Чи достатньо вісім косвенних гіпотез, що описують переваги статевого розмноження, щоб пояснити виникнення статі? Колонка для Комп'ютерри #133
Якщо виграш у виживаності потомства самців і самок не досягає двократного, клональні організми в короткостроковій перспективі витісняють роздвоєно‑статеві. Довгострокові переваги статевого розмноження в такій ситуації просто не встигнуть проявитися! Але чому ж статеве розмноження розповсюджується…
←
Dmytro Shabanov
→
Королева проблем еволюційної біології: кошмар Дженкіна повертається
Чи вистачить восьми непрямих гіпотез, що описують переваги статевого розмноження, щоб пояснити виникнення статі?
Чи правда, що королева еволюційних проблем — це Червона (a.k.a. Чорна) Королева? Не поспішайте з відповіддю — не забувайте про гермафродитів!
Колонка для Комп’ютерри #132
Колонка для Комп’ютерри #133
Колонка для Комп’ютерри #134
Парадоксальність статевого розмноження, про яку йшлося в моїй попередній колонці, така, що деякі читачі не змогли її усвідомити. Подумайте: при статевому розмноженні будь‑який ген індивіда передається нащадку лише з 50‑відсотковою ймовірністю, а при різних формах клонального відтворення — з ймовірністю 100 %!
Розглянемо ще раз сценарій, який обговорює більшість авторів. Вид з клональним відтворенням переходить до статевого розмноження (з самцями і самками). Як правильно зазначив John Maynard Smith, такий перехід може бути обумовлений лише вдвічі вищою виживаністю нащадків від статевого розмноження, причому одразу, у короткій перспективі. Це найпростіше продемонструвати за допомогою таблиці. Уявіть собі популяцію, що ділиться на дві субпопуляції (частини), A і B, які різняться за характером розмноження (класифікація його типів подана в цій колонці).
«Змагання» 1
Тип
відтворення
Виробники
Нащадки
Виживаність
Виживе
Всього
Результат
Субпопуляція A
Клональність
(тип I)
Індивід 1
100
25 %
25
50
Програш
Індивід 2
100
25
Субпопуляція B
Роздільна статевість
(тип III)
Самка
100
51 %
51
51
Перемога
Самець
—
У показаному змаганні субпопуляція B вигоне всіх представників субпопуляції A за відносно невелику кількість поколінь. Але зверніть увагу: перемога статевого розмноження в цьому випадку пов’язана з неймовірним, що перевищує двократний, зростанням виживаності нащадків (ймовірності для них дожити до певного порівнянного етапу їхнього життєвого шляху). Такий стрімкий зріст виживаності має бути зумовлений якимось надзвичайно вагомим перевагою! Хіба це — просто перестановка генів?
А якщо приріст виживаності буде хоча б трохи меншим? Подивимося.
«Змагання» 2
Тип
відтворення
Виробники
Нащадки
Виживаність
Виживе
Всього
Результат
Субпопуляція A
Клональність
(тип I)
Індивід 1
100
25 %
25
50
Перемога
Індивід 2
100
25
Субпопуляція C
Роздільна статевість
(тип III)
Самка
100
49 %
49
49
Програш
Самець
—
Якщо виграш у виживаності нащадків не досягає двократного, клональні організми в короткостроковій перспективі витісняють роздільностатевих. Довгострокові переваги статевого розмноження в такій ситуації просто не встигнуть проявитися!
Але чому ж статеве розмноження так широко поширене? Це вимагає особливого пояснення, і в цій колонці я опишу його різні варіанти. Про «вікаріум з Брея», згаданий минулого разу, ви, я сподіваюся, пам’ятаєте.
Гіпотеза «егоїстичного гена статі» Доукінза — версія, яку читачі трьох останніх колонок згадували найчастіше.
"Труднощі, з якими стикаються теоретики, намагаючись пояснити еволюцію статі, принаймні частково пов’язані з тим, що за їхніми уявленнями індивід намагається максимізувати число своїх виживаних генів. У світлі таких уявлень статеве розмноження сприймається як щось парадоксальне. <...> Однак цей парадокс здасться менш парадоксальним, якщо згідно з аргументами, наведеними у моїй книзі, розглядати індивіда як машину виживання, створену короткоживучою конфедерацією довгоживучих генів. <...> Якщо статеве розмноження як протилежне безстатевому сприятливе для гена, що визначає статеве розмноження, то цього достатньо для пояснення існування останнього. Чи сприятливе воно для всіх інших генів даного індивіда, не дуже важливо. З точки зору егоїстичного гена статеве розмноження зовсім не таке вже й дивне явище". Richard Dawkins. The Selfish Gene
Щоб правильно зрозуміти Доукінза, треба розібратися, що він вкладає у поняття «ген».
