, чувствительные к колебаниям внешних факторов и генетической конституции особей, т. е. однотипные морфозы нормы, которые отбор стабилизировал и усиливал шаг за шагом. Потенциальная способность к такой меланистической реакции вообще широко распространена у бабочек (Дубинин, 1966а), и ее можно вызвать искусственно, например, охлаждением гусениц (Standfuss, 1902). Ясно, что нет никаких оснований приписывать свойства современных нормальных меланистов элементарным мутациям с устойчивым эффектом, возникшим у их предков.
Еще один общий источник предполагаемых доказательств появления стойких адаптации путем единичных генетических изменений составляют биохимические мутации у низших организмов, в частности у бактерий. При пересеве той или иной культуры на неполноценную среду (например, штамма кишечной палочки, сбраживающего галактозу, на среду с лактозой) выживают лишь отдельные мутантные клетки, причем методом реплик можно показать, что адаптивно ценное изменение не индуцировано новыми условиями, а существовало в исходной культуре (Дубинин, 1966а, 1976). Налицо как будто бы спонтанное появление новой адаптации. Но нетрудно увидеть, что здесь отбирается не стойкое приспособление, а просто одна из элементарных модификаций. Так, в нашем примере способность сбраживать лактозу проявляется у мутантных клеток (в той или иной степени) только на соответствующем провокационном фоне, тогда как на исходной галактозной среде они осуществляют свою прежнюю нормальную функцию; иначе такие клетки погибли бы до пересева. В модификационных спектрах мутантов данного типа обе реакции присутствуют одновременно, и выбор между ними определяется только внешними условиями. Устойчивое же закрепление новой реакции, очевидно, может происходить лишь в ходе размножения измененного штамма в новой среде по мере отбора среди множества индивидуальных клеток. Последняя возможность допускается для многих случаев, когда ясно видна постепенность приспособления бактерий к растущим дозам вредного фактора (Дубинин, 1976). Но, по-видимому, не всегда осознается, что в этом и состоит общая закономерность становления новых адаптации. Нужно учитывать, что при высокой скорости размножения бактерий стабилизация отобранных морфозов в новой среде происходит крайне быстро, затемняя их исходную неустойчивость.
Таким образом, представление об эволюции как отборе фенотипически стабильных геновариаций строится на фактах, которые не могут его доказать. Эти факты характеризуют не процесс адаптациогенеза как таковой, а либо его исходную основу (проявления биохимических мутаций), либо конечный результат, т. е. изменения частот уже сложившихся компонентов полиморфных систем. В действительности же становление новых приспособлений невозможно без стабилизации индивидуальных реакций, дающих им начало.
Но если материал эволюции составляют лабильные реакции (модификации), осуществляемые нормообразующими генотипами лишь в уклоняющихся условиях, то ясно, что они, по определению, не подчиняются менделевским правилам. Скрещивание двух элементарных морфозов нормы не может дать стойкого результата, несмотря на генетические различия их носителей. В случае же скрещивания такого морфоза с устойчивым нормальным фенотипом первый будет, естественно, поглощаться вторым (при развитии потомства в нормальных условиях). Единственная природная ситуация, где правильное менделевское расщепление имеет место (если исключить наиболее грубые мутационные нарушения),— это скрещивание вариантов нормы, т. е. стабилизированных фенотипов, составляющих полиморфную систему. Только в этом случае в эпигенетических системах гибридов возникают альтернативные пути устойчивого развития (креоды), между которыми осуществляется упорядоченный выбор. Коротко говоря, менделируют лишь продукты канализированного развития (адаптивные морфы), но не продукты его дестабилизации (морфозы). Поэтому для эпигенетической теории эволюции мир природной изменчивости как целое не описывается в менделистских терминах. Его составляют не аллели (в которых теория видит лишь отношения между определенными типами развития), а только сами фенотипы, т. е. варианты нормы и их аберрации. Поведение отдельных фенотипов в скрещиваниях зависит исключительно от их принадлежности к этим двум классам, характеризующим тип индивидуального развития, а также от самих условий развития. Дарвиновская неопределенная изменчивость есть совокупность аберраций нормы и тем самым — это область фенотипов, не имеющих упорядоченного наследования. Различие, описываемое в менделевских аллелях, это результат отбора по альтернативным фенотипам, а не просто любое проявление генетического различия.
С этих позиций коренным образом меняется объяснение причин, по которым полезные элементарные изменения не растворяются в скрещиваниях и могут сохраняться отбором. В истории критики дарвинизма предположение о таком растворении известно как «кошмар Дженкина», и на протяжении всего нашего столетия эволюционисты не устают повторять, что этот аргумент устранен открытием менделевских законов с их принципом «чистоты гамет». Но эти законы универсальны для хромосом, а не для признаков и неприложимы к неопределенной фенотипической изменчивости. Истинная причина действенности отбора заключается не в высокой наследуемости и строгой дискретности используемых им элементарных изменений, а в принадлежности их к ограниченному пространству аберраций, свойственных данной фенотипической норме. В результате эти малоустойчивые сами по себе вариации суммарно повторяются в поколениях, обеспечивая стабильность потенциального субстрата отбора без стабильности индивидуального наследования. Отбор в пользу одной из них ведет не к сохранению ее «несмешиваемых факторов», а лишь к увеличению численной роли гамет, оставляемых ее носителями независимо от их индивидуальной генетической конституции, меняющейся в каждой генерации. На первых этапах процесса эти гаметы способны воспроизводить отбираемый фенотип лишь в числе других вариантов, составляющих исходную гамму аберраций; однако каждый раз их комбинирование повышает вероятность его дальнейшего осуществления. По мере продолжения отбора на устойчивость самовоспроизведения этого фенотипа (т. е. в результате преимущественного сохранения в каждом поколении тех его потомков, которые сумели его повторить) постепенно возрастает его индивидуальная наследуемость. Этот процесс основан на непрерывной перестройке генотипа гамет в поддерживаемой отбором линии.
