Is the Red (a.k.a. Black) Queen really the queen of evolutionary problems? Don't rush to answer — don't forget about hermaphrodites! Column for Kompyuterra #134
Now I must make a confession. I have my own version of the solution, I simply was not in a hurry to explain it. In order for you to appreciate the way out of the theoretical impasse, you needed to be led into that impasse first. The perspicacious Nick Lane does not separate sex and its «double price». He is wrong. The reduction of eff...
←
Dmytro Shabanov
→
Are eight indirect hypotheses describing the advantages of sexual reproduction sufficient to explain the origin of sex? Is it true that the queen of evolutionary problems is the Red (a.k.a. Black) Queen? Do not rush to answer - do not forget about hermaphrodites! The victory of persistence over optimality, or Why hermaphrodites lose to males and females
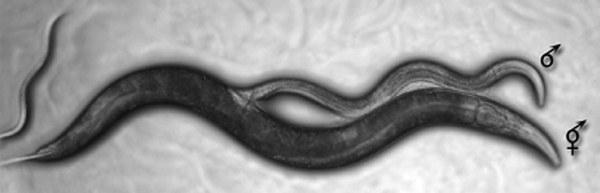
{"translation":"←\nDmytro Shabanov\n→\n\nWill eight indirect hypotheses describing the advantages of sexual reproduction suffice to explain the origin of sex?\nIs the Red (a.k.a. Black) Queen truly the queen of evolutionary problems? Don't rush to answer — don't forget about hermaphrodites!\nThe victory of stability over optimality, or Why hermaphrodites lose to males and females\n\nColumn for Kompyuterra #133\nColumn for Kompyuterra #134\nColumn for Kompyuterra #135\n\nEven for us humans, with all our ability to foresee events, it is not so simple to act in the interests of our own nearest descendants: let us recall deforestation, global warming, and the overpopulation problem. How then could evolution, with its blindness and selfishness, place the long-term benefit of sex for the population above its short-term double cost with all the unpleasant consequences that follow?\nNick Lane. The Tree of Life\n\nI set out to discuss in sufficient detail the problem of the evolution of population reproduction and recombination (including the sex phenomenon). Of course, all key ideas can be concisely outlined in one short text, but they would turn out to be declarative and incomprehensible. To substantiate them, a text exceeding the volume of an entire column would be necessary, but such substantiation would be incomprehensible to non-specialists. I am trying to present both the complex set of well-known ideas and my own developments in a way that would be accessible to understanding by thoughtful non-biologist readers. How can I encourage such readers to delve into the subject under discussion? For me personally, material about branches of knowledge unfamiliar to me, in which the logic of the people developing these branches is visible, is often interesting. It is precisely such logic that I am trying, to the best of my ability and understanding of the material, to reflect in these columns. Naturally, the volume of text necessary for this grows like yeast. This column is already the fifth in the \"sexual\" series; there will be three or more to come.\n\nThe first was devoted to the classification of types of population reproduction. In the second, I briefly described the problem of the evolution of recombination and suggested considering the level of selection that drove this evolution. In the third, I tried to push readers toward understanding the entire paradoxical nature of sexual reproduction and began discussing hypotheses explaining the advantages of sexual reproduction. The review of these hypotheses was completed in the fourth column. The hypothesis that can be considered the favorite did not fit in the fourth column either, so I will discuss it here. After talking about it, we will try to understand whether it resolves the problem or not.\n\nThe Red (also known as Black) Queen hypothesis of Van Valen and Hamilton has recently become the most popular explanation for the sex phenomenon. Its title is an allusion to Carroll's classic text. In the original 19th-century text, the Red Queen was in action (for in chess at that time, white fought against red, not black). Now, when translating \"Alice\" into Russian, the Queen is usually called Black.\n\"— Here, — said Alice, hardly able to speak, — when you run so fast for a long time, you are sure to get somewhere else.\n— What a slow country! — said the Queen. — Well, here, you know, you have to run as fast as you can just to stay in the same place! And if you want to get somewhere else, you must run at least twice as fast!\" (Lewis Carroll, \"Alice in Wonderland\").\n\nAmerican evolutionist Leigh Van Valen formulated the Red Queen principle while trying to explain the patterns of organism extinction. William Hamilton applied the same principle to explain the advantages of sexual reproduction.\n\nOften, when describing the history of any evolutionary branch or life in general, we consider them as a path from achievement to achievement. The body plan characteristic of fish arose at some point and persists to this day. Some modifications of this plan allowed conquest of the land. Then, the restructuring of embryonic development made it possible to provide terrestrial development of the egg...