Чи правда, що королева еволюційних проблем — це Червона (a.k.a. Чорна) Королева? Не поспішайте з відповіддю — не забувайте про гермафродитів! Колонка для Комп'ютерри #134
А тепер я маю зробити визнання. У мене є своя версія розв’язку, просто я не поспішав її пояснювати. Щоб ви оцінили вихід із теоретичного тупика, вас треба було в цей тупик завести. Проникливий Nick Lane не розділяє секс і його «подвійну ціну». Він не правий. Зниження еф...
←
Dmytro Shabanov
→
Чи достатньо вісім непрямих гіпотез, що описують переваги статевого розмноження, щоб пояснити виникнення статі?
Чи правда, що королева еволюційних проблем — це Червона (a.k.a. Чорна) Королева? Не поспішайте з відповіддю — не забувайте про гермафродитів!
Перемога стійкості над оптимальністю, або Чому гермафродити програють самцям і самкам
Колонка для Комп’ютерри #133
Колонка для Комп’ютерри #134
Колонка для Комп’ютерри #135
Навіть нам, людям, зі всім нашим вмінням передбачати події, не так‑то просто діяти в інтересах наших власних найближчих нащадків: згадаємо знищення лісів, глобальне потепління та проблему перенаселення. Як же тоді еволюція, зі своєю сліпотою і егоїзмом, могла поставити довгострокову вигоду сексу для популяції вище його короткострокової подвійної ціни зі всіма наслідками?
Nick Lane. Сходи життя
Я зібрався досить докладно обговорити проблему еволюції популяційного відтворення і рекомбінації (включаючи феномен статі). Звісно, усі ключові думки можна стисло викласти в одному короткому тексті, але при цьому вони виявляться декларативними і незрозумілими. Щоб їх обґрунтувати, потрібен текст, обсягом що перевищує цілу колонку, проте таке обґрунтування виявиться незрозумілим для неспеціалістів. Я намагаюся викласти і складний комплекс загальновідомих ідей, і власні наработки так, щоб вони були доступними для розуміння вдумливих читачів‑небіологів. Як спонукати таких читачів розбиратися в обговорюваному предметі? Для мене самого часто виявляється цікавим матеріал про незнайомі мені галузі пізнання, у якому видно логіку міркувань розвиваючих ці галузі людей. Ось таку логіку я і намагаюся, в міру свого вміння і розуміння матеріалу, відобразити в цих колонках. Звісно, необхідний для цього обсяг тексту зростає, як на дріжджах. Ця колонка — вже п’ята з «статевої» серії; буде ще три чи чотири.
Перша була присвячена класифікації типів популяційного відтворення. У другій я коротко описав проблему еволюції рекомбінації і запропонував задуматися над рівнем відбору, що рухав цю еволюцію. У третій я спробував підштовхнути читачів до розуміння всієї парадоксальності статевого розмноження і почав обговорювати гіпотези, що пояснюють переваги статевого розмноження. Огляд цих гіпотез був завершений у четвертій колонці. Та гіпотеза, яку можна вважати фавориткою, не вмістилася і в четвертій, тому я обговорю її тут. Поговоривши про неї, ми спробуємо зрозуміти, чи закриває вона проблему чи ні.
Гіпотеза Червоного (also known as Чорного) Королеви Ван Валена і Гамільтона останнім часом стала найпопулярнішим поясненням феномена статі. Її назва — відсилання до класичного тексту Керролла. У вихідному тексті XIX століття діяла Червона Королева (бо в тодішніх шахах білі бились з червоними, а не з чорними). Сьогодні, перекладаючи «Алісу» на російську, Королеву зазвичай називають Чорною.
"— У нас, — сказала Аліса, з трудом переводячи подих, — коли довго біжиш усіма ногами, неодмінно потрапиш в інше місце.
— Яка повільна країна! — сказала Королева. — Ну, а тут, знаєш, доводиться бігти усіма ногами, щоб лише залишитися на тому ж місці! Якщо ж хочеш потрапити в інше місце, тоді треба бігти принаймні вдвічі швидше!" (Льюїс Керролл, «Аліса в Країні чудес»).
Американський еволюціоніст Lee Van Valen сформулював принцип Червоного Королеви, намагаючись пояснити закономірності вимирання організмів. Вільям Гамільтон застосував той самий принцип для пояснення переваг статевого розмноження.