Достигнутая таким путем высокая наследуемость фенотипа, основанная на его стабилизации, проявляется теперь в его подавляющем доминировании при скрещиваниях с различными случайными вариациями, чем и демонстрируется его «нерастворимость». Если же подобным образом из начального спектра отбирается сразу несколько фенотипических вариантов, то в последовательных поколениях их гибридизация будет приводить ко все более упорядоченному их наследованию в определенных соотношениях. Таким образом, менделевское расщепление по фенотипам — это не причина сохранения эволюционных новшеств, а результат их стабилизации в ходе отбора.
* * *
Изложенные представления об эпигенетическом механизме эволюционного процесса позволяют оценить сущность понятий, лежащих в основе синтетической теории эволюции. Анализ последней затрудняется тем, что со временем все больше нарастает разрыв между ее фактическими представлениями и теми, которые принимаются декларативно, без использования в ее построениях. Очевидно, что предметом рассмотрения должна быть именно первая категория понятий.
Исходным здесь является положение, что субстрат отбора составляют мозаичные менделевские аллельные факторы, отождествляемые с хромосомными локусами. Эволюция есть изменение генного состава популяции, и скорость ее определяется скоростью замещения аллелей (Dobzhansky, 1947; Грант, 1980). Для хромосомных генов подразумевается упорядоченное фенотипическое выражение, оцениваемое количественно как относительный вклад в приспособленность. Устойчивость фенотипов или признаков рассматривается чисто преформистски как свойство контролирующих их генов, и потому ее объяснение не входит в задачи теории. Понятие устойчивости имеет здесь лишь надындивидуальный смысл и выражает либо фиксацию определенных частот аллелей (генетическое равновесие), либо, в более прямом значении, гомозиготизацию по тому или иному из них, т. е. достижение аллелем 100%-ной частоты в популяции. Последнее происходит за счет устранения альтернативных аллелей — в результате отбора или же генетического дрейфа (при ограничении размеров популяции), когда случайные отклонения частот от равновесия могут приобрести необратимый характер. Соответственно предмет рассмотрения теории составляет не организация генотипа и управляемого им индивидуального развития, а организация генофонда популяции.
Понятия генетической обусловленности и наследственности (устойчивости) отождествляются рассматриваемой теорией, т. е. всякое генетическое изменение понимается как фенотипически наследуемое, хотя и признается, что его эффект может искажаться генотипическими и внешними влияниями. Поэтому наследственные изменения рассматриваются как не зависящие от естественного отбора; последний лишь оперирует ими, но не создает их. Ненаследственные, или модификационные, уклонения понимаются как шумы, затемняющие действие генов и тормозящие отбор генетических изменений (Дубинин, 1966а; Майр, 1968; Грант, 1980). Поскольку между последними, по определению, не признается различий в степени устойчивости их эффекта, то понятия фенотипической нормы и аберрации лишаются в теории какой-либо функциональной роли. Для них нет иных характеристик, кроме частот соответствующих фенотипов и генов в популяции. По той же причине представление об эволюционном поглощении одних фенотипов другими через систему скрещиваний является здесь совершенно бессодержательным; речь может идти лишь об их замене путем вытеснения, выражающей замену соответствующих аллелей. Это замещение означает уменьшение генетической дисперсии популяции по приспособленности и соответственно уменьшение генетического груза. Чем большее число генов одновременно затрагивается отбором, тем больше величина груза и тем медленнее должны идти изменения (при условии сохранения достаточной численности популяции).
Поскольку всякая мутация рассматривается теорией как аллельное изменение, контролирующее устойчивый признак, то это означает фактическое признание скачкообразной эволюции, хотя на словах ее часто отрицают (Майр, 1968; Рьюз, 1977; и др.). Ограничения, налагаемые на сальтационизм, носят здесь технический, а не принципиальный характер. Там, где между морфами или расами устанавливается простое аллельное различие, это считается указанием на возникновение одной формы из другой путем единичной мутации (Dobzhansky,1947, с. 50, 190; Грант,1980,с. 88, 174); там же, где предполагается множество таких различий (например, между преемственными видами), эволюционное преобразование рассматривается как последовательность мутаций (Dobzhansky, 1947, с. 52). Именно мутация выступает здесь как созидающий фактор, тогда как творческая роль отбора сводится лишь к гомозиготизации рецессивных генов, выявляющей новые фенотипы, или же к комбинированию генных эффектов, или, наконец, к созданию оптимальной генотипической среды для выражения гена. Эволюция рассматривается здесь как диалог между средой и генами (Новинский, 1978).
Эти редукционистские принципы теории, в особенности отбор генов и наделение их коэффициентами приспособленности, все меньше удовлетворяют ее сторонников и уже давно характеризуются как «сверхупрощения» (Dobzhansky, 1947, с. 106). Но отсюда не следует большой ясности в вопросе о том, что же в таком случае составляет подлинный субстрат отбора. В качестве него указываются генотипы и генные комбинации (Шеппард, 1970), или же только фенотипы (Майр, 1968), или же все это вместе (Грант, 1980). Все это отражает скорее намерение отойти от старых представлений, чем подлинный отход от них. Поэтому не удивительно, что механизм движущего отбора по-прежнему сводят к замещению одного аллеля другим. (Грант, 1980, с. 144). По признанию Р. Левонтина (Lewontin, 1970), современная популяционная генетика, на которой строится синтетическая теория, все еще исходит из представления о чисто мозаичном комбинировании генов, даже не учитывая их организацию в группы сцепления.
Наиболее ясным указанием на сохранение редукционистского понимания сути отбора является характер представлений о результате его действия, свойственный синтетической теории. Всякий эффективный отбор должен приводить к гомогенизации своего субстрата, и, предполагая отбор аллелей, мы, естественно, должны ожидать (в пределе) их перехода в гомозиготное состояние. Но именно это и предполагается синтетической теорией по сегодняшний день (Шеппард, 1970; Айала, 1981; Солбриг, Солбриг, 1982). Несоответствие реальной картины этим ожиданиям (гетерогенность природных популяций) считается указанием на наличие факторов, действующих, «несмотря на силы отбора» (Айала, 1981, с. 51). Даже когда делаются попытки объяснить гетерогенность отбором комплексов модификаторов, выравнивающих приспособленность для максимума вариантов генетического фона (Майр, 1974), речь все равно идет о сохранении суммы генов, ответственной за этот эффект.