\n\nIn these and many other cases, the evolutionary acquisition achieved at a certain stage is preserved in the future in the arsenal of the group in question. This is because, generally speaking, since the time fish emerged onto land, water has remained water and land has remained land (although in reality both water and land have changed quite significantly). However, the biotic, living environment changes simply swiftly. Here it is impossible to solve the problem of adaptation to any conditions once and for all. Just as the predator adapts to catch and eat prey — snap, and the prey have already changed, and new techniques must be developed! As soon as the host learns to defend against parasite attacks — look, the parasites are no longer those of yesterday, and the defense methods that worked quite recently turn out to be ineffective. In this continuously changing environment, even to remain in place (to preserve one's role in the ecosystem), one must continuously \"run\" (evolve).\n\nRegarding sexual reproduction, the Red Queen hypothesis is as follows. Between interacting species, primarily between parasites and their hosts, there is a continuous evolutionary \"arms race\" that requires constant change. Since the sexual process accelerates the potential rate of evolution, this promotes the success of organisms with sexual reproduction. Note: the discussion here concerns primarily immunological evolution, the manifestations of which may be imperceptible to an external observer!\n\nLet us consider this hypothesis from the standpoint of its \"urgency\" (sensu Protopopov, i.e., its position on the \"short-term — long-term\" axis). The Red Queen describes relatively short-term advantages of sexual reproduction, at least compared to Muller's ratchet and the \"vicar of Bray.\" The thing is that part of this hypothesis is an explanation of why the environment to which adaptation is required changes rapidly and requires rapid changes from the organisms in question. On the other hand, the advantage described by the Red Queen will hardly manifest in the very first generation, whose numbers would have to be halved due to the \"double cost of sex.\"\n\nThe Red Queen is not merely a speculative construct; this hypothesis has received experimental confirmation! Read about this in Elena Naimark's news item on \"Elements.\"\n\nThe roundworm Caenorhabditis elegans, a classic model organism in developmental genetics, has two sexes — self-fertilizing hermaphrodites and males.\n[IMG_1]\nMating of a male and hermaphrodite Caenorhabditis elegans\n\nStrictly speaking, reproduction of hermaphrodites in Caenorhabditis is not clonal, since their offspring result from fertilization of their own eggs by their own sperm. During gamete formation, the genetic material is randomly reduced by half. Since for each locus (position in chromosomes) gametes carry only half of the parent's alleles (versions of a particular gene), the offspring of a hermaphrodite will receive two copies of the same parental allele for half of the loci. After several cycles of self-fertilization, its offspring will become completely homozygous (carrying identical alleles at each locus), and their reproduction will become equivalent to clonal (there will be nothing left to lose).\n\nThe offspring of hermaphrodites are hermaphrodites (and rarely-rarely, due to non-disjunction of sex chromosomes, — males). Half of the offspring from crossing males and hermaphrodites are males, half are hermaphrodites. This simplifies assessment of the proportion of crossings with recombination in the worm population: it is proportional to the proportion of males.\n\nIf worms develop under conditions that do not require rapid evolution from them, hermaphrodites outcompete males, since their reproductive rate turns out to be twice as high (either each hermaphrodite leaves one brood of offspring, or two individuals — hermaphrodite + male). Therefore, under stable conditions, the proportion of males in the population decreases to a low level.\n\nHowever, the greater the danger from parasites threatening the worms, the higher the proportion of males. Among the experiments described in Naimark's article, the most convincing is this one. The experimenters infected worms in the experimental population with bacteria, whose culture at each stage was collected from the corpses of their kin from the previous generation, killed by infection. This promoted the rapid evolution of bacteria, adapting to overcome the hosts' defense mechanisms. And under these conditions, the proportion of males (and, correspondingly, the proportion of offspring carrying recombined genomes) steadily increases.\n\n\"These results unequivocally prove that the low level of recombination, dependent on the number of crossing individuals, directly determines resistance to infections. Under conditions of the constant presence of infectious agents and their continuous development, asexual reproduction (self-fertilization) inevitably leads to extinction. In this context, parasites can be replaced by any stressful agent that changes over time. Clearly, such agents can be predators and competitors, as well as abiotic factors — they are rarely constant over a number of generations in animals and plants\" (Elena Naimark).