Короткий виклад епігенетичної теорії еволюції, або ЕТЕ для зайнятих. Колонка для Комп'ютерри #128
Чим складніший організм, чим більше взаємозв’язків проявляється в його онтогенезі, тим суттєвіша передісторія виду, досягнутий ним адаптивний компроміс буде спрямовувати його можливі зміни. ЕТЕ дозволяє нам зрозуміти, що життя «йде на ощуп» (© П. Тейяр де Шарде...
←
Dmytro Shabanov
→
Як у ході еволюції виникають адаптивні ознаки, або Яку з теорій еволюції підтверджують дані сучасної генетики?
Коротке викладення епігенетичної теорії еволюції, або ЕТЕ для зайнятих
Україна — велика Врадіївка. Вибрані місця з листування з російськими та проросійськими друзями
Колонка для Комп’ютерри #127
Колонка для Комп’ютерри #128
Колонка для Комп’ютерри #129
Два тижні пройшли у мене під знаком співпереживання потрясінням в українській політиці та спорів про епігенетичну теорію еволюції. Про політику — не зараз; тут ми обговоримо ЕТЕ. Я регулярно чую докори в відсутності її короткого викладення. Ця колонка — моя спроба такого викладення, що враховує свіжий досвід обговорення ЕТЕ на сайті КТ, у ЖЖ Олександра Маркова, на моєму сайті, на засіданні клубу «Еволюція» в Києві (ось розширена презентація мого доповіді), на круглому столі з Сергієм Яструбовим, проведеному під час молодіжної конференції харківського біофаку.
Так, не забудьте:
— тут не оспорюється факт еволюції, а йдеться про проблеми вивчення її механізмів;
— викладені тут погляди не мають нічого спільного з ідеями Т. Д. Лисенка, «розумним замислом», «науковим креаціонізмом» та іншими конструкціями, що судять про еволюцію на основі ідеологічних чи релігійних догм;
— тут відображено моє розуміння ЕТЕ; її творці та інші прихильники можуть мати (і часто мають) інше бачення багатьох важливих для цієї теорії питань.
І ще. Уточнимо терміни.
Наука (приватна) — розвиваючийся комплекс уявлень про певний аспект дійсності, технологій його вивчення та зміни, який може включати в себе різні, зокрема частково суперечливі один одному, гіпотези та теорії.
Теорія — цілісна система поглядів, у якій одні положення виводяться з інших. Теорією може стати в процесі свого розвитку гіпотеза, що пояснює певні феномени і має прогностичну цінність.
Еволюція — незворотні зміни біосистем протягом історичного часу біосфери. Призводить до зміни існуючих біосистем, зокрема їх ускладнення, підвищення їх адаптованості до середовища, зростання їх стійкості, виникнення у них нових властивостей і появу нових типів біосистем. Еволюція — багаторівневий процес; еволюціонують популяції, види, надвидові групи, а також спільноти та екосистеми.
Еволюція еволюції — зміни механізмів еволюції у міру еволюції біосистем.
Еволюційна біологія — наука, що вивчає механізми еволюції. Вивчення того, як саме відбувалась еволюція, теж часто включають у сферу компетенції цієї науки, але тут вона трактується в вузькому, зазначеному сенсі.
Онтогенез — індивідуальний розвиток особини, сукупність її закономірних і випадкових перетворень протягом її життя.
Відбір — переважне розмноження особин і їх груп, залежне від їх властивостей; відбір переважно зберігає і розмножує більш адаптивних особин.
Адаптивність — відповідність організму тому середовищу, у якому він розвивається, що дозволяє успішно пройти онтогенез і залишити нащадків.
Отже, як ви зрозуміли, еволюційна біологія — наука, яка включає безліч теорій. Її розвиток не завершений, і повної картини механізмів багаторівневого процесу еволюції на сьогодні немає. Розглядаючи історію еволюційної біології, ми можемо побачити, що в ній то поширювалися ідеї, прийняті більшістю вчених, то настали часи розбіжностей у поглядах. Щоб їх описати, я скористаюсь схемою, запропонованою Н. Н. Воронцовим (заповнюючи останній рядок таблиці, заздалегідь відобразивши те, до чого приходжу в цій колонці).