Косвенным свидетельством подобного понимания отбора является и сам факт появления нейтралистской гипотезы эволюции (Kimura, 1968), выдвинутой как альтернатива синтетической теории. Одним из ее оснований является, как известно, наличие природного полиморфизма по изоферментам, указывающего на множественность соответствующих аллелей, которая рассматривается здесь как нечто не согласующееся с идеей отбора. Другими словами, благодаря синтетической теории дарвиновский отбор настолько прочно отождествился в умах эволюционистов с отбором аллелей, что отсутствие последнего выглядит для них равнозначным отсутствию отбора вообще и заставляет их говорить о «недарвиновской» эволюции. Это красноречиво показывает, что, несмотря на формальное признание синтетической теорией отбора по фенотипам, «ее идеалы остались прежними» (Новинский, 1978).
Таким же неизменным остается для синтетической теории и признание фиксированных соотношений между геном и признаком (формально отрицаемых хромосомной генетикой). Подчинять природную изменчивость менделевским правилам — это и значит признавать, что законы мейотической рекомбинации хромосомных локусов имеют однозначное выражение в изменениях фенотипов — хотя бы оно и затемнялось внешней или генотипической средой. Далее, поскольку давление отбора измеряется единственно по изменению частоты форм в поколениях (Шеппард, 1970), очевидно, что все суждения об изменении генного состава на этой основе возможны лишь при наличии четкого соответствия между генами и фенотипами. Точно так же и представление о закреплении аллелей (путем отбора или дрейфа) как источнике эволюционных новшеств предполагает ту же самую связь.
С точки зрения эпигенетической концепции исходным пунктом всех этих построений является сведение целостных свойств организма, созданных естественным отбором, к изначальным характеристикам геномных единиц, существующим независимо от отбора. Это выражается не только в наделении генов вкладом в приспособленность и устойчивым фенотипическим эффектом, но прежде всего в самом понимании гена. Менделевский фактор, который объективно выражает различие двух созданных отбором вариантов организации генотипа (контролирующих соответствующие креоды), отождествляется здесь с участком хромосомы, т. е. различие в свойствах двух систем переносится на их элементы. Локус, определяющий переключение креодов в данной системе скрещиваний, превращается в носителя неизменных характеристик, сохраняемых им во всех рекомбинациях. Соответственно мутация локуса превращается в механизм создания менделевского различия, т. е. результат исторического процесса заменяется одноактным событием. Это совершенно та же логика, которая заставляет многих эволюционистов-эмбриологов видеть в расхождении типов онтогенеза у разных групп организмов (онтогенетической дивергенции Бэра) доказательство их скачкообразного происхождения друг от друга путем одноактной девиации (Шишкин, 1981). Для эпигенетической концепции реальный природный мир менделевских соотношений — это лишь совокупность стабилизированных фенотипов, характеризующая полиморфную систему вида и объединенная свободным скрещиванием. С этой точки зрения модель эволюционных событий, принимаемая синтетической теорией, исходит из перенесения на неопределенную изменчивость свойств адаптивной полиморфной системы, т. е. сырой материал эволюции подменяется ее продуктами — мозаикой стабилизированных форм. Отбор здесь «вводится в действие» в условиях, когда его работа уже сделана. Представлению о смене частот аллелей соответствует в природных популяциях лишь одна реальность — изменение концентраций устойчивых морф, дающих в скрещиваниях менделевские расщепления.
Поэтому исходить из конкуренции аллелей как двигателя эволюции — значит оперировать одними адаптивными нормами, тогда как неопределенные аберрации, вызываемые нарушениями развития и составляющие подлинный субстрат отбора, в этом случае вообще устраняются из рассмотрения. Только при подобном ограничении материала скрещивании вариантами нормы или искусственно стабилизированными формами (чистые линии) имеет место ситуация, когда фенотипы не поглощают друг друга, а лишь меняют частоты в ответ на кратковременное действие отбора. И если принять, что устойчивые различия этих фенотипов, выражаемые в менделевских факторах, создаются мутациями, а не отбором, то на долю последнего не останется ничего, кроме распространения или уничтожения фенотипических результатов мутирования. В таком случае действие отбора действительно не должно принципиально отличаться от результатов случайных частотных флюктуаций (дрейфа) аллельных фенотипов, и это объясняет причины самого возникновения идеи дрейфа.
Таким образом, при сравнении двух изложенных систем взглядов на природу эволюционного процесса обнаруживается их диаметральное различие. Если для синтетической теории отбор — это перераспределение аллельных факторов, возникающих помимо него и понимаемых как состояния хромосомных локусов, то для эпигенетической теории результатом отбора, если использовать тот же язык, является создание и размывание аллельных свойств фенотипов в процессе смены адаптивных норм. Иначе говоря, аллели создаются и уничтожаются отбором. Если, например, два фенотипа, описываемые менделевским анализом как гомозиготы АА и аа, дают в скрещиваниях третий фенотип Аа, имеющий селективное преимущество, то для синтетической теории его фиксация в природе не означает ничего большего, чем установление баланса аллелей, при котором численность таких гибридных форм уравновешивается элиминацией менее приспособленных гомозигот. Напротив, для эпигенетической теории отбор в пользу фенотипа Аа означает его стабилизацию по отношению к двум другим, т. е. такую перестройку его генотипа и системы развития, в результате которой он все более эффективно поглощает в скрещиваниях исходные варианты, а значит, и перестает закономерно расщепляться на формы АА и аа. Вместе с канализацией его развития и размыванием (у потомства) альтернативных креодов, соответствующих исходным чистым линиям, утрачивается и сама возможность интерпретации этого фенотипа в терминах аллелей А и а. Если искать аналогии этим взглядам в генетическом мышлении, то можно сказать, что идея «отбора в пользу фенотипа лучшей из гомозигот», принимаемая фишеровской гипотезой эволюции доминантности (Fisher, 1930), расширяется здесь в представление об отборе на доминирование лучшего фенотипа вообще.