"...загальноприйнятого визначення гена не існує. <...> Я хочу скористатися визначенням, що належить J. Williams: ген — будь‑яка порція хромосомного матеріалу, що зберігається протягом достатньої кількості поколінь, щоб слугувати одиницею природного відбору. <...> Ген — це реплікатор з високою точністю копіювання". Richard Dawkins. The Selfish Gene
Я, звичайно, не прив’язував би так однозначно поняття «природний відбір» до рівня генів... Зберігаючи сенс, підхід Доукінза можна сформулювати так: ген — фрагмент генетичного тексту, який передається з покоління в покоління як єдине ціле (що не виключає можливості його змін). Подивимося, чи існує «ген статевого розмноження» як щось єдине.
Ні! У позапрошлій колонці я не випадково витратив чимало сил, пояснюючи, що статеве розмноження — феномен, пов’язаний з гапло‑диплоїдним життєвим циклом з заплідненням і мейозом. У цьому еволюційному винаході зійшлися механізми гомологічної рекомбінації, що досягли досконалості в мейозі, а також системи, що забезпечують сингамію і кариогамію (пояснення цих термінів — у тій колонці). Пояснювати виникнення статевого розмноження як виникнення якогось «гена», який раптом набув такого ефекту, — вірити в казку.
Чи ж пояснення Доукінза повністю помилкове? Теж ні. Згадайте (знову ж із позапрошлої колонки) про інфекційну передачу «статі» у бактерій. Кишкова паличка, у цитоплазмі якої знаходиться невелика кільцева молекула ДНК, названа F‑фактором, у зв’язку з цим набуває здатність формувати пілі (структури для ін’єкції ДНК) і передавати іншим бактеріям і F‑фактор, і інші фрагменти ДНК. «Статева» особина заражає «статтю» безстатеву, всклавши в ту піль. У цьому випадку пояснення Доукінза виявляється, принаймні, цілком працездатним. У організмів, що мають гапло‑диплоїдний життєвий цикл з заплідненням і мейозом, сама готовність особи до розмноження — результат узгодженої роботи багатьох незалежно виниклих і еволюціонуючих генів, кожен з яких стикається з 50‑відсотковою ймовірністю не потрапити у наступне покоління. «Егоїстичного гена статі» у таких організмів просто не існує!
А яка ж перевага забезпечила тріумф статевого розмноження? На цю тему висловлено чимало гіпотез, і кілька з них я хочу коротко описати. Детальніше ці гіпотези охарактеризовані у «Червоній королеві» Метта Рідлі, у книзі Вігена Геодакяна «Два статі. Навіщо і чому?», а також у статтях К. Ю. Попадіна та І. А. Рухленка.
Гвинтик Меллера. Герман Меллер звернув увагу на те, що при клональній передачі геномів виникаючі в них помилки — мутації — можуть бути виправлені лише смертю або відстороненням від розмноження їх носія. Якщо носії генотипів, вільних від помилок, зникнуть через мутаційний процес або просто випадковістю, най«чистішими» стануть генотипи, що несуть по одній мутації. Ця ситуація може зберігатися певний час, але все одно рано чи пізно завершиться випадковим зникненням генотипів з однією мутацією. Тоді кращими стануть генотипи з двома мутаціями. «Безпомилковий» генотип вже ніколи не відновиться. Таке безперервне погіршення клональних геномів змушує згадати про гвинтик — таке колесо, через яке наручники можуть лише затягуватись, а не розслаблятись.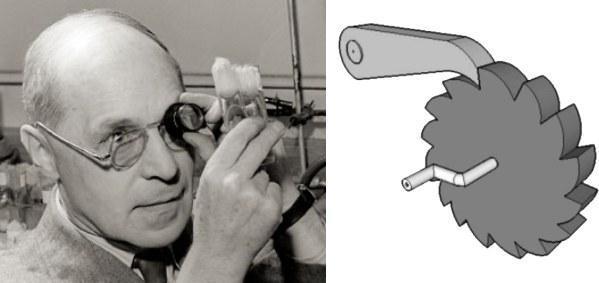
Зліва — Герман Джозеф Меллер (1890–1967). Справа — гвинтик: зубчасте колесо, яке може обертатися лише в один бік (у показаному на рисунку випадку — за годинниковою стрілкою)
Гвинтик Меллера, наприклад, відповідає за дегенерацію людської Y‑хромосоми, і причина цього саме в тому, що вона передається з покоління в покоління клонально. А гіпотеза, пов’язана з цим механізмом, полягає в тому, що саме він обмежує можливості існування клональних видів, створюючи переваги для статевих.