\n\nIt seems these results help find an answer to Williams' balance argument, which I wrote about in the third of the \"sexual\" columns. This problem consists of finding an explanation for why populations with sexual reproduction are not displaced by clonal mutants, which reproduce twice as intensively. It turns out that, at least in some cases, clonal lineages can be destroyed by parasites. Having optimally adapted to a particular genotype, parasites can wipe out all representatives of the clone possessing this genotype, and the reproductive intensity of the clone will have no significance in this situation: however many individuals appear, the parasites will kill them all. And about why clones sometimes nevertheless persist, we will have to talk later...\n\nBut do these experiments explain the origin of sex? That the Red Queen has explained the sex phenomenon is agreed upon by such authorities as Alexander Markov or Matt Ridley. However, it is said that Alexander Kondrashov (the author of Kondrashov's axe, mentioned in the previous column) continues to consider the sex phenomenon as still lacking its explanation.\n\nThe experiments about which Naimark writes are undoubtedly convincing. However, don't forget: they cannot prove that the \"forces\" of the Red Queen would suffice to overcome the double drop in fertility caused by the transition from clonality to gonochorism at the time of the origin of sexual reproduction.\n\nIn the case of Caenorhabditis elegans, we are dealing with a species in which sexual reproduction is already perfectly formed. The fact that it is degenerate in hermaphrodites due to self-fertilization is a secondary effect. By the way, when analyzing the described experiment, one must bear in mind that offspring resulting from self-fertilization turn out to be more or less homozygous (having identical alleles), and worms appearing from crossing males and hermaphrodites turn out to be much more heterozygous (having different versions of homologous alleles at many loci). Increased heterozygosity can promote increased resistance by itself, without accounting for the consequences of recombination.\n\nIn any case, Caenorhabditis produces both eggs and sperm. The haploid-diploid life cycle with fertilization and meiosis is fully formed in these worms. The Red Queen can increase the proportion of worms with sexual development. At the same time, I doubt that it could ensure the evolution from clonal reproduction to sexual reproduction, stepping over the twofold loss in reproductive efficiency.\n\nAnd now I must make a confession. As I wrote back in the column before last, I have my own version of the solution, I just was not in a hurry to explain it. To appreciate the way out of the theoretical dead end, you needed to be led into this dead end. I also will not have time to give the full substantiation of the solution in this column, but I will be able to outline it.\n\nThe perceptive (no joke) Nick Lane, quoted in the epigraph, does not separate sex and its \"double cost.\" He is wrong. The decrease in reproductive efficiency is connected not with sexual reproduction as such, but with the presence of males, which themselves do not leave offspring. What John Maynard Smith called the \"double cost of sex\" would more correctly be called the \"double cost of gonochorism.\" Hermaphrodites do not pay this price!\n\nLet me remind you of the illustration I gave when starting the conversation about the evolution of recombination.\n[IMG_2]\nClassification of types of population reproduction and transitions between them\n\nPay attention to the red, green, and red-green arrows. We have been speaking all this time about the \"red-green\" transition — I→IIIa. But, as I wrote when explaining this table, I consider another variant much more probable: first the \"green\" emergence of the sexual process in cross-fertilizing hermaphrodites I→II, and then the \"red\" transition to gonochorism — II→IIIa.\n\nWhy do most authors consider precisely the transition I→IIIa? Tradition. The habit of considering gonochorism as the typical case of sexual reproduction. The inertia of thought, predetermined by Williams' balance argument. Finally, explaining how gonochorism defeats hermaphroditism is also a difficult task...\n\nI am quite confident that the sexual process, as well as the haploid-diploid cycle with fertilization and meiosis, arose in cross-fertilizing hermaphrodites. It was they who transitioned from non-homologous recombination during horizontal transfer to ordered homologous recombination during meiosis. It was they who assembled the mechanism of sexual reproduction from individual previously existing molecular blocks. And since they did not have to pay the double cost of sex, the drop in their reproductive efficiency did not block this fateful process.\n\nRemember, in the previous column I compared the reproductive efficiency of clonal and gonochorous organisms? In the case of transition I→IIIa, if the advantage of gonochorous organisms was less than twofold (!), clonal organisms won. Whereas in transition I→II, only a small advantage of hermaphrodites is sufficient. Look.