Итоговое сравнение двух эволюционных концепций приводит к заключению, что первая из них (эпигенетическая) является по существу гораздо более «генетической», чем та, с которой связывают это определение. Такие фундаментальные эмпирические обобщения генетики, как устойчивость нормы («дикого типа») по сравнению с аберрациями и способность к их поглощению; нарушения менделевского наследования, выражаемые в понятиях экспрессивности и пенетрантности; гетерогенность любых классов фенотипов; зависимость признаков от генотипа в целом; влияние отбора на их доминирование,— все это, как мы видели, относится к основополагающим понятиям эпигенетической теории, но не находит выражения на языке противоположной концепции. Синтетическая теория исходит на деле не столько из реальных достижений хромосомной генетики, сколько из абстракций раннего менделизма, опиравшегося на однозначное соответствие генов и признаков и отождествление эволюционных новшеств с заменой генов.
Эти выводы сегодня уже не кажутся неожиданными и находят свое подтверждение во все усиливающейся тенденции к критической переоценке основ синтетической теории. Генетики все чаще характеризуют ее лишь как «полезную абстракцию» (Камшилов, 1979, с. 141), представляющую собой «временное и упрощенное» построение (Солбриг, Солбриг, 1982, с. 78); видеть в ней действительный синтез генетики и дарвинизма — значит «выдавать желаемое за действительность» (Бабков, 1981, с. 413). Определение эволюции как изменения генетического состава популяции ныне рассматривается как редукционистское (Майр, 1981); в основе изменения признаков предлагается видеть перестройку генотипов, а не просто замещение генов (Майр, 1968). К числу вынужденных упрощений теории эволюционисты-генетики относят сегодня все главные ее принципы — отбор аллелей, оценку их по приспособленности и однозначность соответствия генотипа и фенотипа; анализ генетических изменении по фенотипическому составу признается произвольным и не учитывающим закономерностей индивидуального развития (Солбриг, Солбриг, 1982, с. 77, 78, 154, 263). Используемые теорией модели действия отбора на частоты локуса с двумя аллелями признаются «совершенно нереалистичными» (Там же, с. 258). Увеличивается и скептицизм в отношении роли генетического дрейфа как фактора эволюции (Шеппард, 1970; Солбриг, Солбриг, 1982). Напротив, прежнее отрицание эволюционной роли модификационных изменений сменяется теперь ее формальным признанием (Майр, 1968, 1974), хотя, как справедливо указывалось ранее, это равносильно «пересмотру основных положений генетической концепции эволюции популяций» (Дубинин, 1966а, с. 373). Еще одним свидетельством отхода теории от своих фундаментальных принципов является ее стремление отказаться от идеи генетического груза (Майр, 1974; Грант, 1980; Мейнард-Смит, 1981; Галл, 1980), вызванное трудностями совмещения ее с фактами генетического полиморфизма внутри природных популяций. Но вместе с идеей груза неизбежно устраняется и единственное предлагаемое теорией объяснение механизма движущего отбора как уменьшения дисперсии генетического состава популяции по приспособленности. Понятие отбора в пользу лучших аллелей тем самым лишается содержания. Все эти тенденции объективно отражают одно и то же — несоответствие языка синтетической теории представлениям о целостном организме как предмете естественного отбора. При очевидном отсутствии упорядоченных (преформированных) соотношений между геномными единицами и свойствами организма результаты отбора по фенотипам принципиально не могут описываться в терминах генных частот. Одним из косвенных и независимых признаний этого факта является мысль о замене генов хромосомными континуумами в качестве будущей основы построений популяционной генетики (Lewontin, 1970).
Чем больше выдвигается поправок, направленных на модернизацию синтетической теории, тем труднее становится определить, что же составляет ее сущность. Теория не может оставаться сама собой, отказываясь от собственных основ. Создается впечатление, что для нее в действительности нет иного логического фундамента, кроме последовательного преформистского редукционизма, связывающего гены с признаками, и что вне его она превращается в эклектическое построение. Очевидно, именно поэтому, несмотря на все поправки, она остается для критиков концепцией, абстрагирующейся от проблемы целостности организма и его индивидуального развития, и в качестве таковой признается неприемлемой для объяснения эволюции, по крайней мере на уровне ее макрохарактеристик (Bertalanffy, 1969; Но, Saunders, 1979; Alberch, 1980, 1982; Wake et al., 1983; Maderson et al., 1982; Блюменфельд, 1974; Новинский, 1978).
Очевидная невозможность совместить дарвиновское представление о естественном отборе с упрощениями мутационизма и раннего менделизма вызывает не только стремление ограничить роль этих упрощений путем пересмотра синтетической теории. Наряду с этим наблюдаются противоположные попытки — решить возникающее противоречие за счет дарвинизма. Так, М. Д. Голубовский (1981), почти повторяя слова Иогансена об устранении генетикой основ дарвиновской теории (Филипченко, 1977, с. 191), считает, что каждый шаг в развитии учения о наследственности означал последовательное ограничение постулатов селекционизма. По его мнению, нет дарвиновской неопределенной изменчивости — ее заменила строгая упорядоченность расщеплений, ограниченная менделевскими правилами и не зависящая от отбора. Нет созидательной роли отбора, ибо он может сохранять лишь то, что создается мутациями (как один из примеров приводится индустриальный меланизм у Biston betularia). Поэтому нет и постепенности эволюционных изменений — ее место должны занять мутационные скачки, оцениваемые отбором. Этот призыв вернуться к представлениям де Фриза и Иогансена не кажется случайным; он отражает, хотя и в своеобразной форме, ясное понимание иллюзорности того, что сегодня называется синтезом дарвинизма и генетики.
