Українська мова (найновіша версія) / Русский язык (обновление прекращено)
3.08. Сукцессии. Основные понятия
Приходилось ли вам оказываться на заброшенном поле или в покинутом саду? Вы, наверное, догадываетесь, что по их внешнему виду можно установить, когда за ними прекратили уход. Благодаря чему это становится возможным? Предоставленная сама себе экосистема (в данных примерах — искусственная сельскохозяйственная экосистема — агросистема) начинает изменяться. Зная скорость таких изменений, можно оценить, в течение какого времени оно происходило.
А теперь представьте себе реку, текущую по широкой долине. Русло такой реки образует петли — меандр. Там, где русло делает поворот, текущая вода подмывает берег, в который она упирается, и формирует береговой обрыв. А на противоположном крае такого берега разрастается песчаный пляж. У самой воды песок голый, чуть дальше — порос травой, еще дальше — трава образует плотную дерновину, и, наконец, на каком-то расстоянии от берега растут кустарники. Легко понять, что этот переход от одного сообщества к другому (экоклина) отражает динамику изменений каждого отдельного участка берега во времени. Там, где сейчас растет кустарник, когда-то только намывался пляж, образующий новый участок берега… Петли реки растут и со временем соединяются друг с другом. Русло прокладывает себе более короткую дорогу, а его прежние участки образуют старицы. Со временем старицы превращаются в пойменные озера, наполняются осадками и становятся частями пойменного луга. Приведенные нами примеры показывают способность экосистем к изменениям во времени — сукцессиям (от лат. successio — преемственность, наследование).
Сукцессия — это последовательная смена сообществ в одном местообитании. Это направленный, контролируемый сообществом процесс, ведущий к определенному состоянию. Сукцессия в биогеоценозах является более длительным процессом, чем сезонные изменения, но и не столь длительным, как эволюция экосистем. Экоклина, таким образом, является сукцессией, наблюдаемой в пространстве.
Обычно в ходе сукцессии можно выделить преходящие стадии, которые называются серийными сообществами (или попросту сериями), и окончательное устойчивое состояние — климаксное сообщество (климакс). Учение о сукцессиях разработано в 20-е годы XX века Фредериком Клементсом, американским экологом, который рассматривал сукцессию как некий аналог онтогенеза экосистемы.
Причиной сукцессий является не только изменение местообитаний какими-то внешними факторами (например, текущей водой, как в описанном случае с меандром реки), но и вся совокупность взаимодействий между компонентами сообществ, а иногда — и воздействия со стороны человека.
«Характерна динамика сообществ, описанная для конца XIX — начала XX веков в вересковых торфяниках Шотландии. Хозяин этих земель использовал их для охоты на многочисленных здесь шотландских куропаток — граусов. В 1892–1893 гг. на верещатнике поселилось несколько пар чаек. Взятые хозяином под охрану, они в начале нашего столетия уже образовали большую колонию, насчитывавшую 1,5—2 тыс. пар. Птицы обильно удобряли почву, гигроскопичный материал их гнезд накапливал влагу. В результате началось постепенное заболачивание, которое привело к постепенному исчезновению вереска и замене его жесткими злаками. Угодья стали использоваться как сенокосные. Однако в дальнейшем злаки были вытеснены ситником, позднее — конским щавелем и другими сорняками. Параллельно шло уменьшение численности грауса; на заболоченных участках появились утки — вид, менее ценный в качестве охотничьего трофея. Все это привело к отмене охраны чаек; более того, их гнезда стали намеренно разорять. К 1917 г. осталось около 30 пар, постепенно восстанавливался верещатник, исчезли утки, вновь появились граусы. Однако и к этому времени восстановление исходного сообщества было неполным: на вересковой пустоши сохранились злаки и некоторые сорняки — «след» сукцессионной серии» (И.А. Шилов, 1998).
Основная причина сукцессий — отсутствие равновесия между продукцией и дыханием в экосистеме, то есть неравновесность экологического баланса. Это приводит к изменению запаса органики в экосистеме и, в конечном счете, к изменению сообщества. Расположив разнообразные экосистемы на плоскости «продукция-дыхание» (рис. 3.8.1), мы можем убедиться, что устойчивыми из них оказываются только те, в которых продукция и дыхание уравновешивают друг друга.
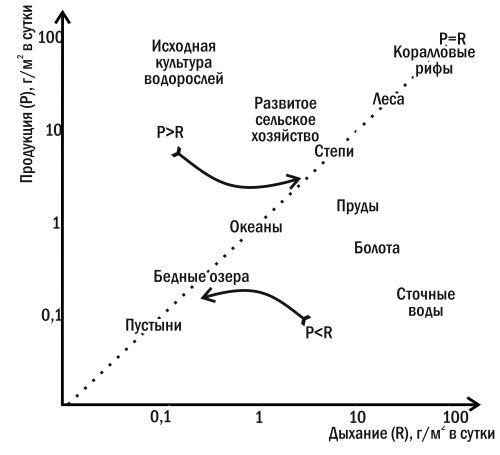
Рис. 3.8.1. Ординация (расположение) различных типов экосистем на плоскости «продукция — дыхание»
Если в каком-то сообществе продукция превышает дыхание (например, в исходной культуре водорослей — растворе минеральных солей, в который заселены фитопланктонные организмы), в нем начинается автотрофная сукцессия. Избыток органики накапливается в такой экосистеме, меняя ее свойства и создавая среду для размножения гетеротрофов. Продукция в сообществе снижается, дыхание увеличивается, и, в конце концов, эти две величины достигают равенства.
