Загадка сексу. Cui prodest: ген, індивід, група? Колонка для Комп'ютерри #131
Увага: питання! Якого рівня відбір (генетичного, індивідуального чи групового) є причиною переходу до статевого розмноження? І, якщо хочете, підказка. У тих випадках, коли якісь еволюційні нововведення сприяють збереженню та відтворенню як генів, так і індивідів і груп...
←
Dmytro Shabanov
→
Походження статі, роздвоєності та геміклонального успадкування. Постановка задачі
Загадка сексу. Cui prodest: ген, індивід, група?
Королева проблем еволюційної біології: кошмар Дженкіна повертається
Колонка для Комп'ютерри #130
Колонка для Комп'ютерри #131
Колонка для Комп'ютерри #132
Ну ось: я сам собі розписав програму майбутніх колонок, а тепер доведеться її виконувати! Час передсвятковий, у пристойних людей голови зайняті у кого підготовкою до застілля, у кого — оздобленням ялинки, у кого — складанням новорічних побажань. А мені доведеться виконати власну обіцянку і обговорити проблему статевого розмноження. Що ж, будемо вважати, що тема сексу сама по собі є досить святковою. На жаль, закрити її однією колонкою я не зможу. Давайте я зроблю так: запропоную вам новорічну колонку з безліччю картинок, що ілюструють походження статевого розмноження. Ця колонка завершиться питанням, свою версію відповіді на яке я дам лише наступного року. Буду радий почути (побачити в коментарях) вашу.
Почну здалеку. Ви, напевно, знаєте, що у земних організмів є два принципово різних типи клітин — прокаріотичний (доядерний) і еукаріотичний (ядерний). Вважається, що з трьох надцарств (найвищих систематичних категорій) два, бактерії і археобактерії, належать до прокаріот. Їхнє генетичне різноманіття, хоча б приблизно, можна оцінити за допомогою наступної схеми.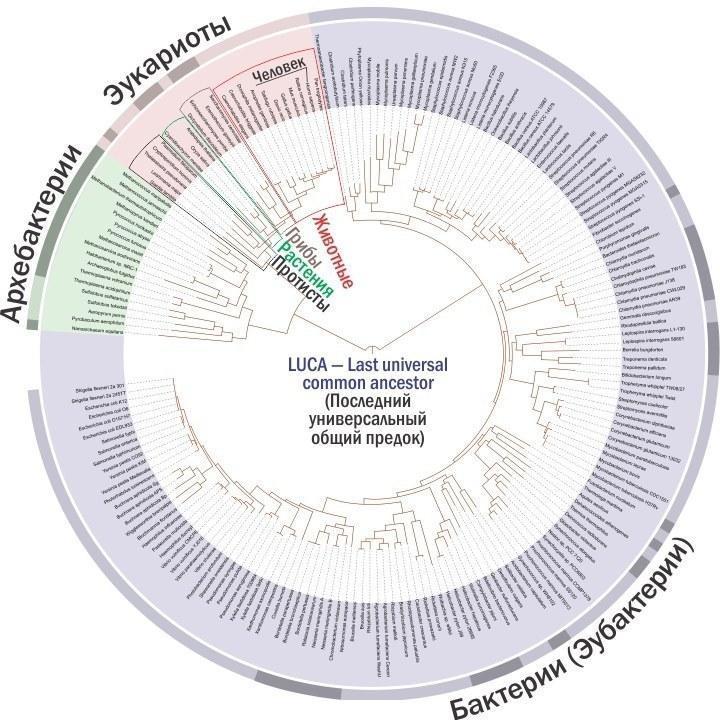
Це найзагальніша дендрограма (схема, що показує характер розгалуження «дерева життя»), що описує відносини між головними групами земних організмів. Тут можна розглянути її основу докладніше.