Подлинная ассимиляция достижений генетики дарвиновским учением не может строиться на абсолютизации выявленных ею закономерностей (как и вообще любых форм биологической упорядоченности) в качестве универсальных факторов, действующих вне связи с естественным отбором. Наоборот, они должны быть истолкованы как результаты отбора, не существующие вне его. Задачей эволюционной теории является не только вскрытие биологических законов, но и определение условий их выполнения, т. е. осознание их относительности. Поэтому, говоря об эволюционном объяснении данных генетики, мы должны подразумевать все реальное многообразие накопленных ею фактов, а не только сумму правил, используемых для гибридологического анализа чистых линии. Единственной приемлемой основой такого объяснения представляется эпигенетическая теория Шмальгаузена—Уоддингтона, рассматривающая наследственность как выражение устойчивости индивидуального развития, создаваемой естественным отбором. Можно утверждать, что именно с этой теорией, ставящей в центр внимания свойства целостной системы развития организма и стремящейся к описанию фактов эмбриологии и генетики на одном языке, связаны перспективы будущего развития дарвинизма.
ЛИТЕРАТУРА
Агаев М. Г. Экспериментальная эволюция. Л.: Изд-во ЛГУ, 1978.
Айала Ф. Механизмы эволюции // Эволюция. М.: Мир, 1981. с. 33–65.
Астауров Б. Л. Генетика пола // Актуальные вопросы современной генетики. М.: Изд-во МГУ, 1966. С. 65–113.
Астауров Б. Л. Homo sapiens et humanus — человек с большой буквы // Новый мир. 1971. № 10. С. 214–229.
Бабков В. В. Системный стиль в изучении естественного отбора // Системные исследования. М.: Наука, 1981. С. 404–419.
Балкашина Е. И. Случай наследственного гомеозиса. Геновариация aristopedia у Drosophila mеlanogaster // Журн. эксперим. биологии. 1928. Т. 4. вып. 2.
Балкашина Е. И., Ромашов Д. Д. Генетическое строение популяций Drosophila // Биол. журн. 1935. Т. 4, № 1. С. 81–106.
Бауэр Э. С. Теоретическая биология. М.; Л.: ВИЭМ, 1935. 205 с.
Белоусов Л. В. Истоки, развитие и перспективы теории биологического поля // Физические и химические основы жизненных явлений. М.: Изд-во АН СССР, 1963. С. 59–117.
Белоусов Л. В. Целостные и структурно-динамические подходы к онтогенезу // Журн. общ. биологии. 1979. Т. 40, № 4. С. 514–529.
Белоусов Л. В., Чернавский Д. С. Неустойчивость и устойчивость в биологическом морфогенезе // Онтогенез. 1977. Т. 8, № 2. С. 99–114.
Блюменфельд Л. А. Физические аспекты биологической эволюции // Философия в современном мире. Философия и теория эволюции. М.: Наука, 1974. С. 56–71.
Бляхер Л. Я., Воронцова М. А., Лиознер Л. Д. Каузально-аналитический метод в учении об индивидуальном развитии // Тр. Ин-та эксперим. морфогенеза. 1935. Т. 3. С. 223–239.
Волькенштейн М. В. Физический смысл нейтралистской гипотезы эволюции // Журн. общ. биологии. 1981а. Т. 42, № 5. С. 680–686.
Волькенштейн М. В. Биофизика. М.: Наука, 1981б. 575 с.
Галл Я. М. И. И. Шмальгаузен и проблема факторов эволюции // Историко-биологические исследования. М.: Наука. 1980. Вып. 8. С. 106–123.
Гершензон С. М. "Мобилизационный резерв" внутривидовой изменчивости // Журн. общ. биол. 1941. Т. 2, № 1. С. 85–107.
Гершензон С. М. Роль естественного отбора в распространении и динамике меланизма у хомяков (Cricetus cricetus L.) //Журн. общ. биол. 1946. Т. 7, № 2. С. 97–127.
Гершензон С. М. Молекулярная биология и теория эволюции // Философия в современном мире. Философия и теория эволюции. М.: Наука, 1974. С. 72–89.
Гершковчч И. Генетика. М.: Наука, 1968. 687 с.
Голубовский М. Д. Некоторые аспекты взаимодействия генетики и теории эволюции / Методологические и философские проблемы биологии. Новосибирск: Наука, 1981. С. 69–92.
Голубовский М. Д. Критические исследования в области генетики // Александр Александрович Любищев. Л.: Наука, 1982. С. 52–65.
Голубовский М. Д. Организация генотипа и формы наследственной изменчивости у эукариот // Методологические проблемы медицины и биологии. Новосибирск: Наука. 1985. С. 135–152.
Голубовский М. Д., Иванов Ю. Н., Захаров И. К., Берг Р. Л. Исследование синхронных и параллельных изменений генофондов в природных популяциях плодовых мух Drosophila melanogaster // Генетика. 1974. Т. 10, № 4. с. 73–81.
Гольдшмидт Р. Генетика и физиология развития // Природа. 1933. № 56. С. 124–133.
Грант В. Эволюция организмов. М.: Мир, 1980. 407 с.
Гурвич А. Г. Теория биологического поля. М.: Сов. наука, 1944. 156 с.
Дарвин Ч. Изменения домашних животных и культурных растений // Сочинения. М.; Л.: Изд-во АН СССР, 1951. Т. 4. 883 с.
Дарвин Ч. Происхождение видов путем естественного отбора. М.: Сельхозгиз, 1952. 484 с.
Дриш Г. Витализм: его история и система. М.: Наука, 1915. 279 с.
Дубинин Н. П. Эволюция популяций и радиация. М.: Атомиздат, 1966а. 743 с.
Дубинин Н. П. Основы популяционной генетики // Актуальные вопросы современной генетики. М.: Изд-во МГУ, 19666. С. 221–265.
Дубинин Н. П. О сущности явления наследственности//Журн. общ. биологии. 1973. Т. 34, № 1. С. 3–12.
Дубинин. Н. П. Общая генетика. М.: Наука, 1976. 590 с.
Дубинин Н. П., Ромашов Д. Д., Гептнер М. А., Демидова 3. А. Аберративный полиморфизм у Drosophila fasciata Meig. (syn.— melanogaster Meig.) // Биол. журн. 1937. Т. 6, № 2. С. 311–354.
Дэвидсон Э. Действие генов в раннем развитии. М.: Мир, 1972. 342 с.