Часто, описуючи історію якоїсь еволюційної гілки або життя в цілому, ми розглядаємо їх як шлях від досягнення до досягнення. План будови, характерний для риб, виник колись і існує до сьогодні. Деякі зміни цього плану дозволили освоїти сушу. Потім перестройка ембріонального розвитку дала можливість забезпечити наземний розвиток яйця...
У цих і багатьох інших випадках еволюційне набуття, досягнуте на певному етапі, у майбутньому зберігається в арсеналі розглянутої групи. Це пов’язано з тим, що, загалом, з часів виходу риб на сушу вода залишилася водою, а суша — сушею (хоча насправді і вода, і суша досить сильно змінилися). Проте біотична, жива середа змінюється просто швидко. Тут неможливо вирішити задачу пристосування до якихось умов раз і назавжди. Тільки хижак пристосується ловити і поїдати жертв — хапати, а жертви вже змінилися, і треба розробляти нові прийоми! Ледь господар навчився захищатися від атак паразитів — гляди, паразити вже не ті, що вчора, і способи захисту, які працювали зовсім недавно, виявляються неефективними. Ось у цьому безперервно мінливому середовищі, навіть щоб залишитися на місці (зберегти свою роль в екосистемі), треба безперервно «бігти» (еволюціонувати).
Щодо статевого розмноження гіпотеза Червоного Королеви полягає в наступному. Між взаємодіючими видами, передусім між паразитами і їх господарями, відбувається безперервна еволюційна «гонка озброєнь», яка вимагає постійних змін. Оскільки статевий процес прискорює потенційну швидкість еволюції, це сприяє успіху організмів зі статевим розмноженням. Зверніть увагу: мова йде передусім про імунологічну еволюцію, прояви якої можуть бути непомітні для зовнішнього спостерігача!
Розглянемо цю гіпотезу з точки зору її «терміновості» (sensu Protopopov, тобто положення на осі «короткостроковість — довгостроковість»). Червона Королева описує відносно короткострокові переваги статевого розмноження, принаймні у порівнянні з шестернею Мюллера і «вікарієм з Брея». Справа в тому, що частиною цієї гіпотези є пояснення того, чому середовище, до якого доводиться пристосовуватись, змінюється швидко і вимагає швидких змін від організмів, що нас цікавлять. З іншого боку, описана Червоною Королевою перевага навряд чи проявиться у першому ж поколінні, чисельність якого доведеться уполовинити через «подвійну ціну статі».
Червона Королева — не просто умисний конструкт; ця гіпотеза отримала експериментальне підтвердження! Ось, прочитайте про це в новині Олени Наймарк на «Елементах».
У круглоголового черв’яка Caenorhabditis elegans, класичного об’єкта генетики розвитку, є два статі — самовисплідні гермафродити і самці.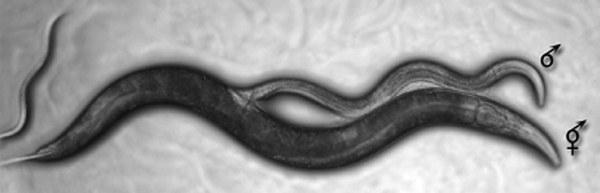
Спарювання самця і гермафродита Caenorhabditis elegans
Строго кажучи, розмноження гермафродитів у Caenorhabditis не клональне, оскільки їх потомство отримується від запліднення власних яйцеклітин власними сперматозоїдами. При утворенні статевих клітин відбувається випадкове скорочення генетичного матеріалу вдвічі. Оскільки за кожним з локусів (позицій у хромосомах) статеві клітини несуть лише половину алелей (версій певного гена) батька, то нащадки гермафродита за половину локусів отримають дві копії одного й того ж батьківського алеля. Після кількох циклів самозапліднення її нащадки стануть повністю гомозиготними (несучими за кожним локусом ідентичні алелі), і їх розмноження стане еквівалентним клональному (втрачати вже не буде чого).
Нащадки гермафродитів — гермафродити (і рідко‑рідко, через невідповідність статевих хромосом, — самці). Половина нащадків від схрещування самців і гермафродитів — самці, половина — гермафродити. Це спрощує оцінку частки схрещувань з рекомбінацією в популяції черв’їв: вона пропорційна частці самців.
Якщо черв’ї розвиваються в умовах, які не вимагають від них швидкої еволюції, гермафродити обігрують самців, адже їх швидкість розмноження виявляється вдвічі вищою (один вивідок нащадків залишає або кожен гермафродит, або дві особи — гермафродит + самець). Тому в стабільних умовах частка самців у популяції знижується до низького рівня.