Вам здається, що тварини займають на цій схемі непропорційно мало місця? Так, непропорційність очевидна, лише вона має протилежний характер. Схема відображає генетичне різноманіття, і на ній саме непропорційно багато уваги приділено царствам Рослини, Гриби і Тварини — маленьким гілкам на гілці царства Протисти (куди входять і «прості» типу амеб і інфузорій, і різноманітні водорості, і багато одноклітинних грибоподібних організмів). А всі разом ці чотири царства еукаріот представляють лише невелику частину генетичного різноманіття прокаріот. З іншого боку, якби ми відображали кількість видів, представники багатомільйонного царства Тварини затиснули б інші групи.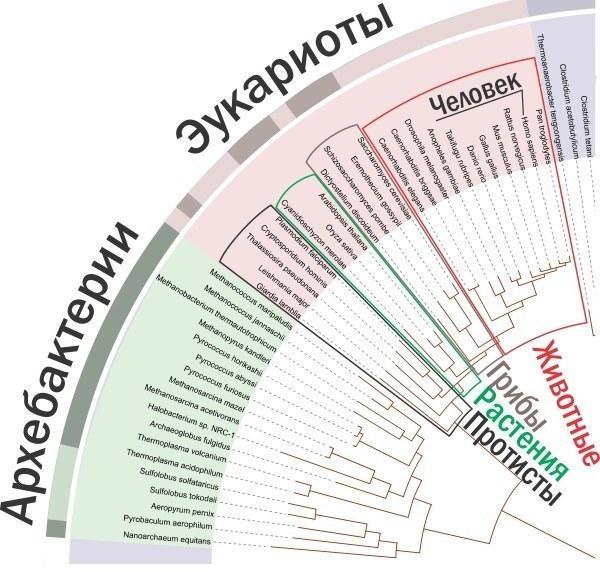
Збільшений фрагмент тієї ж схеми.
Подаючи схеми, треба розібратися, як вони будуються. Філогенетичні (відображаючі еволюційну історію) відносини реконструюються за генетичними даними за допомогою алгоритмів, що будують розгалужені дерева так, щоб мінімізувати передбачувану кількість еволюційних змін (про економну реконструкцію філогені я писав у колонці про звіра‑пегасофера). Коли мова йде про реконструкцію відносин всередині царства Тварини, цей підхід виявляється відносно адекватним. А ось походження рослин або виникнення самого надцарства Еукаріоти реконструюються з неминучими спотвореннями. Справа в тому, що еукаріотична клітина утворилася в результаті симбіозу кількох прокаріотичних клітин. Деталями зараз не займатимусь і обмежусь досить простою (і в силу цього відносно умовною) схемою. Крім того, на цій схемі не відображено, що в деяких групах ми реєструємо наслідки повторних симбіозів.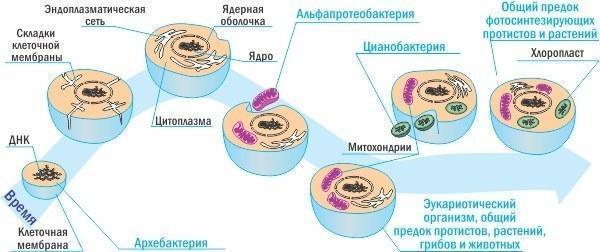
Симбіогенетичне походження еукаріотичної клітини (спрощена, дуже спрощена схема!)
Отже, цитоплазма (основний вміст) і ядро клітин тварин утворилися з предкової археобактерії, а мітохондрії — з предкової альфа‑протеобактерії (представника тієї ж групи еубактерій, до якої належить, наприклад, згадана в попередній колонці вольбахія — внутрішньоклітинний паразит/симбіонт членистоногих, здатний викликати паразитарно індукований партеногенез). Частина генів, що належала предку мітохондрії, перемістилася з мітохондрії в ядро. Вас дивує, як таке могло статися?
Дуже просто. Мітохондрія (як і будь‑який інший внутрішньоклітинний симбіонт) може загинути і розпастися. Її ДНК вивільниться в цитоплазму клітини‑хазяїна і, після ряду подорожей, може вбудуватися в хазяйську ДНК. Є ймовірність, що ядро почне виробляти ті білки, які раніше мітохондрія збирала для себе сама. Отримуючи ці білки від клітини‑хазяїна, мітохондрія підвищить ефективність своєї роботи, і така зміна може бути підтримана відбором. Однак тепер відбір перестане підтримувати працездатність вихідного гена в самій мітохондрії. З часом у мітохондрії цей ген дегенерує і зникне, а в ядрі залишиться його працездатна копія.