Инге-Вечтомов С. Г. Система генотипа. Физиологическая генетика. Л.: Медицина, 1976. С. 57–113.
Инге-Вечтомов С. Г., Сойдла Т. Р. Эволюционные аспекты проблемы доминирования и молекулярные взаимодействия // Итоги науки и техники. Общая генетика. М., 1978. Т. 3: Эволюция и популяционная генетика. С. 7–37.
Иогансен В. Элементы точного изучения наследственности и изменчивости. М.; Л.: Сельхозгиз, 1933. 410 с.
Камшилов М. М. Генотип как целое // Успехи совр. биологии. 1934. Т. 3, вып. 2. С. 181–207.
Камшилов М. М. Является ли плейотропия свойством гена? // Биол. журн. 1935. Т. 4, № 1. С. 113–144.
Камшилов М. М. Отбор как фактор, меняющий зависимость признака от изменения внешних условий//Докл. АН СССР. 1939. Т. 23, № 4. С. 361–364.
Камшилов М. М. Изменчивость и проявление. Проблема нормального фенотипа // Докл. АН СССР. 1940. Т. 29, № 3. С. 239.
Камшилов М. М. К вопросу об отборе на холодоустойчивость // Журн. общ. биологии. 1941. Т. 2, № 2, С. 211–227.
Камшилов М. М. Отбор как фактор усложнения организации // Изв. АН СССР. Сер. биол. 1948. № 3. С. 349–356.
Камшилов М. М. Роль фенотипа в эволюции // Генетика. 1967. № 12. С. 108–116.
Камшилов М. М. Фенотип и генотип в эволюции // Пробл. эволюции. 1972. Т. 4. С. 28–44.
Камшилов М. М. Эволюция биосферы. М.: Наука, 1979. 256 с.
Кастлер Г. Возникновение биологической организации. М.: Мир, 1967. 90 с.
Кирпичников В. С. У истоков теории стабилизирующего отбора // История и теория эволюционного учения. Л., 1974. Вып. 2. С. 61–67.
Кожанчиков И В. Об условиях возникновения биологических форм // Тр. Зоол. ин-та АН СССР. 1941. Т. 6, № 4. С. 16–32.
Конюхов Б. В., Нончев С. Г. Экспрессия доминантных и рецессивных признаков в онтогенезе млекопитающих // Журн. общ. биологии. 1981. Т. 42, № 3. С. 325–334.
Лобашев М. Е. Физиологическая (паранекротическая) гипотеза мутационного процесса // Вестн. ЛГУ. 1947. Вып. 8. С. 10–29.
Лобашев М. Е. Генетика. Л.: Изд-во ЛГУ, 1963. 484 с.
Лобашев М. Е. Принципы генетического анализа // Актуальные вопросы современной генетики. М.: Изд-во МГУ, 1966. С. 7–22.
Майр Э. Зоологический вид и эволюция. М.: Мир. 1968. 597 с.
Майр Э. Популяции, виды и эволюция. М.: Мир, 1974. 460 с.
Майр Э. Эволюция. М.: Мир, 1981. С. 11–32.
Мейнард-Смит Дж. Эволюция полового размножения. М.: Мир, 1981. 271 с.
Мейстер Г. К. Критический анализ основных понятий генетики. М.; Л.: Сельхозгиз, 1934. 202 с.
Михайлова Н. Н., Симаров Б. В., Тихомирова В. Л., Инге-Вечтомов С. Г. Действие повышенной концентрации ионов магния и натрия на трансляционном и посттрансляционном уровнях у дрожжей Sacharomyces cerevisiae // Исследования по генетике. Л.: Изд-во ЛГУ, 1981. № 9. С. 65–76.
Морган Т. Г. Структурные основы наследственности. М.; Л.: Госиздат, 1924. 310 с.
Морган Т. Механизм наследственности, устанавливаемый на основании наследования сцепленных признаков // Избранные работы по генетике. М.; Л.: Сельхозгиз, 1937а. С. 125–135.
Морган Т. О механизме наследственности // Избранные работы по генетике. М.; Л.: Сельхозгиз, 19376. С. 189–224.
Морган Т. Возникают ли рецессивные мутации путем утраты генов // Избранные работы по генетике. М.; Л.: Сельхозгиз, 1937в. С. 226–242.
Морган Т. Значение генетики для физиологии и медицины (Нобелевская лекция) // Избранные работы по генетике. М.; Л.: 1937. С. 256–271.
Морган Т. Г. Развитие и наследственность. М.; Л.: Биомедгиз, 1937д. 242с.
Мюнтцинг А. Генетика. М.: Мир, 1967. 610 с.
Новинский Ч. Теория эволюции как теория процесса самоорганизации // Проблемы взаимосвязи организации и эволюции в биологии. М.: Наука, 1978. С. 72–102.
Оленов Ю. М. Проблемы молекулярной генетики. Л.: Наука, 1977. 207 с.
Пеннет Р. Менделизм. М.; Л.: Госиздат, 1930. 240 с.
Промптов А. Н. Ген и признак в онтогенезе // Успехи соврем, биологии. 1934. Т. 3. вып. 2. С. 145–180.
Рапопорт И. А. О законе распространения и погашения генного действия // Докл. АН СССР. 1941. Т. 36, № 4. С. 392–395.
Рапопорт И. А. Генетический анализ зависимой дифференциации в эмбриогенезе двукрылых // Изв. АН СССР. Сер. биол. 1943. № 2. С. 80–92.
Рьюз М. Философия биологии. М.: Прогресс, 1977. 320 с.
Самохвалова Г. В. Получение наследственных изменении у тлей при перемене кормовых растений //Жури. общ. биол. 1951. Т. 12, № 3. С. 176–191.
Самохвалова Г. В. Получение направленных наследственных изменений у тлей при перемене кормовых растений. Сообщ. второе // Зоол. журн. 1954. Т. 33, № 5. С. 1032–1040.
Светлов П. Г. Теория критических периодов развития и ее значение для понимания принципов действия среды на онтогенез // Вопросы цитологии и общей физиологии. М.; Л.: Изд-во АН СССР, 1960. С. 263–285.