Однією з теорій, що з’явилися в епоху кризи II синтезу, стала ЕТЕ. Вона базується на результатах, отриманих у 1940‑х і 1950‑х роках радянським зоологом І. І. Шмальгаузеном (теорія стабілізуючого відбору) і англійським генетиком К. Х. Воддингтоном (епігенетичний ландшафт і «генетична асиміляція модифікацій»). Основи ЕТЕ сформулював московський палеонтолог М. А. Шишкін у працях, опублікованих з 1984 по 1988 рік. У розробку теорії внесли також його колеги А. П. Расницин (метафора адаптивного компромісу) і А. С. Раутиан (еволюція як підтримання стійкості).
Описуючи ЕТЕ, її порівнюють зі СТЕ, синтетичною теорією еволюції, маючи на увазі саме цілісну теорію, що склалася до середини ХХ століття. Чому? Сучасна еволюційна біологія — досить розпливчастий і в певній мірі внутрішньо суперечливий комплекс концепцій. Кожна з них більш‑менш пояснює певний комплекс факторів, ігноруючи інші дані. Однак СТЕ завдяки своїй простоті досі залишається версією «за замовчуванням»: її викладають у школах і вузах, намагаючись ототожнити з сучасною еволюційною біологією цілком.
Настав час дати короткий опис ЕТЕ.
Епігенетична теорія еволюції розглядає еволюцію як процес зміни одних стабілізованих шляхів онтогенезу іншими. У представників високоорганізованих груп результат онтогенезу обумовлений надзвичайно складним комплексом факторів і результатами взаємодії їх ефектів.
На онтогенез впливає взаємодія наступних факторів і їх ефектів:
наступних спадкових задатків:
генетичних (послідовностей нуклеотидів у нуклеїнових кислотах — НК);
епігенетичних (хімічних і просторових модифікацій макромолекул НК);
інших (пов’язаних з організацією цитоскелету, набором РНК і регуляторних молекул, конформацією білків тощо);
різноманітних середовищних впливів;
випадковості.
Результат онтогенезу неможливо передбачити однозначно. Його можна охарактеризувати лише розподілом ймовірностей різних результатів, серед яких слід виділити норму (стан, який підтримується стабілізуючим відбором) і різноманітні морфози (відхилення, аберації). Метафора, що описує розподіл можливих підсумків онтогенезу, — епігенетичний ландшафт К. Х. Воддингтона. З цієї точки зору можливі онтогенетичні шляхи можна описати як сукупність стабілізованих ділянок (креодів), точок біфуркації та розділяючих креодів сукупностей малоймовірних і нестійких станів. Стабілізуючий відбір — переважне збереження і розмноження особин, онтогенез яких привів до норми, — підвищує стійкість розвитку норми (збільшує її ймовірність). Ця стійкість зростає і завдяки зростанню еквіфінальності розвитку норми (здатності реалізовувати норму у все більш різних особин), і завдяки підвищенню автономності такого розвитку (здатності реалізовувати норму у все більш різних умовах середовища). Це забезпечується тим, що відбір перебудовує систему управління онтогенезом в цілому (і генотип зокрема). У моделі епігенетичного ландшафту дія стабілізуючого відбору виглядає як поглиблення відповідного креода.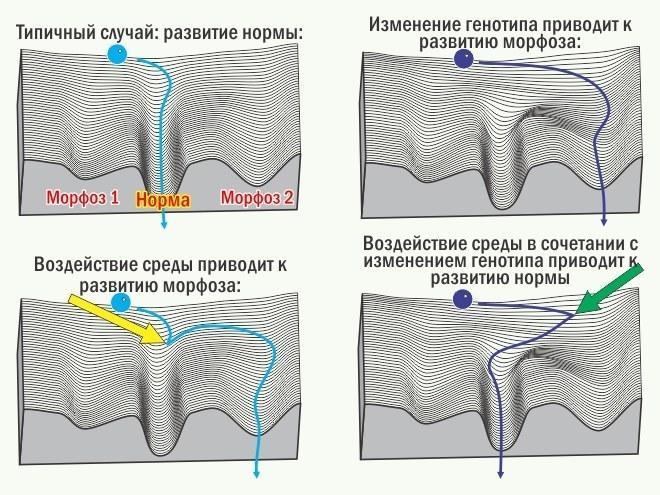