Гетеротрофная сукцессия (рис. 3.8.2) начинается с состояния, в котором дыхание сообщества превышает продукцию. Постепенно гетеротрофы разрушают избыток органики, и в системе достигается равновесие между дыханием и продукцией.
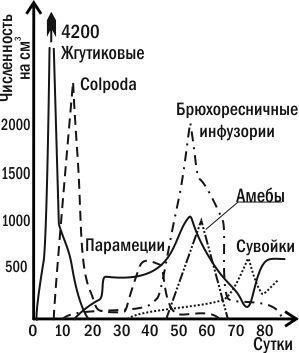
Рис. 3.8.2. Пример гетеротрофной сукцессии: динамика численности различных простейших в сенном настое
Характерным случаем автотрофной сукцессии можно считать заселение организмами территории, на которой нет запасов органического вещества. Рассмотрим следующий пример. На голой скальной породе поселяются лишайники, постепенно разъедающие камень с помощью выделяемых ими кислот. В трещинах горных пород появляется почва. На скале начинают расти мхи, а потом и травы. Формируя дерновину из переплетенных отмерших стеблей и корневищ, травы удерживают детрит и даже собирают пыль. Через очень длительное время на месте прежних скал появляются кустарники, а потом и лес. Это была первичная сукцессия — сукцессия, которая шла там, где не было запасов органического вещества от прежних сообществ (рис. 3.8.3).
Рис. 3.8.3. Пример первичной сукцессии: заиливание озера
Через какое-то время лес, выросший в ходе первичной сукцессии, уничтожается пожаром. Образуется гарь. На гари сохраняются остатки лесной почвы (обильно присыпанной пеплом) и даже семена многих видов растений. Уже на следующий год гарь зарастет травами (например, в таких условиях очень характерен иван-чай). Через несколько лет на месте гари появятся кустарники и молодые деревья, а уже через несколько десятилетий или через век здесь разовьется лес, напоминающий тот, который рос здесь до пожара. Вторичная сукцессия проходит в местообитаниях, где сохранились остатки органического вещества от предыдущих этапов развития данной экосистемы (рис. 3.8.4).
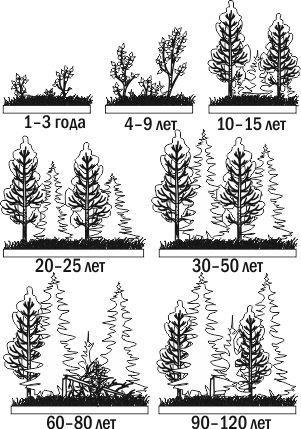
Рис. 3.8.4. Пример вторичной сукцессии: зарастание пожарища смешанным лесом
Гетеротрофная сукцессия требует для своего протекания запаса органики. Большинство примеров гетеротрофной сукцессии, которые мы можем наблюдать, разворачиваются не в масштабах биогеоценозов, а в пределах относительно небольших экосистем. Например, к их числу принадлежат тела умерших животных или стволы упавших растений. Протекающие в таких временных экосистемах сукцессии называются деструктивными — они не ведут к климаксу, а попросту уничтожают то местообитание, в котором происходят. Иногда можно наблюдать экоклины и в гетеротрофных сукцессиях. Раскопайте опавшую хвою и верхний слой почвы в хвойном лесу. Сверху хвоинки почти не повреждены, а чем глубже — тем более они оказываются преобразованными бактериями, грибами и животными-детритофагами. Каждая опавшая иголка проходит этот путь разрушения, соответствующий последовательности слоев хвойного опада.
Клементс был сторонником концепции моноклимакса. Сейчас восторжествовала концепция поликлимакса, согласно которой для каждого региона существует определенный климатический или зональный климакс, и, кроме того, в нем могут быть устойчивы определенные эдафические (т.е. обусловленные особенностями почв) или локальные климаксы. Описан также циклический климакс, при котором экосистема проходит через совокупность определенных сменяющих друг друга по кругу состояний. Иногда эти изменения определяются катострофическими событиями, закономерно происходящими на определенном этапе развития экосистем. Так, ельник по достижении определенной зрелости может закономерно уничтожаться вредителями; чапарраль может регулярно выгорать при пожарах.
Можно рассматривать как общие, так и частные сукцессии. Так, в плодовом саду на молодых деревьях питаются многоядные насекомые-фитофаги: хрущи, чернотелки, пяденицы, шелкопряды, листовертки. Когда деревья начинают плодоносить, появляются питающиеся цветками и плодами плодожорки, цветоеды, долгоносики. По мере старения сада его населяют стволовые вредители (короеды, древоточцы, стекляницы).
Коментарі
Проясните, пожалуйста
"Гетеротрофная сукцессия начинается с состояния, в котором дыхание сообщества превышает продукцию. Постепенно гетеротрофы разрушают избыток органики, и в системе достигается равновесие между дыханием и продукцией."
Вы пишете, что дыхание преобладает, и одновременно - что существует избыток органики, как и в случае с автотрофной сукцессией. Разве не должен быть недостаток этой органики, который ограничит размножение гетеротрофных организмов?
Или же я неправильно поняла...
Что означает, что сукцессия - гетеротрофная?
Это означает, что активность гетеротрофов (суммарно с дыханием автотрофов) превышает продукцию автотрофов. Это возможно только в том случае, если в местообитании есть избыток органики. Когда этот избыток органики будет уничтожен геторотрофами, дыхание и продукця в системе придут к равновесию...