Светлов П. Г. О целостном и элементаристическом методах в эмбриологии // Арх. анатомии, гистологии и эмбриологии. 1964. Т. 46, № 4. С. 3–26.
Светлов П. Г. Онтогенез как целенаправленный (телеономический) процесс // Арх. анатомии, гистологии и эмбриологии, 1972. Т. 63, № 8. С. 5–16.
Светлов П. Г. Физиология (механика) развития. Л.: Наука, 1978. Т. 1. 279 с.; Т. 2. 262 с.
Сойдла Т. Р. Особенности генов эукариот // Инге-Вечтомов С. Г. Введение в молекулярную генетику. М.: Высш. шк., 1983.
Солбриг О., Солбриг Д. Популяционная биология и эволюция. М.: Мир, 1982. 488 с.
Спенсер Г. Основания биологии. СПб.: Изд. Иогансона, 1899. Т. 1. 456 с.; Т. 2. 380 с.
Стент Г., Кэлиндар Р. Молекулярная генетика. М.: Мир, 1981, 646 с.
Столетов В. Н. Предисловие к русскому изданию // Мюнтцинг А. Генетика. М.: Мир, 1967. С. 5–9.
Тимофеев-Ресовский Н. В., Иванов В. И. Некоторые вопросы феногенетики // Актуальные вопросы современной генетики. М.: Изд-во МГУ, 1966. С. 114–130.
Тимофеев-Ресовский Н. В., Тимофеева-Ресовская Е. А., Циммерман И. К. Экспериментально-систематический анализ географической изменчивости и формообразования у Epilachna chrysomelina F. (Coleoptera, Coccinelidae) // Тр. Ин-та биологии УФАН. 1965. Вып. 44. С. 27–63.
Уоддингтон К. Канализация развития и наследование приобретенных признаков // Успехи совр. биологии. 1944. Т. 18, вып. 3. С. 393–396.
Уоддингтон К. Организаторы и гены. М.: Изд-во иностр. лит., 1947. 240 с.
Уоддингтон К. Дифференцировка клеток // Физика и химия жизни. М.: Изд-во иностр. лит., 1960. С. 215–226.
Уоддингтон К. Морфогенез и генетика. М.: Мир, 1964. 259 с.
Уоддингтон К. X. Основные биологические концепции // На пути к теоретической биологии. I. Пролегомены. М.: Мир, 1970а. С. 11–38.
Уоддингтон К. X. Зависит ли эволюция от случайного поиска? // На пути к теоретической биологии. I. Пролегомены. М.: Мир, 1970б. С. 108–115.
Филипченко Ю. А. Наследственность. М.: Пг.: Госиздат, 1924. 252 с.
Филипченко Ю. А. Эволюционная идея в биологии. М.: Наука, 1977. 227 с.
Харланд С. К. Генетическая концепция вида // Успехи соврем, биологии. 1937. Т. 6. вып. 3. С. 512–543.
Четвериков С. С. О некоторых моментах эволюционного процесса с точки зрения современной генетики // Журн. эксперим. биологии. Сер. А. 1926. № 2. С. 3–54.
Шапошников Г. X. Специфичность и возникновение адаптации к новым хозяевам у тлей (Нomоptera, Aphidoidea) в процессе естественного отбора (экспериментальные исследования) // Энтомол. обозр., 1961. Т. 40, № 4. С. 739–762.
Шапошников Г. X. Морфологическая дивергенция и конвергенция в эксперименте с тлями (Ноmoptera, Aphidenea) // Энтомол. обозр., 1965. Т. 44. С. 3–25.
Шапошников Г. X. Возникновение и утрата репродуктивной изоляции и критерий вида // Энтомол. обозр. 1966. Т. 45, № 1. С. 3–35.
Шапошников Г. X. Динамика клонов, популяций и видов и эволюция // Журн. общ. биологии. 1978. Т. 39, № 1. С. 15–33.
Шварц С. С. Принцип оптимального фенотипа (к теории стабилизирующего отбора) // Журн. общ. биологии. 1968. Т. 29, № 1. С. 12–24.
Шеппард Ф. М. Естественный отбор и наследственность. М.: Просвещение, 1970. 216 с.
Шишкин М. А. Закономерности эволюции онтогенеза // Журн. общ. биологии. 1981. Т. 42. № 1. С. 38–54.
Шишкин М. А. Индивидуальное развитие и естественный отбор // Онтогенез. 1984а. Т. 15, № 2. С. 115–136.
Шишкин М. А. Фенотипические реакции и эволюционный процесс // Экология и эволюционная теория. Л.: Наука, 1984б. С. 196–216.
Шишкин М. А. Менделевский фактор как свойство эпигенетической системы // Макроэволюция: Материалы I Всесоюз. конф. по проблемам эволюции. М.: Наука, 1984в. С. 238–239.
Шмальгаузен И. И. Изменчивость и смена адаптивных норм в процессе эволюции // Журн. общ. биологии. 1940а. Т. 1, № 4. С. 509–528.
Шмальгаузен И. И. Пути и закономерности эволюционного процесса. М.; Л.: Изд-во АН СССР, 19406. 231 с.
Шмальгаузен И. И. Стабилизирующий отбор и его место среди факторов эволюции // Журн. общ. биологии. 1941. Т. 2, № 3. С. 307–354.
Шмальгаузен И. И. Регуляция формообразования в индивидуальном развитии. М.: Изд-во АН СССР. 1964. 132 с.
Шмальгаузен. И. И. Кибернетические вопросы биологии. Новосибирск: Наука, 1968а. 223 с.
Шмальгаузен И. И. Факторы эволюции: Теория стабилизирующего отбора. М.: Наука, 1968б. 451 с.
Шмальгаузен И. И. Проблемы дарвинизма. Л.: Наука, 1969. 492 с.
Шмальгаузен И. И. Организм как целое в индивидуальном и историческом развитии. М.: Наука, 1982. 228 с.
Шпеманн Г. Наследственность и механика развития // Журн. эксперим. биологии. Сер. Б. 1925. Т. I, вып. 34. С. 119–144.