Якщо характер відбору змінюється і він перестає підтримувати попередню норму, її розвиток дестабілізується і проявляються різноманітні морфози. Якщо якийсь з морфозів виявляється адаптивним, відбір вибірково зберігає ті системи управління онтогенезом, які привели до такого адаптивного стану. Нащадки подібних особин з більшою ймовірністю будуть адаптивними, якщо їх онтогенез приведе до того ж результату. Отже, відбір буде підтримувати тих нащадків, у яких розвиток адаптивного в даних умов стану буде ставати все більш стійким (все більш ймовірним). Результатом стає підвищення стійкості розвитку підтримуваного відбором морфоза, тобто зростання його успадкованості. Таким чином, сам феномен успадкованості виявляється результатом відбору.
Типовим випадком зміни норм у ході еволюції слід вважати той, при якому адаптивний морфоз виникає як адекватна відповідь системи управління онтогенезом на змінені умови розвитку. Якщо відбір підтримує такий морфоз, він стає новою нормою, його розвиток автономізується і набуває незалежності від специфічних зовнішніх впливів.
У міру еволюції відбувається ускладнення системи управління онтогенезом і вдосконалення механізмів, що забезпечують пошук адаптивних морфозів при зміні характеру відбору. Поява генетичної успадкованості, статевого розмноження, культурної успадкованості, складної соціальної організації — деякі етапи цього процесу.
Чим описаний підхід відрізняється від підходу СТЕ? Для СТЕ еволюція — це перебудова генотипу внаслідок відбору за результатами активності генів, відображених у ознаках організму. Саме тому СТЕ намагається описати онтогенез як сукупність відносно незалежних причинно‑наслідкових зв’язків. Будь‑які взаємодії, що ускладнюють передачу інформації від генотипу до фенотипу, для СТЕ просто перешкоди, що ускладнюють перебудову генотипу за його фенотипічними ефектами.
Якщо інформація від генотипу відображається у фенотипі безпосередньо, механізм СТЕ працює цілком задовільно. Рівняння Харді‑Вайнберга описує, як співвідношення алелів (альтернативних варіантів одного гена) у нащадків залежить від такого у предків. Математичний апарат СТЕ базується на тому, що кожен алель вносить постійний внесок у підсумкову пристосованість свого носія. Якщо алель підвищує пристосованість організму, відбір підвищуватиме його частоту, і з часом сприятливий алель витіснить свої альтернативи. Є випадки, коли така модель працює. Розглянемо два штами бактерій. Один росте швидше, але нестійкий до антибіотика. Інший — стійкий і «платить» за це повільнішим ростом. Їх ознаки однозначно відображають їх генотип. Їх динаміка на середовищах з різним вмістом антибіотиків добре описується рівняннями відбору за СТЕ.
Якщо модель СТЕ вірна, еволюція еволюції повинна призводити до того, що фенотип буде все ефективніше, все «прозоріше» відображати генотип. У таких організмів відбір перестраюватиме генотип особливо ефективно. Вони почнуть швидше виробляти пристосування і отримають перевагу в мінливому середовищі. А ось організми зі складними взаємодіями різних факторів в онтогенезі мають еволюціонувати повільно. Особливо «важко» — види, що складаються з довгоживучих і малоплідних особин. Чи виправдане це передбачення?
Ні! Я писав про це, пам’ятаєте?