Як наші алгоритми реконструюють передісторію такої химерної клітини? Досліджуючи виключно ядерні гени, вони зблизять таку гібридну клітину з родичами її цитоплазми і ядра (у розглянутому нами прикладі — з археобактеріями, як показано на схемі). Досліджуючи мітохондріальні за походженням гени, ці алгоритми знайдуть її найближчих родичів серед альфа‑протеобактерій. Сітчастий характер філогенезу не може бути відображений у структурі розгалуженого дерева, а типові алгоритми реконструкції філогенезу вміють будувати лише такі дерева!
Реконструкція філогенії прокаріот ще складніша, бо їхні гени «гуляють» від групи до групи в результаті горизонтального перенесення (передачі між неродичними організмами; «вертикальною» вважається передача від предків до нащадків). Наші інструменти для реконструкції філогенії виявляються неадекватними для реконструкції таких відносин. Потрібно розробляти нові – і тоді вже будувати щось на кшталт схем, показаних нижче.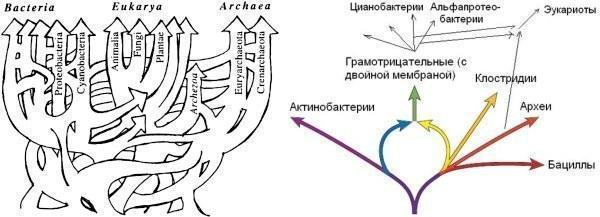
Зліва — наполовину умовна «сітчаста» схема, що показує відносини між трьома надцарствами (джерело). У основі схеми — мережа, яка стала результатом численних горизонтальних переносів. Справа — «кільцева» схема еволюції основних груп бактерій (джерело). Найрізноманітніша група, грамнегативні (до яких, окрім інших, належать предки мітохондрій і хлоропластів), розглядається тут як результат симбіозу двох інших груп, актинобактерій і клостридій.
А як же LUCA, «останній спільний предок», показаний на першій схемі? Я згоден з тими, хто вважає, що LUCA був не окремим видом, а складним співтовариством, члени якого обмінювалися генами як їм заманеться.
Знаєте, навіщо я розповідав вам про реконструкцію філогенії прокаріот? Щоб переконати вас: прокаріоти досягли успіху в рекомбінації, утворенні нових поєднань генетичної інформації. Виділяють різні типи рекомбінацій (якщо хочете розібратися докладніше — дуже рекомендую ці і ці статті). З них для нашої подальшої розмови найцікавіша гомологічна рекомбінація.
Уявіть собі клітину кишкової палички, найкраще вивченої бактерії, яка отримала фрагмент чужої ДНК. У цитоплазмі цих бактерій може знаходитися плазміда, невелика кільцева молекула ДНК, яка називається F‑фактором. Бактерія, що має F‑фактор, здатна вирощувати пілі (щось на кшталт трубопроводу) і закачувати по ньому, по‑перше, копію F‑фактора, і, по‑друге, скопійовані фрагменти бактеріальної хромосоми.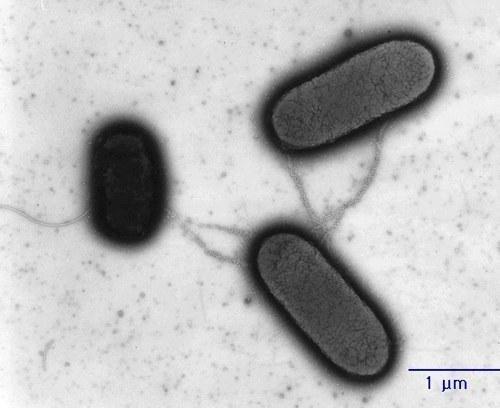
Знаєте, що роблять ці три кишкові палички, Escherichia coli? Вони обмінюються фрагментами своєї ДНК за допомогою пілій, що нагадують голку шприца (джерело).