У результаті дії гвинтика Меллера подавляюча більшість видів, що відмовились від рекомбінації, по суті приречена. Ті, що мають відносно невеликий геном, високу чисельність і перебувають під ефективним контролем відбору, вимеруть відносно повільніше, але їхні перспективи все одно сумні. Прогресивну еволюцію до появи статевого розмноження забезпечували інші форми рекомбінації, а після виникнення статі розвиток і просто збереження працездатного генотипу стало пов’язане з нею. Оцініть самі: при схрещуванні організмів, кожен з яких несе по одній несприятливій мутації, чверть їхніх нащадків не успадкує жодної з них!
Гіпотеза Нанні — це розвиток ідеї гвинтика Меллера. L. Nannie припустив, що всі види зі статевим розмноженням, у еволюції яких виникали безстатеві клони, були витіснені цими клонами і вимерли, а пізніше (внаслідок дії гвинтика Меллера) вимерли і ці клони. Тому в результаті групового відбору залишились лише такі види зі статевим розмноженням, які з якоїсь причини втратили можливість перейти до клонального розмноження.
А як же відмовляються від статевого процесу дафнії, про які я розповідав у минулій колонці? З точки зору гіпотези Нанні вони — на півдорозі до вимирання...
"Топір Кондрашова" — мутаційна детерміністична гіпотеза Кондрашова. Висунута російським (зараз російсько‑американським) біологом Олексієм Симоновичем Кондрашовим. Ця гіпотеза звертає увагу на те, як слабко шкідливі окремо мутації розподіляються в популяції організмів зі статевим розмноженням. Деякі особи несуть невелику кількість таких мутацій, інші — багато. Припустимо, що дія цих мутацій має накопичувальний ефект: чим їх більше, тим нижча життєздатність. Розглянемо розподіл особин за кількістю несених ними мутацій. Відбір буде відсікати, як топором, найбільш навантажені класи особин, просто видаляючи «хвіст» розподілу. Кожна смерть особини з цих класів розподілу уносить з популяційного генофонду безліч мутацій; у безстатевих організмів таке ж очищення вимагало б загибелі багатьох особин.
Крім того, Кондрашов звернув увагу на еволюційні наслідки чергування гаплоїдності і диплоїдності, про які ми говорили минулого разу. Летальна мутація у гаплоїдного організму призводить до його смерті. Диплоїдний організм може в обох своїх хромосомних наборах мати кілька летальних мутацій і прекрасно себе почувати. З іншого боку, ті гени, які на гаплоїдній фазі призводять до загибелі (або окремого гаплоїдного покоління, чи хоча б гаплоїдних статевих клітин), будуть ефективно усуватись з популяції, не доходячи до диплоїдної фази.
На відміну від «вікаріума з Брея» та гвинтика Меллера, гіпотеза Кондрашова стосується короткострокових переваг організмів, що виникли в результаті статевого процесу.
Гіпотеза статевого відбору Меннінга. Англійський біолог John Manning звертає увагу на те, що самки, обираючи самців, можуть проводити досить ефективну оцінку пристосованості самців. Це продемонстровано і для людини, і для багатьох інших тварин — зокрема тих, що характеризуються складною поведінкою. Сумніватися в тому, що цей механізм у багатьох випадках працює, не доводиться. З іншого боку, цією гіпотезою не пояснити перехід до статевого відтворення від клонального, коли тонкі механізми статевого відбору ще не виникли.
"Еволюційну теорію статі" Геодакяна я назвав би інформаційною гіпотезою Геодакяна. Російський фізик і генетик Віген Геодакян висунув ряд оригінальних ідей, що стосуються того, як інформація про те, які генотипічні особливості особин виявляються затребуваними середовищем, відображається у генофонді виду. Він переконливо показує, що еволюційні нововведення випробовуються перш за все на самцях (на представниках того статі, якого менше жалко). Звичайно, про погляди Геодакяна треба писати окремо — і бажано докладно. Тут скажу лише, що, на мій погляд, у нього є цілий ряд хороших ідей, але й не мало явно сумнівних суджень. І, звичайно, я не можу погодитися з його інтегральною оцінкою результату його творчості.