\n\n\"Compe-\ntition\" 2\n\nType of repro-duction\n\nProducers\n\nOff-spring\n\nSur-vival\n\nWill survive\n\nTotal\n\nResult\n\nVariant A\n\nClonality\n(type I)\n\nIndividual 1\n\n100\n\n25%\n\n25\n\n50\n\nLoss\n\nIndividual 2\n\n100\n\n25\n\nVariant D\n\nCross hermaph-roditism\n(type II)\n\nPartner 1\n\n100\n\n26%\n\n26\n\n52\n\nVictory\n\nPartner 2\n\n100\n\n26\n\nThe reasons for the transition from clonal reproduction to cross-fermaphroditism can be imagined quite well. The development of new adaptations was accelerated thanks to the \"vicar of Bray\" and Bell's shore, and the action of the Red Queen made this process endless. Getting rid of deleterious mutations was facilitated thanks to escaping from the power of Muller's ratchet and the action of Kondrashov's axe. The spread of hermaphrodites was promoted by Williams' lottery. And there was no need to pay Maynard Smith's \"double cost of sex.\"\n\nOne question remains. But how could hermaphrodites transition to the separation of male and female roles? After all, this required payment of the \"double cost of gonochorism\" — a twofold decrease in reproductive efficiency.\n\nI have an answer to this question. I will give it in the next column.\n\n←\nDmytro Shabanov\n→\n\nWill eight indirect hypotheses describing the advantages of sexual reproduction suffice to explain the origin of sex?\nIs the Red (a.k.a. Black) Queen truly the queen of evolutionary problems? Don't rush to answer — don't forget about hermaphrodites!\nThe victory of stability over optimality, or Why hermaphrodites lose to males and females\n\nColumn for Kompyuterra #133\nColumn for Kompyuterra #134\nColumn for Kompyuterra #135"}
Even we humans, with all our ability to predict events, find it not so easy to act in the interests of our own immediate descendants: consider deforestation, global warming, and the problem of overpopulation. How then could evolution, with its blindness and selfishness, have prioritized the long-term benefit of sex for the population over its short-term double cost with all its consequences? Nick Lane. The Vital Question I intended to discuss the problem of the evolution of population reproduction and recombination (including the phenomenon of sex) in considerable detail. Of course, all the key ideas can be briefly stated in a single short text, but they will appear declarative and incomprehensible. To justify them, a text exceeding an entire column is needed, yet such a justification will be incomprehensible to non-specialists. I am trying to present both a complex set of well-known ideas and my own findings in a way that is accessible to thoughtful non-biologist readers. How to encourage such readers to delve into the subject? I myself often find material on unfamiliar fields of knowledge interesting, in which the logic of the reasoning of people developing these fields is evident. It is this logic that I am trying, to the best of my ability and understanding of the material, to reflect in these columns. Of course, the necessary text volume grows like yeast. This column is already the fifth in the "sexual" series; there will be three or four more. The first was devoted to the classification of types of population reproduction. In the second, I briefly described the problem of the evolution of recombination and suggested considering the level of selection that drove this evolution. In the third, I tried to push readers towards understanding the entire paradox of sexual reproduction and began to discuss hypotheses explaining the advantages of sexual reproduction. The review of these hypotheses was completed in the fourth column. The hypothesis that can be considered a favorite did not fit into the fourth either, so I will discuss it here. After discussing it, we will try to understand whether it closes the problem or not. The Red (also known as Black) Queen Hypothesis Van Valen and Hamilton have recently become the most popular explanation for the phenomenon of sex. Its name is a reference to Carroll's classic text. In the original 19th-century text, the Red Queen was active (because in chess at that time, white fought with red, not black). Today, when translating "Alice" into Russian, the Queen is usually called Black. "— It takes all the running you can do, to keep in the same place," said the Queen. "If you want to get somewhere else, you must run at least twice as fast as that!" (Lewis Carroll, "Alice's Adventures in Wonderland"). American evolutionist Lee Van Valen formulated the Red Queen principle while trying to explain the patterns of organism extinction. William Hamilton applied the same principle to explain the advantages of sexual reproduction. Often, when describing the history of an evolutionary branch or life as a whole, we view them as a path from achievement to achievement. The body plan characteristic of fish arose once and exists to this day. Some changes to this plan allowed for the colonization of land. Then, a restructuring of embryonic development made it possible to ensure terrestrial egg development... In these and many other cases, an evolutionary acquisition achieved at a certain stage is preserved in the arsenal of the group in question in the future. This is because, in general, since the time fish emerged onto land, water has remained water, and land has remained land (although in reality, both water and land have changed quite significantly). However, the biotic, living environment changes very