Про природу чоловічого та жіночого, або від кон'югації до оогамії. Колонка для Комп'ютерри #138
Чому я витратив так багато місця на обговорення випадку інфузорій? Він показує, що розмноження без утворення специфічних клітин можуть використовувати надзвичайно високоорганізовані організми. Більше того, на їхньому прикладі простежується зв’язок між статевим процесом і феноменом смерті. Повірте...
←
Dmytro Shabanov
→
Обговорення переходу від гермафродитизму до роздельнополості як приклад некласичного розвитку природничої гіпотези
Про природу чоловічого і жіночого, або від кон'югації до оогамії
Ненаписана колонка. Про насильство: проста етична задача і розбір трьох прикладів із сучасного українського життя
Колонка для Комп'ютерри #137
Колонка для Комп'ютерри #138
Я продовжую серію «статевих» колонок, точніше — колонок про еволюцію рекомбінації. Це — вже восьма за рахунком, а от — перша). Після того як я пояснив, чому припускаю, що статеве розмноження могло виникати у гермафродитів, хочу обговорити теорію, яка створює серйозні проблеми для таких уявлень. Мене питали, чому я «молчав» (не писав нові колонки) два тижні. Не міг у голові уявити, як те, що я викладу тут, поєднується з тим, про що писав раніше. Питання для мене залишається відкритим. Рішення, яке я знайшов, таке: тут я викладу суттєві для мене факти, які зроблять для читачів більш зрозумілим різноманіття форм статевого процесу, а з головними висновками, які я намагаюся сформулювати, повішаю.
Я почну з істин із розряду тих, що викладаються в підручниках. Статевий процес пов’язаний із заплідненням — злиттям спадкового матеріалу двох клітин. Як я писав у другій колонці з «статевої серії», у типічному випадку запліднення складається з двох етапів — злиття двох клітин (сингамії) і злиття їх ядер (кариогамії). Що це за клітини, які зливаються? Це можуть бути неспеціалізовані клітини, що становлять частину тіла або все тіло одноклітинного організму, — і такий випадок можна назвати соматогамією. Найчастіше поняття «соматогамія» застосовують лише до грибів, у яких можуть з’єднуватися клітини з різних гіф, але в ширшому сенсі його можна застосувати і до багатьох інших випадків, коли для запліднення використовуються клітини, які виконують і інші функції. У цій, широкій трактовці найпоширенішим типом соматогамії є кон'югація. При ній клітини двох організмів (одноклітинних або багатоклітинних) з’єднуються мостиками, через які передаються мігруючі ядра (іноді — разом із усім клітинним вмістом). При кон'югації інформація може передаватися в обидва боки (наприклад, у інфузорії туфельки) або лише в один (наприклад, у водорості спирогири). Природно, перший випадок аналогічний перехресному гермафродитизму, а другий — роздельнополості.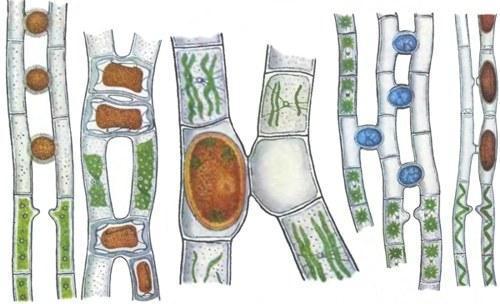
Так відбувається кон'югація у деяких водоростей (рисунок із класичного довідника «Життя рослин»)
Кон'югація (і інші форми соматогамії у широкому розумінні слова) здається вам примітивним, архаїчним рішенням? Дозвольте розповісти, як вона відбувається у найскладніших за будовою клітинах, створених земною еволюцією. Я говорю про найвищоорганізовані одноклітинні — про інфузорії, такі як «шкільна» інфузорія туфелька.
Розмножуються (діляться) інфузорії, перетинаючи свою довгу вісь. А при кон'югації дві інфузорії притискаються одна до одної довгими сторонами.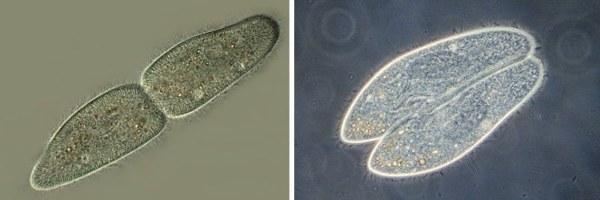
Розмноження (зліва) і кон'югація (справа) у інфузорії туфельки
Розмноження відбувається без статевого процесу, а статевий процес — без розмноження. Інша справа, що після статевого процесу інфузорії стають, по суті, новими індивідами, які далі переходять до ділення — клонального розмноження. Як би це не здивувало, клон, що складається з багатьох поколінь клітин, проходить стадії незрілості, зрілості, зрілості і старості. Після завершення періоду незрілості (протягом якого змінюється певна кількість поколінь клітин, що розмножуються діленням; у туфельки він триває близько місяця) клон виявляється здатним до кон'югації. Вона можлива з генетично іншими клонами, які належать до іншого типу спарювання (якомусь аналогові статі). Кількість типів спарювання у різних інфузорій може бути різною: 2, 4, 6, 7, 8, 10...