Проте чим сильніша небезпека від загрозливих черв’їв‑паразитів, тим вище частка самців. Серед описаних у статті Наймарк експериментів найпереконливіший такий. Експериментатори заражали черв’їв у експериментальній популяції бактеріями, культуру яких на кожному етапі збирали на трупах їх співчотириків з попереднього покоління, погублених інфекцією. Це сприяло швидкій еволюції бактерій, що пристосовувалися подолати захисні механізми господарів. І в цих умовах частка самців (і, відповідно, частка нащадків, що несуть рекомбінаційні геноми) стабільно зростає.
"Ці результати беззастережно доводять, що низький рівень рекомбінації, що залежить від числа схрещуваних особин, безпосередньо визначає стійкість до інфекцій. За умов постійної присутності інфекційних агентів і їх безперервного розвитку безстатеве розмноження (самозапліднення) неминуче веде до вимирання. У цьому контексті паразитів можна замінити будь‑яким стресовим агентом, який змінюється з ходом часу. Очевидно, що таким агентом можуть бути і хижаки, і конкуренти, а також абіотичні фактори — вони рідко бувають постійними протягом кількох поколінь у тварин і рослин" (Олена Наймарк).
Схоже, ці результати допомагають знайти відповідь на балансний аргумент Уільямса, про який я писав у третій з «статевих» колонок. Ця проблема полягає у пошуку пояснення, чому популяції зі статевим розмноженням не витісняються клональними мутантами, які розмножуються вдвічі інтенсивніше. Виявляється, що, принаймні у деяких випадках, клональні лінії можуть бути знищені паразитами. Оптимально пристосувавшись до певного генотипу, паразити можуть викоренити всіх представників клону, що мають цей генотип, і інтенсивність розмноження клону не матиме в цій ситуації жодного значення: скільки з’явиться особин, стільки паразити їх і вб’ють. А про те, чому клони іноді все ж зберігаються, треба буде поговорити пізніше...
Але чи пояснюють ці експерименти походження статі? З тим, що Червона Королева пояснила феномен статі, згідні такі авторитети, як Alexander Markov або Matt Ridley. Проте, кажуть, Alexander Kondrashov (автор згаданого в попередній колонці топора Кондрашова) продовжує вважати феномен статі так і не отримавшим свого пояснення.
Експерименти, про які пише Наймарк, безсумнівно переконливі. Проте не забувайте: довести, що «сили» Червоного Королеви були б достатніми, щоб подолати подвійну падіння плодючості, викликану переходом від клональності до роздвоєної статі у часи виникнення статевого розмноження, вони не можуть.
У випадку Caenorhabditis elegans ми маємо справу з видом, у якого вже прекрасно сформовано статеве розмноження. Те, що у гермафродитів воно виродилося внаслідок самозапліднення, — вторинний ефект. До речі, аналізуючи описаний експеримент, треба мати на увазі, що нащадки, що утворилися в результаті самозапліднення, виявляються більш‑або‑менш гомозиготними (маючи ідентичні алелі), а черв’ї, що з’явилися від схрещування самців і гермафродитів, виявляються набагато більш гетерозиготними (за багатьма локусами мають різні версії гомологічних алелей). Підвищена гетерозиготність може сприяти зростанню стійкості і сама по собі, без урахування наслідків рекомбінації.
У будь‑якому випадку Caenorhabditis виробляє і яйцеклітини, і сперматозоїди. Гапло‑диплоїдний життєвий цикл з заплідненням і мейозом у цих черв’їв цілком сформований. Червона Королева може підвищити частку черв’їв зі статевим розвитком. Водночас я сумніваюся, що вона могла б забезпечити еволюцію від клонального розмноження до статевого, переступивши через двократну втрату ефективності відтворення.
А тепер я маю зробити визнання. Як я і писав ще у позапрошлій колонці, у мене є своя версія рішення, просто я не поспішав її роз’яснювати. Щоб ви оцінили вихід з теоретичного тупика, вас треба було в цей тупик завести. Я і в цій колонці не встигну дати всі обґрунтування рішення, але позначити його зможу.
Проникливий (крім жартів) Nick Lane, процитований в епіграфі, не розділяє секс і його «подвійну ціну». Він не правий. Зниження ефективності відтворення пов’язане не зі статевим розмноженням як таковим, а з наявністю самців, які самі не залишають нащадків. Те, що John Maynard Smith назвав «подвійною ціною статі», правильно було б назвати «подвійною ціною роздвоєної статі». Гермафродити не платять цю ціну!