rapidly. It is impossible to solve the problem of adaptation to certain conditions once and for all here. As soon as a predator adapts to catch and eat prey, the prey has already changed, and new tactics must be developed! As soon as the host learns to defend itself against parasite attacks, look, the parasites are no longer the same as yesterday, and the defense methods that worked very recently turn out to be ineffective. It is in this constantly changing environment that, even to stay in place (maintain one's role in the ecosystem), one must constantly "run" (evolve). Regarding sexual reproduction, the Red Queen hypothesis is as follows. Between interacting species, primarily between parasites and their hosts, there is a continuous evolutionary "arms race" that requires constant changes. Since sexual reproduction accelerates the potential rate of evolution, it contributes to the success of sexually reproducing organisms. Note: this primarily concerns immunological evolution, the manifestations of which may be imperceptible to an external observer! Let's consider this hypothesis from the perspective of its "urgency" (sensu Protopopov, i.e., its position on the "short-term-long-term" axis). The Red Queen describes relatively short-term advantages of sexual reproduction, at least compared to Müller's ratchet and the "viceroy of Bray." The fact is that part of this hypothesis is an explanation of why the environment to which adaptation is required is changing rapidly and demands rapid changes from the organisms of interest. On the other hand, the advantage described by the Red Queen is unlikely to manifest in the very first generation, whose numbers will have to be halved due to the "double cost of sex." The Red Queen is not just an intentional construct; this hypothesis has received experimental confirmation! Read about it in Elena Naimark's news on "Elementy." The roundworm Caenorhabditis elegans, a classic object of developmental genetics, has two sexes: self-fertilizing hermaphrodites and males. Mating of a male and a hermaphrodite Caenorhabditis elegans Strictly speaking, hermaphrodite reproduction in Caenorhabditis is not clonal, as their offspring are produced from the fertilization of their own egg cells by their own sperm. During the formation of gametes, the genetic material is randomly halved. Since gametes carry only half of the parent's alleles (versions of a particular gene) for each locus (position in the chromosomes), the offspring of a hermaphrodite will receive two copies of the same parental allele for half of the loci. After several cycles of self-fertilization, its offspring will become completely homozygous (carrying identical alleles for each locus), and their reproduction will become equivalent to clonal (there will be nothing left to lose). The offspring of hermaphrodites are hermaphrodites (and very rarely, due to sex chromosome mismatch, males). Half of the offspring from the cross of males and hermaphrodites are males, and half are hermaphrodites. This simplifies the estimation of the proportion of crosses with recombination in the worm population: it is proportional to the proportion of males. If worms develop in conditions that do not require rapid evolution, hermaphrodites outperform males, as their reproduction rate is twice as high (one brood of offspring is left by either each hermaphrodite, or two individuals - a hermaphrodite + a male). Therefore, in stable conditions, the proportion of males in the population decreases to a low level. However, the greater the danger from threatening parasitic worms, the higher the proportion of males. Among the experiments described in Naimark's article, the most convincing is the following. Experimenters infected worms in an experimental population with bacteria, the culture of which was collected at each stage from the corpses of their conspecifics from the previous generation, destroyed by infection. This promoted the rapid evolution of bacteria that adapted to overcome the host's defense mechanisms. And in these conditions, the proportion of males (and, consequently, the proportion of offspring carrying recombinant genomes) steadily increases. "These results unequivocally prove that a low level of recombination, dependent on the number of individuals mating, directly determines resistance to infections. Under conditions of constant presence of infectious agents and their continuous evolution, asexual reproduction (self-fertilization) inevitably leads to extinction. In this context, parasites can be replaced by any stress agent that changes over time. It is obvious that such an agent can be predators, competitors, as well as abiotic factors - they are rarely constant for several generations in animals and plants" (Elena Naimark). It seems these results help answer Williams's balancing argument, which I wrote about in the third "sexual" column. This problem lies in finding an explanation for why sexually reproducing populations are not displaced by clonal mutants that reproduce twice as fast. It turns out that, at least in some cases, clonal lines can be destroyed by parasites. Having optimally adapted to a specific genotype, parasites can eradicate all representatives of the clone with this genotype, and