Толчком для переходу до статевого процесу може стати погіршення (з точки зору інфузорії) умов існування. Поки все йде добре, інфузорії діляться, зберігаючи відповідний (раз для його власника все йде добре) умовам життя генотип. Однак якщо інфузорія ділилася сотні раз і при цьому не проходила через статеве «оновлення», то з часом вона втрачає здатність до подальших поділів і гине.
Уявіть: у культурі росте і розвивається старіючий клон інфузорій. Оскільки всі індивіди належать до одного клону, вони мають один тип спарювання. Починаючи з певного моменту вся експериментальна популяція виявляється приреченою: її особи не можуть знайти партнерів для статевого процесу. Клон постарів: інфузорії живуть, діляться, але при будь‑якому розвитку подій через певну кількість поколінь їх розмноження зупиниться… Парадокс? Безумовно. Тут проявляється якийсь механізм, що не дозволяє інфузоріям відмовитися від періодичного статевого процесу.
Клітина інфузорії дуже складна, і необхідні для її життєдіяльності гени багаторазово копіюються в одному з двох ядер інфузорії — великому ядрі, макронуклеусі. Перед кон'югацією макронуклеус руйнується, а микронуклеус, мале ядро, проходить мейоз, ділячись на чотири унікальних ядра з зменшеною вдвічі кількістю генетичної інформації. Три з чотирьох ядер гинуть, а одне ділиться ще раз за допомогою мітозу, утворюючи стаціонарне («жіноче») і мігруюче («чоловіче») ядра. Дві інфузорії обмінюються мігруючими ядрами, які зливаються зі стаціонарними ядрами партнера‑інфузорії, утворюючи нові микронуклеуси. У результаті отримуються два генетично ідентичних індивіда‑екскон'югати…
Чому я витратив стільки місця на обговорення випадку інфузорій? Він показує, що розмноження без утворення специфічних клітин можуть використовувати надзвичайно високоорганізовані істоти. Більше того, на їхньому прикладі простежується зв’язок між статевим процесом і феноменом смерті. Повірте, хоча цей зв’язок недостатньо усвідомлений, він дуже глибокий. Не витрачаючи поки часу на його обґрунтування, висловлю важливу думку: причини, які забезпечують статевий процес, тісно пов’язані з причинами, що забезпечують запрограмовану смерть індивідів...
Перейдемо до другого, набагато поширенішого типу статевого процесу — з використанням спеціалізованих статевих клітин, гамет. Цей тип статевого процесу називається гаметогамією. Гамети можуть продукувати як одноклітинні, так і багатоклітинні організми. Відомі три основні форми гаметогамії. При ізогамії гамети, які зливаються один з одним, мають однаковий розмір. Це підвижні клітини, які інтенсивно шукають один одного. Ці клітини можуть бути зовні зовсім однаковими, але при цьому вони все ж несуть біохімічні маркери, що забезпечують злиття (сингамію) різних клітин. Про стать тут говорити не треба, різні типи гамет можна позначити, наприклад, знаками «+» і «—». При анізогамії підвижні гамети належать до двох типів — великого і малого. У цьому випадку зрозуміло, що великі гамети відповідають жіночій статі, а малі — чоловічій. Логічним завершенням цієї еволюції є оогамія. Жіноча гамета втрачає активну підвижність і називається яйцеклітиною; її пошук — завдання малих і підвижних гамет, які називаються сперматозоїдами.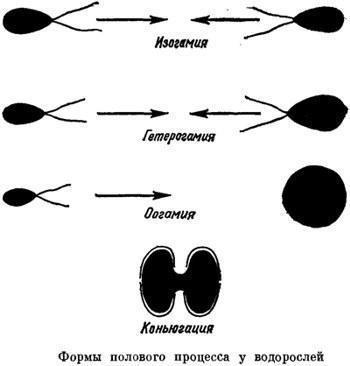
Порівняння трьох форм гаметогамії та кон'югації («Життя рослин»)
Ймовірно, найпростішим варіантом гаметогамії є ізогамія. Як здається, цей спосіб розмноження є най«логічнішим» і простим. Чому ж тоді у більшості високоорганізованих видів ми спостерігаємо оогамію?