У тому світі, який ми спостерігаємо, найскладніші, найшвидше еволюціонуючі з точки зору змін своєї будови та поведінки організми виявляються абсолютно неймовірними з точки зору СТЕ. Візьмемо, наприклад, людей. Наш генотип містить зовсім небагато інформації, вміщуючись у незаархівованому вигляді на CD‑диску. Самі гени становлять його невелику частину; близько 25 тисяч наших генів вимагає для запису менше 10 мегабайт (а архіватор їх ще сильніше стискає). Нагадаю відому жарт, що файл з детальним описом форми колінного чашечка (однієї з найпростіших наших кісток) у AutoCad займе більше місця. Інформація, необхідна для опису нашої будови, несумісно більша за об’єм нашого геному. Ще дивовижніше — масив інформації у нашій психіці. Це означає, що в нашому онтогенезі відбувається головокружна кількість виборів можливих шляхів розвитку з запам’ятовуванням їх результату.
Типовим шляхом еволюційних змін для ЕТЕ виявляється той, що відповідає логіці експериментів Г. К. Воддингтона. Воддингтон викликав у експериментальних тварин морфози за допомогою зовнішніх впливів. Потомство від схрещування носіїв морфозів знову піддавали аналогічним впливам, і знову для розмноження відбирали носіїв того ж морфозу. Через невелику (перші десятки) кількість поколінь ці морфози почали розвиватися без специфічних впливів. Нестабільний, залежний від середовища шлях розвитку ставав стабільним. Щоб переконатися, що мова не йде про успадкування набутих ознак, порівняйте ці експерименти з класичними експериментами Вейсмана.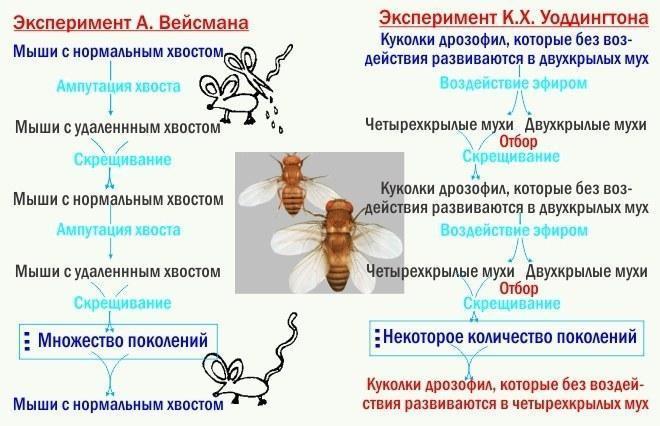
На мові СТЕ досліди Воддингтона описуються натягом. Відбір на здатність розвивати певну модифікацію (неусадкована ознака) привів до зміни множини генів‑модифікаторів і в кінцевому підсумку до «генетичної асиміляції модифікації», передачі управління розвитком цією ознакою в генотип. Це пояснення передбачає існування певних генів‑модифікаторів, не виявлених геномікою. Воно передбачає швидку еволюцію цих модифікаторів, що не відповідає моделям відбору, розробленим у самій СТЕ. Якщо прийняти, що мова йде не про віртуальні модифікатори, а про інші структурні гени, стає незрозуміло, чому «генетична асиміляція» контролю розвитку однієї ознаки не призводить до хаосу в розвитку інших.
На мові ЕТЕ досліди Воддингтона описуються просто. Підтримуючи морфоз, стабілізуючий відбір підвищує стійкість його розвитку. І — зверніть увагу! — жодних натягнутих спроб представити організм як суму ознак, а генотип — як суму генів. Досліди Воддингтона описують не перетворення «неусадкованих» ознак у «усадковані», а вплив відбору на стійкість розвитку. І, до речі, приклад людей показує, що часто «неусадковані» ознаки (те, чому ми навчаємося) не менш важливі для нас, ніж якісь інші.