the reproduction rate of the clone will be irrelevant in this situation: no matter how many individuals appear, the parasites will kill them. As for why clones sometimes still survive, we will have to talk about that later... But do these experiments explain the origin of sex? Authorities like Alexander Markov or Matt Ridley agree that the Red Queen explains the phenomenon of sex. However, Alexander Kondrashov (author of the "Kondrashov's axe" mentioned in the previous column) reportedly continues to believe that the phenomenon of sex has not yet been explained. The experiments that Naimark writes about are undoubtedly convincing. However, do not forget: they cannot prove that the "forces" of the Red Queen were sufficient to overcome the double decline in fertility caused by the transition from clonality to gonochorism at the time of the origin of sexual reproduction. In the case of Caenorhabditis elegans, we are dealing with a species that already has well-developed sexual reproduction. The fact that it has degenerated into self-fertilization in hermaphrodites is a secondary effect. By the way, when analyzing the described experiment, it should be borne in mind that offspring produced by self-fertilization are more or less homozygous (having identical alleles), while worms resulting from the cross of males and hermaphrodites are much more heterozygous (having different versions of homologous alleles at many loci). Increased heterozygosity can contribute to increased resistance on its own, without considering the effects of recombination. In any case, Caenorhabditis produces both eggs and sperm. The haplo-diploid life cycle with fertilization and meiosis is fully formed in these worms. The Red Queen can increase the proportion of worms with sexual development. At the same time, I doubt that it could have ensured the evolution from clonal reproduction to sexual reproduction, overcoming the twofold loss of reproductive efficiency. And now I must make a confession. As I wrote in the column before last, I have my own version of the solution, I just wasn't in a hurry to explain it. To appreciate the way out of the theoretical impasse, you had to be led into that impasse. Even in this column, I won't have time to provide all the justifications for the solution, but I will be able to outline it. The insightful (no joke) Nick Lane, quoted in the epigraph, does not distinguish between sex and its "double cost." He is wrong. The decrease in reproductive efficiency is not related to sexual reproduction as such, but to the presence of males who themselves do not leave offspring. What John Maynard Smith called the "double cost of sex" would be more correctly called the "double cost of gonochorism." Hermaphrodites do not pay this price! Let me remind you of the illustration I used when starting the discussion about the evolution of recombination. Classification of types of population reproduction and transitions between them Note the red, green, and red-green arrows. We have been talking about the "red-green" transition – I→IIIa. But, as I wrote when explaining this table, I consider another option much more likely: first, the "green" origin of sexual reproduction in cross-fertilizing hermaphrodites I→II, and then the "red" transition to gonochorism – II→IIIa. Why do most authors consider the transition I→IIIa? Tradition. The habit of considering gonochorism as the typical case of sexual reproduction. Inertia of thought, predicted by Williams's balancing argument. Finally, explaining how gonochorism overcomes hermaphroditism is also a difficult task... I assume with great confidence that sexual reproduction, as well as the haplo-diploid cycle with fertilization and meiosis, arose in cross-fertilizing hermaphrodites. It was they who transitioned from non-homologous recombination during horizontal gene transfer to ordered homologous recombination during meiosis. It was they who assembled the mechanism of sexual reproduction from previously existing molecular blocks. And since they did not have to pay the double cost of sex, the decline in their reproductive efficiency did not block this crucial process. Remember, in the previous column, I compared the reproductive efficiency of clonal and gonochoric organisms? In the case of the transition I→IIIa, if the advantage of gonochoric organisms was less than twofold (!), clonal organisms won. However, in the transition I→II, only a small advantage for hermaphrodites is sufficient. Look.
"Competition" 2
Mode of reproduction
Producers
Descendants
Survival rate
Will survive
Total
Result
Variant A
Clonality (Type I)
Person 1
100
25%
25
50
Loss
Person 2
100
25%
—
Variant D
Cross-fertilization (Type II)
Partner 1
100
26%
26
52
Victory
Partner 2
100
26%
—
Причини, що зумовили перехід від клонального розмноження до перехресного гермафродитизму, можна уявити досить добре. Вироблення нових пристосувань прискорювалося завдяки «вікарію з Брея» та берегу Белла, а дія Червоного Королеви робила цей процес безкінечним. Позбавлення від несприятливих мутацій полегшувалося завдяки виходу з‑під влади шестерні Мёллера і дії топора Кондрашова. Поширенню гермафродитів сприяла лотерея Уільямса. І жодної «подвійної ціни статі» Мейнарда Сміта платити не треба було.