Згадайте колонку, у якій я розповідав про можливий механізм переходу від гермафродитизму до роздельнополості. Успішність того чи іншого рішення залежить не від його оптимальності, а від його стійкості (або нестійкості). Ізогамія виявляється нестійкою.
Можливість такого механізму переходу від ізогамії до анізогамії, про який я буду говорити, усвідомлена вже давно. Перша публікація про це датується 1972 роком (Parker, G. A., Baker, R. R. and Smith, V. G. F. The origin and evolution of gamete dimorphism and the male-female phenomenon. Journal of Theoretical Biology, 1972, v. 36, p. 529–553). Цю концепцію прийнято називати PBS‑теорією, за першими літерами прізвищ трьох її авторів. З часу створення цієї теорії моделювання взаємодії гамет різного розміру перетворилося на окрему галузь науки.
Опишу основну ідею, користуючись своєю моделлю, значно примітивнішою, ніж найкращі зразки, створені для вивчення цієї проблеми. Модель можна завантажити з мого сайту: або версію для Excel‑2013, з якою я працював, або трохи спрощений варіант для Excel‑2003.
Розглянемо популяцію організмів, які викидають гамети у воду. Початковий стан — типова ізогамія. Організми, що належать до обох типів спарювання (1 і 2), виробляють однакові за розміром гамети. У випадку, показаному на рисунку, розмір усіх гамет становить 12,5 % від максимально можливого.
Гамети зустрічаються в товщі води, зливаються і утворюють зиготу. З імовірністю 50 % особина належить до першого або другого типу. Вона буде виробляти такі ж за розміром гамети, які виробляв її батько, що належить до її типу спарювання, з невеликим уточненням. З невеликою імовірністю d (у показаному прикладі d=0,005) особина переходить до виробництва гамет сусіднього розмірного класу: в 2 рази більші або в 2 рази менші. Усі особи вкладають у виробництво гамет одну й ту ж кількість енергії, і тому ті особи, які виробляють вдвічі менші гамети, викидають у воду в 2 рази більше їх кількість.
Останнє, що треба пояснити перед обговоренням результатів, — це те, що імовірність виживання зигот залежить від кількості енергії, отриманої ними від гамет (просто від розміру). Найпростіший варіант залежності — пряма пропорційність розміру зиготи і її шансів на виживання. Подивимось, що вийде.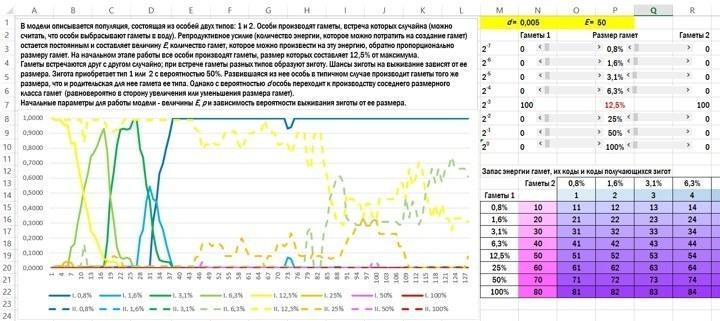
Вікно з коротким описом моделі, клітинками для вводу початкових параметрів і підсумковим графіком її роботи. Гамети першого «пола» показані суцільною лінією, другого — пунктирною. Виживаність зигот прямо пропорційна їх енергетичному запасу. У початковому стані обидва типи організмів виробляють гамети в 12,5 % від максимуму; до кінця імітації перший тип виробляє виключно гамети розміром 1,6 %, а другий — в основному 6,3 % від максимуму.
Обидва «пола» зменшують розмір своїх гамет. Чому — легко зрозуміти. При постійності розміру статевих клітин партнера та особина, яка зменшить свої гамети вдвічі, удвічі збільшить кількість своїх нащадків, а їх виживаність скоротиться лише на 75 % (запас енергії зиготи залежить від розміру обох гамет). У цій ситуації обидва «пола» скорочують розмір своїх статевих клітин. Це триває до межі, при якій виживаність зигот стає загрозливо низькою.