Эшби У. Р. Введение в кибернетику. М.: Изд-во иностр. лит., 1959. 432 с.
Эшби У. Р. Конструкция мозга. М.: Изд-во иностр. лит., 1962. 398 с.
Яблоков А. В. История биологии и концепция естественного отбора: [Предисловие] \\ Рубайлова Н. Г. Формирование и развитие теории естественного отбора. М.: Наука, 1981. С. 3–7.
Alberch P. Ontogenesis and morphological diversification // Amer. Zool. 1980. Vol. 20. P. 653–667.
Alberch P. Developmental constraints in evolutionary processes // Life Sci. Res. Rep. 1982. N 22. P. 313–332.
Baer К. Ueber Entwicklungsgeschichte der Thiere. T. I. Koenigsberg: Kupfertaf, 1828. 271 S.
Baur E. Einfuhrung in die experimentelle Vererbungslehre. В.: Borntraeger, 1922. 436 S.
Beer G. de. Charles Darvin: Evolution by Natural Selection. L.: Nelson, 1963. 290 p.
Bertalanffy L. Das biologische Weltbild. Bern: Francke, 1949. 202 S.
Bertalanffy L. General System Theory. N. Y.: Braziller, 1969. 289 p.
Bridges С. The origin of variations in sexual and sex-limited characters // Amer. Natur. 1922. Vol. 56. P. 51–63.
Castle W. E. Can selection cause genetic change? // Amer. Nat. 1916. Vol. 50. N 592. P 248–256.
Dobzhansky Th. Genetic and the Origin of Species, 3d ed. N. Y.: Columbia Univ. press. 1947. 446 p.
Driesch Н. The Science and Philosophy of the Organism. L.: Black, 1908. Vol. 1. 329 p.; Vol. 2. 381 р.
Fisher R. Genetical Theory of Natural Selection. Oxford: Clarendon press, 1930. 272 p.
Goldschmidt R. Physiologische Theorie der Vererbung. В.: Springer, 1927. 247 S.
Goldschmidt R. Physiological Genetics. N. Y.; L. McGraw Hill Book Co., 1938. 375 p.
Goldschmidt R. The Material Basis of Evolution. New Haven: Yale Univ. press, 1940. 436 p.
Goldschmidt R. Mimetic polymorphism, a controversial chapter of Darwinism // Quart. Rev. Biol. 1945. Vol. 20, N 3. P. 205–230.
Goldschmidt R. Theoretical Genetics. Los Angeles: Univ. Cal. press, 1955. 563 p.
Goodwin В. С. Biological stability // Towards a Theoret. Biology / Ed. C. Waddington. Chicago: Aldine, 1970. P. 1–17.
Gurwitsch A. Ueber Determination, Normierung und Zufall in der Ontogenese // Roux' Arch. 1910. Bd. 30. S. 133–193.
Gurwitsch A. Die Vererbung als Verwirklichungsvorgang // Biol. Zentr.-BI. 1912. Bd. 32. S. 458–486.
Gurwitsch A. Ueber den Begriff des embryonalen Eeldes // Roux' Arch. 1922. Bd. 51, h. 3/4. S. 383–415.
Haldane S. The Biochemistry of Genetics. L.: Alien and Unwin, 1954. 144 p.
Но М. W., Sounders Р. Т. Beyond neo-Darvinism — an epigenetic approach to evolution // J: Theor. Biol. 1979. Vol. 78, N 4. P. 573–591.
Johannsen W. Elemente der exakten Erblichkeitslehre. Aufl. 3. Jena: Fischer, 1926. 736 S.
Kettlewell H. B. D. A. resume on investigations on the evolution of melanism in the Lepidoptera // Proc. Roy. Soc. London B. 1956. Vol. 145, N 920. P. 297–303.
Kimura М. Genetic variability maintained in a finite population due to mutational production of neutraly or nearly neutral isoalleles // Genet. Res. 1968. Vol. 2. P. 247–269.
Kojima K. 1. Is there a constant fitness value for a given genotype? // Evolution. 1971. Vol. 25. P. 281–285.
Lewontin R. C. On the irrelevance of genes // Towards a Theoretical Biology // Ed. C. Waddington. Chicago: Aldine, 1970. P. 67–88.
Maderson P. F. A. et al. The role of development in macroevolutionary change. Croup report // Life Sci. Res. Rep. 1982. N 22- P. 279–312.
Roux W. Gesammelte Abhandlungen uber Entwicklungsmechanik der Organismen. Leipzig: Engelmann, 1895. Bd. 1. 816 S.; Bd. 2. 1075 S.
Simpson G. G. Major Features of Evolution. N. Y.: Columbia Univ. press. 1953. 434 p.
Standfuss М. Zur Frage des Gestaltung und Vererbung auf Grund 28-jahriger Experimente // Insektenborse. 1902. Bd. 19. S. 155–169.
Waddington C. H. Genetic assimilation of an acquired characters // Evolution. 1953. Vol. 7. P. 118–126.
Waddington С. Н. The Strategy of the Genes: a Discussion on Some Aspects of Theoretical Biology. L.: Alien and Unwin, 1957. 262 p.
Waddington С. Н. Principles of Development and Differentiation. N. Y.: Macmillan, 1976. 115 p.
Wake D. В., Roth G., Wake M. Н. On the problem of stasis in organismat evolution // J. Theor. Biol. 1983. Vol. 101. P. 211–224.
Wolpert L. Mechanisms of limb development and malformation // Brit. Med. Bull. 1976. Vol. 32, N 1. P. 65–70.
Woodger I. Н. What do we mean by «inborn»? // Brit. J. Philos. Sci. 1953. Vol. 3. P. 319.
Wright S. An intensive study of the inheritance of color // Carnegie Inst. Wash. Publ. 1916. Vol. 241, pt. 2. P. 59–160.
Wright S. Evolution organic// Encyclopaedia Britanica. L., 1964. Vol. 8. P. 915–931.
Zimmermann K. Ober Mutationen in wilden Populationen // Mitt. Zool. Mus. Berlin. 1933. Bd. 19. S. 439–452.