Дозвольте нагадати ілюстрацію, яку я наводив, починаючи розмову про еволюцію рекомбінації.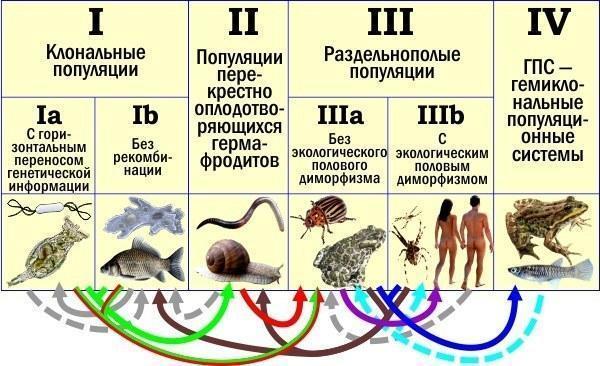
Класифікація типів популяційного відтворення та переходи між ними
Зверніть увагу на червону, зелену та червоно‑зелену стрілки. Ми досі говорили про «червоний‑зелений» перехід — I→IIIa. Але, як я писав, пояснюючи цю таблицю, я вважаю значно більш імовірним інший варіант: спочатку «зелений» виникнення статевого процесу у перехресно запліднюваних гермафродитів I→II, а потім уже «червоний» перехід до роздвоєної статі — II→IIIa.
Чому більшість авторів розглядає саме перехід I→IIIa? Традиція. Звичка вважати роздвоєну стать типічним випадком статевого розмноження. Інерція думки, передбачена балансним аргументом Уільямса. Нарешті, пояснити, як роздвоєна стать перемагає гермафродитизм — теж складне завдання...
Я з великою впевненістю припускаю, що статевий процес, а також гапло‑диплоїдний цикл з заплідненням і мейозом виникали у перехресно запліднюваних гермафродитах. Це вони перейшли від негомологічної рекомбінації в процесі горизонтального перенесення до упорядкованої гомологічної рекомбінації під час мейозу. Це вони зібрали механізм статевого розмноження з окремих раніше існуючих молекулярних блоків. І оскільки їм не доводилося платити подвійну ціну статі, падіння ефективності їх розмноження не блокувало цей вирішальний процес.
Пам’ятаєте, у попередній колонці я порівнював ефективність відтворення клональних і роздвоєних організмів? У випадку переходу I→IIIa, якщо перевага роздвоєних організмів була менша за двократну (!), клональні організми вигравали. Однак при переході I→II достатньо лише невеликої переваги гермафродитів. Дивіться.
«Змагання» 2
Тип
відтворення
Виробники
Нащадки
Виживаність
Виживе
Всього
Результат
Варіант A
Клональність
(тип I)
Особа 1
100
25%
25
50
Програш
Особа 2
100
25%
—
Варіант D
Перехресний
гермафродитизм
(тип II)
Партнер 1
100
26%
26
52
Перемога
Партнер 2
100
26%
—
Причини, що зумовили перехід від клонального розмноження до перехресного гермафродитизму, можна уявити досить добре. Вироблення нових пристосувань прискорювалося завдяки «вікарію з Брея» та берегу Белла, а дія Червоного Королеви робила цей процес безкінечним. Позбавлення від несприятливих мутацій полегшувалося завдяки виходу з‑під влади шестерні Мюллера і дії топора Кондрашова. Поширенню гермафродитів сприяла лотерея Уільямса. І жодної «подвійної ціни статі» Мейнарда Сміта платити не треба було.
Залишається одне питання. А як же гермафродити могли перейти до розподілу ролей самців і самок? Адже це вимагало сплати «подвійної ціни роздвоєної статі» — зниження ефективності відтворення вдвічі.
У мене є відповідь на це питання. Я дам її в наступній колонці.
←
Dmytro Shabanov
→
Чи достатньо вісім непрямих гіпотез, що описують переваги статевого розмноження, щоб пояснити виникнення статі?
Правда, що королева еволюційних проблем — це Червона (a.k.a. Чорна) Королева? Не поспішайте з відповіддю — не забувайте про гермафродитів!
Перемога стійкості над оптимальністю, або Чому гермафродити програють самцям і самкам
Колонка для Комп’ютерри #133
Колонка для Комп’ютерри #134
Колонка для Комп’ютерри #135