Клітина‑донор може використовувати чужі фрагменти ДНК для гомологічної рекомбінації. При цьому вона встановлює відповідність між отриманим фрагментом ДНК і власною ДНК клітини. Спеціальна білкова система клітини‑донора замінює наявний генетичний текст на отриманий.
Важливою особливістю гомологічної рекомбінації є те, що новий генетичний текст вбудовується не кудись, а саме в те місце на бактеріальній хромосомі, де була його «рідна» версія. Нові гени потрапляють при цьому на ті ділянки, де розташовані системи, що керують їх активністю.
У гомологічній рекомбінації у бактерій можна спостерігати один із етапів становлення тієї форми гомологічної рекомбінації, яка характерна для еукаріот. Я говорю про статеве розмноження.
Строго кажучи, статевий процес зовсім не обов’язково пов’язаний з розмноженням. І у кишкової палички, і, наприклад, у інфузорій (еукаріотичних організмів) статевий процес протікає сам по собі, а розмноження — ніби сам по собі. Але все ж найхарактернішим для еукаріот виявляється гапло‑диплоїдний життєвий цикл з заплідненням і мейозом. Потрібно пояснити. Колись я це вже робив, але зараз повторю ще раз, інакше (можете для кращого розуміння порівняти два пояснення).
Почнемо з чого. Залежно від кількості комплектів генетичної інформації (хромосомних наборів), що містяться в ядрі, еукаріотичні клітини діляться на гаплоїдні (один комплект), диплоїдні (два хромосомних набори) і ще цілий ряд інших типів, які зараз не важливі. Є два головних типи поділу клітин еукаріот. При мітозі кількість генетичної інформації не змінюється, утворюються дві клітини, генетично ідентичні одна одній і материнській клітині. При мейозі (який, по суті, складається з двох поділів, схожих на мітоз, але не передбачає подвоєння генетичної інформації між ними) отримуються чотири клітини зі вдвічі зменшеною кількістю генетичної інформації. Усі ці клітини генетично унікальні, бо між гомологічними парами хромосом відбувається гомологічна рекомбінація.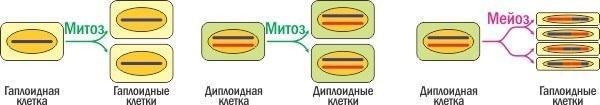
Порівняння двох основних типів поділу еукаріотичних клітин.
Якщо мейоз зменшує кількість хромосом удвічі, то в життєвому циклі, де він присутній, має відбуватись щось, що компенсує їх нормальну кількість. Це запліднення, злиття двох клітин, яке складається з двох етапів — сингамії (злиття цитоплазм) і кариогамії (злиття ядер).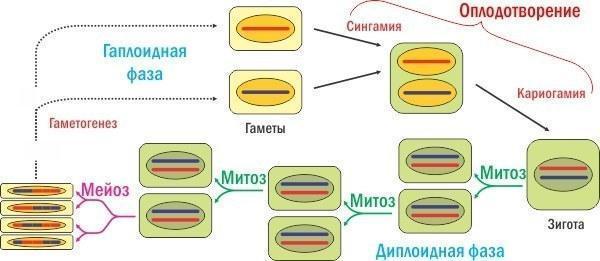
Приклад гапло‑диплоїдного життєвого циклу з заплідненням і мейозом. Чи могло таке складне винахід незалежно виникнути в ході еволюції життя багато разів, у різних групах?
На намальованій мною схемі мітози показані лише на диплоїдній фазі, як це зазвичай буває у високоорганізованих тварин, і у людей включно. Це не єдине рішення. Є види, у яких мітози і ріст організму відбуваються на гаплоїдній фазі, а диплоїдна виявляється дуже короткою (одразу після запліднення зигота ділиться за допомогою мейозу). У більшості рослин ріст відбувається на обох фазах. У мохів переважає гаплоїдна фаза, а, наприклад, у квіткових — диплоїдна.