Тепер можна обговорити різноманітність оцінок ЕТЕ. Вони дуже різні. Один полюс полягає в тому, що ЕТЕ — повна альтернатива СТЕ, і жодного компромісу між ними неможливо. На іншому полюсі знаходяться ті, хто оголошує ЕТЕ лженаукою і навіть намагається заборонити згадування імен її прихильників у присутності студентів, щоб не посіяти сумніви в безальтернативній істинності СТЕ. Повірте, я пишу про такі спроби не абстрактно, а на основі сумного досвіду... Моя оцінка ближче до першого полюса, хоча відрізняється від нього. Я вважаю, що ЕТЕ може стати ядром III синтезу: потенціал для пояснення того, що відбувається на організмовому рівні, є лише у цієї теорії.
Звичайно, у ЕТЕ зберігається безліч недостатньо розроблених питань. Одне з них — опис різноманітності ознак з точки зору регуляції їх розвитку в онтогенезі. Ймовірно, навіть у складних організмів є відносно прості ознаки, майже однозначно залежні від стану окремих алелів. У цих випадках моделі СТЕ будуть описувати еволюцію таких ознак досить адекватно. Порушення нормальних шляхів розвитку, ймовірно, регулюються простіше, ніж активація вже існуючих генних механізмів у тих тканинах, де вони зазвичай працюють. Але виникнення принципово нових ознак такими механізмами не пояснити...
Отже, на мій погляд, ЕТЕ — більш широке узагальнення, ніж СТЕ, і випадки, коли підхід СТЕ виявляється застосовним, можна розглядати (за вдалим виразом С. Яструбова) як вироджені (спрощені) випадки застосовності ЕТЕ.
Важливою перевагою ЕТЕ, на мій погляд, є її здатність пояснювати швидке (за еволюційними мірками) появу адаптивних нововведень, гармонійно вписаних у комплекс ознак організму. Для СТЕ нові адаптивні ознаки — результат щасливої випадковості, мутації структурного або регуляторного гена, яка виявилася корисною. Чим складніший організм, тим рідше мають відбуватись такі випадковості. Для ЕТЕ нові ознаки виникають як відповідь цілісного організму на змінені умови його розвитку. У формуванні цієї відповіді відображається весь досвід попередньої еволюції, результати відбору в еволюційному минулому. Шансів, що така відповідь буде адекватною новим умовам, набагато більше.
Чим складніший організм, чим більше взаємозв’язків проявляється в його онтогенезі, тим суттєвіше попередня історія виду, досягнутий ним адаптивний компроміс будуть спрямовувати його можливі зміни. ЕТЕ дозволяє нам зрозуміти, що життя «йде на дотик» (© П. Тейяр де Шарден), а не дрейфує за волею випадковості.
Я дякую за критику цього тексту Олександра Павловича Расницина. Я зміг частково врахувати його зауваження, але хочу підкреслити, що він не несе відповідальності за недоліки моїх пояснень. Чи вдалося мені згадати все, що важливо для розуміння ЕТЕ? Звичайно, ні. Декілька моментів, про які я мовчав, можна зрозуміти з презентації. Вступ (обговорення термінів, дисклеймер) у цій колонці побудовано приблизно так само, як у презентації, а ось у подачі самої характеристики ЕТЕ є відмінності. Якщо хочете по‑справжньому розібратися, спробуйте вникнути і в іншу версію того ж пояснення.
Думати про ці речі мені цікаво. А вам?
←
Dmytro Shabanov
→
{
"title": "Як у ході еволюції виникають адаптивні ознаки, або Яку із теорій еволюції підтверджують дані сучасної генетики?",
"summary": "Короткий виклад епігенетичної теорії еволюції, або ЕТЕ для зайнятих Україна — велика Врадієвка. Вибрані місця з переписки з російськими та проросійськими друзями",
"body": "Колонка для Компьютерри #127
Колонка для Компьютерри #128
Колонка для Компьютерри #129"
}
Колонка для Комп'ютерри #127
Колонка для Комп'ютерри #128
Колонка для Комп'ютерри #129