Чи реалістичні використані мною умови? Не зовсім. Ймовірно, зигота, яка менша за певний мінімальний розмір, взагалі не має шансів на виживання. Життєздатність зиготи в такому випадку має бути пропорційна різниці між її розміром і цим мінімумом.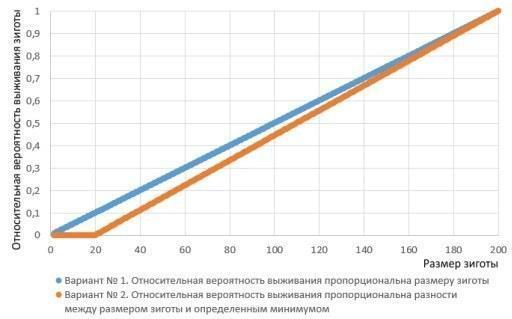
Два варіанти залежності шансів на виживання зиготи від її розміру (зумовленого розміром гамет)
Слід перейти з першого варіанту залежності шансів на виживання зиготи до другого, поведінка моделі кардинально змінюється. Збільшення розмірів гамет також виявляється вигідною стратегією (зверніть увагу: події на картинці внизу починаються з того, що «пол», показаний пунктиром, переходить до виробництва більших статевих клітин).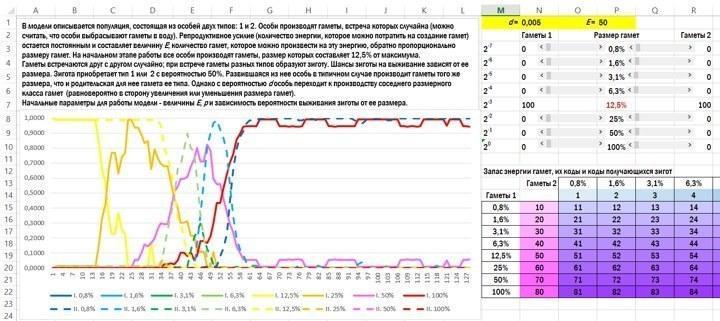
Тут реалізовано другий варіант залежності шансів на виживання зиготи від її розміру. На початку імітації обидва «пола» виробляють гамети в 12,5 % від максимуму; до кінця (через 128 поколінь) перший «пол» дає майже виключно гамети в 0,8 %, а другий — у 100 % від максимального розміру.
За цих умов модель швидко переходить до стану, де один «пол» виробляє найбільші з можливих гамет, а інший — найменші. Який саме «пол» виявиться «великогаметним», а який — «малогаметним», вирішує випадок.
Як бачите, у нашій моделі ми спостерігали перехід від ізогамії до анізогамії.
Як я вже сказав, для вивчення цього переходу розроблені досить складні моделі, які враховують і інші фактори. Наприклад, очевидно, що імовірність зустрічі гамет залежить від ефективності їх плавання, яка, у свою чергу, залежить від їх розмірів (зокрема, впливу на число Рейнольдса — величини, що визначає характер руху обтікальної гами течії). Урахування таких факторів призводить до складніших залежностей, які, проте, підтверджують загальну закономірність: ізогамія відносно нестійка і з великою ймовірністю витісняється анізогамією.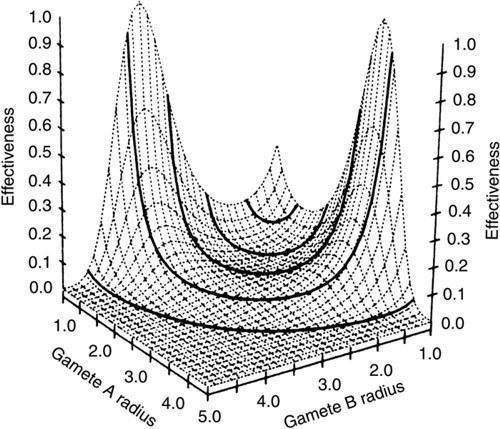
Тривимірна візуалізація результатів значно складнішого моделювання. Висота піків відображає стійкість різних комбінацій розмірів гамет. Ізогамії відповідає невеликий пік посередині, а анізогамії — два симетрично розташованих вищих піка по краях (з книги: T. Togashi and P. A. Cox (editors). The Evolution of Anisogamy. Cambridge University Press, 2011, 250 p.)
І знаєте, що ми в кінці‑кінців отримали? Роздельнополість! Справа в тому, що самці — це «пол», який виробляє багато малих статевих клітин, а самки — «пол», який виробляє небагато великих. Усі інші невідповідності між самками і самцями є наслідками цього первинного різницевого розподілу!
Тепер, як мені здається, уважним читачам восьми «статевих» колонок надано ключові факти і припущення, на основі яких можна судити про можливі сценарії виникнення статевого розмноження. Цим питанням ми займемося наступного разу.
←
Dmytro Shabanov
→
Обговорення переходу від гермафродитизму до роздельнополості як приклад некласичного розвитку природничої гіпотези
Про природу чоловічого і жіночого, або від кон'югації до оогамії
Ненаписана колонка. Про насильство: проста етична задача і розбір трьох прикладів із сучасного українського життя
Колонка для Комп'ютерри #137
Колонка для Комп'ютерри #138