Тепер зрозуміло? І, звичайно, найцікавіше, як у високоорганізованих еукаріот міг виникнути такий складний життєвий цикл. Його різні етапи виникали окремо, і досі окремо спостерігаються у деяких протистів і грибів. Свої (досить складні) механізми забезпечують у деяких еукаріот сингамію (наприклад, зрощення окремих грибних ниток). Відомо багато багатоядерних еукаріот; у деяких з них зафіксовано кариогамію. Як я вже казав, мейоз включає високоорганізований механізм гомологічної рекомбінації, простіші форми якої виникли ще у прокаріот. Є вагомі підстави вважати, що мейоз еволюціонував як форма відновлення, ремонту (репарації) генетичного апарату клітини — і лише потім став невід'ємним етапом гаплоїдно‑диплоїдного життєвого циклу.
Цілий ряд авторитетів передбачає, що гаплоїдно‑диплоїдний життєвий цикл із заплідненням і мейозом виникав у ході еволюції неодноразово! Це підтверджується тим, що інтимні молекулярні механізми, що забезпечують ці суттєві перестройки клітин, у різних групах виявляються різними. Додатковим аргументом, що підтримує цю гіпотезу, є поширення сингамії, кариогамії і мейозу у різних групах протистів: у багатьох їх типах ці феномени зафіксовані у просунутих представників і відсутні у примітивних. Якщо це так, ми переконуємося, що перехід до такого життєвого циклу — закономірна подія в еволюції багатьох груп еукаріот.
І ось тепер настав час задати питання, заради якого я писав цю колонку. Воно стосується рівня відбору, відповідального за появу статевого розмноження.
Мені доводилося писати, що проблема рівня, на якому відбувається відбір, є предметом запеклої дискусії багатьох біологів:
— Чарльз Дарвін передбачав, що еволюція — результат відбору індивідів, а іноді — і їх груп;
— Верно Вінн‑Едвардс вважав головним механізмом еволюції відбір груп;
— Джордж Вільямс розбив багато аргументів Вінна‑Едвардса, обґрунтовуючи, що еволюцію рухає практично виключно відбір індивідів;
— Річард Докінз, розвивавший ідеї Вільяма Гамільтона та багатьох інших теоретиків (включаючи Едварда Вілсона), заявив, що еволюцію рухає відбір окремих генів;
— Едвард Вілсон змінив свою точку зору і підкреслив важливість групового відбору для деяких ключових еволюційних переходів;
— багато біологів старшого покоління кардинально заперечують ідею «егоїстичного гена»;
— багато молодих біологів (особливо молекулярних, не звиклих задумуватись про загальні питання) переконані, що «сучасна» біологія «довела»: відбір відбувається лише на рівні генів.
Ось біологію і розривають дискусії про «егоїстичний ген», «егоїстичний індивід» і «егоїстичну групу».
Тож, увага: питання!
Відбір якого рівня (генного, індивідуального чи групового) є причиною переходу до статевого розмноження?
І, якщо хочете, підказка. У випадках, коли якісь еволюційні нововведення сприяють збереженню і відтворенню як генів, так і індивідів і груп, ми не зможемо встановити, відбір якого рівня відповідає за їх розвиток. Потрібно шукати такі зміни, які, припустимо, вигідні для відтворення генів, що їх викликають, але порушують розмноження індивідів і груп (або, наприклад, сприяють збереженню, розселення та розмноженню груп, але при цьому порушують відтворення генів і індивідів).
З Новим, 2014 роком вас, дорогі читачі!
←
Dmytro Shabanov
→
Походження статі, роздвоєності та геміклонального успадкування. Постановка задачі
Загадка сексу. Cui prodest: ген, індивід, група?
Королева проблем еволюційної біології: кошмар Дженкіна повертається
Колонка для Комп'ютерри #130
Колонка для Комп'ютерри #131
Колонка для Комп'ютерри #132