"Широта теорії, її пояснювальний і предиктивний потенціал дозволяють їй зайняти місце багатьох часткових і недосконалих теорій у галузі статі, зокрема теорії статевого відбору Ч. Дарвіна". V. Geodakyan. Two Sexes. Why and Why Not. – s. 5.
У цю колонку вмістяться ще лише дві екологічні гіпотези, що стосуються короткострокових екологічних переваг організмів, що виникли під час статевого розмноження.
Гіпотеза лотереї Уільямса подобається мені особливо. З простого приводу: я раніше сам до неї дійшов — і лише потім прочитав про неї у Рідлі та Геодакяна. Я думав про брекетинг експозиції у фотографії. Коли фотограф не знає, чи підходить встановлена ним експозиція до наявних умов освітлення і особливостей об’єкта зйомки, а також коли йому не потрібно берегти кожен кадр, він може використовувати брекетинг. Один кадр автоматично робиться з тими налаштуваннями, які встановлені в камері, один — з підвищеною на крок експозицією, один — зі зниженою. Експозицію можна лише збільшувати або зменшувати, а для зміни властивостей нащадків існує безліч ступенів свободи. Що треба змінювати? Ті ознаки, за якими батько (пройшовший горнило відбору) відрізняється від своїх сородичів, також «перевірені» природним відбором.
На перший погляд, гіпотеза Джорджа Вільямса добре узгоджується з аналізом життєвих циклів. Поки умови залишаються постійними, дафнії і тлі розмножуються клонально. Коли умови змінюються, з’являється статне покоління, що підвищує різноманітність нащадків. Коли земляничний кущ або колонія коралових поліпів освоюють найближче оточення, вони роблять це клонально: земляниця викидає вуса, а корали розпускаються. Однак віддалені місця оселяються за допомогою насіння або личинок, що утворюються при статевому розмноженні. Організми, які шукають місця для своїх нащадків лише далеко (як в'їзди чи устриці), обходяться виключно статевим розмноженням.
На жаль, вважається, що аналіз поширення клональних і безстатевих форм спростовує гіпотезу Вільямса. За її словами, можна було б припустити, що статеве розмноження буде ефективніше в суворій і змінній середовищі, а безстатеве — у сприятливій і стабільній. Водночас відомо, що до безстатевого розмноження частіше переходять види, що населяють екстремальні місця.
Гіпотеза зарослого берега Белла — результат розвитку гіпотези лотереї. Грем Белл виходив із того, що статеве розмноження вигідно там, де надмірна кількість нащадків конкурує перш за все між собою. Чим більш схожі один на одного особи, тим гостріша конкуренція між ними; місткість середовища для ідентичних організмів нижча, ніж для різноманітних. Назва цієї гіпотези відсилає до знаменитого фінального пасажу «Походження видів» Дарвіна.
"Цікаво споглядати густо зарослий берег, вкритий численними, різноманітними рослинами з співаючими в кущах птахами, порхаючими навколо комах, що повзають у вологій землі черв’яками, і думати, що всі ці прекрасно побудовані форми, так різні одна від одної і так складно взаємозалежні, були створені законами, що й досі діють навколо нас". Charles Darwin. On the Origin of Species
Проте гіпотеза зарослого берега погано узгоджується з тим, що найбільші і найвдосконаліші організми, часто продукуючі небагато нащадків, розмножуються майже виключно статевим шляхом.
Серед гіпотез, що пояснюють переваги статевого розмноження, я не згадав лише одну — ту, яку вважають фавориткою багатьох авторитетних авторів. Це гіпотеза Червоного Короля Ван Валенa — Гамілтона. Про неї треба розповідати докладно, і в цю колонку її обговорення не вміститься. Що ж — наступного разу. Подивимося, чи зможе вона пояснити різке підвищення виживаності нащадків удвічі...
←
Dmytro Shabanov
→
Королева проблем еволюційної біології: кошмар Дженкіна повертається
Чи вистачить восьми непрямих гіпотез, що описують переваги статевого розмноження, щоб пояснити виникнення статі?
Правда, що королева еволюційних проблем — це Червона (a.k.a. Чорна) Королева? Не поспішайте з відповіддю — не забувайте про гермафродитів!
Колонка для Комп’ютерри #132
Колонка для Комп’ютерри #133
Колонка для Комп’ютерри #134