Залишається одне питання. А як же гермафродити могли перейти до розподілу ролей самців і самок? Адже це вимагало сплати «подвійної ціни роздвоєної статі» — зниження ефективності відтворення вдвічі.
У мене є відповідь на це питання. Я дам її в наступній колонці.
←
Dmytro Shabanov
→
Are the eight indirect hypotheses describing the advantages of sexual reproduction sufficient to explain the origin of sex? Is it true that the queen of evolutionary problems is the Red (a.k.a. Black) Queen? Don't rush to answer – don't forget about hermaphrodites! The victory of stability over optimality, or Why hermaphrodites lose to males and females
{"translation":"←\nDmytro Shabanov\n→\n\nWill eight indirect hypotheses describing the advantages of sexual reproduction suffice to explain the origin of sex?\nIs the Red (a.k.a. Black) Queen truly the queen of evolutionary problems? Don't rush to answer — don't forget about hermaphrodites!\nThe victory of stability over optimality, or Why hermaphrodites lose to males and females\n\nColumn for Kompyuterra #133\nColumn for Kompyuterra #134\nColumn for Kompyuterra #135\n\nEven for us humans, with all our ability to foresee events, it is not so simple to act in the interests of our own nearest descendants: let us recall deforestation, global warming, and the overpopulation problem. How then could evolution, with its blindness and selfishness, place the long-term benefit of sex for the population above its short-term double cost with all the unpleasant consequences that follow?\nNick Lane. The Tree of Life\n\nI set out to discuss in sufficient detail the problem of the evolution of population reproduction and recombination (including the sex phenomenon). Of course, all key ideas can be concisely outlined in one short text, but they would turn out to be declarative and incomprehensible. To substantiate them, a text exceeding the volume of an entire column would be necessary, but such substantiation would be incomprehensible to non-specialists. I am trying to present both the complex set of well-known ideas and my own developments in a way that would be accessible to understanding by thoughtful non-biologist readers. How can I encourage such readers to delve into the subject under discussion? For me personally, material about branches of knowledge unfamiliar to me, in which the logic of the people developing these branches is visible, is often interesting. It is precisely such logic that I am trying, to the best of my ability and understanding of the material, to reflect in these columns. Naturally, the volume of text necessary for this grows like yeast. This column is already the fifth in the \"sexual\" series; there will be three or more to come.\n\nThe first was devoted to the classification of types of population reproduction. In the second, I briefly described the problem of the evolution of recombination and suggested considering the level of selection that drove this evolution. In the third, I tried to push readers toward understanding the entire paradoxical nature of sexual reproduction and began discussing hypotheses explaining the advantages of sexual reproduction. The review of these hypotheses was completed in the fourth column. The hypothesis that can be considered the favorite did not fit in the fourth column either, so I will discuss it here. After talking about it, we will try to understand whether it resolves the problem or not.\n\nThe Red (also known as Black) Queen hypothesis of Van Valen and Hamilton has recently become the most popular explanation for the sex phenomenon. Its title is an allusion to Carroll's classic text. In the original 19th-century text, the Red Queen was in action (for in chess at that time, white fought against red, not black). Now, when translating \"Alice\" into Russian, the Queen is usually called Black.\n\"— Here, — said Alice, hardly able to speak, — when you run so fast for a long time, you are sure to get somewhere else.\n— What a slow country! — said the Queen. — Well, here, you know, you have to run as fast as you can just to stay in the same place! And if you want to get somewhere else, you must run at least twice as fast!\" (Lewis Carroll, \"Alice in Wonderland\").\n\nAmerican evolutionist Leigh Van Valen formulated the Red Queen principle while trying to explain the patterns of organism extinction. William Hamilton applied the same principle to explain the advantages of sexual reproduction.\n\nOften, when describing the history of any evolutionary branch or life in general, we consider them as a path from achievement to achievement. The body plan characteristic of fish arose at some point and persists to this day. Some modifications of this plan allowed conquest of the land. Then, the restructuring of embryonic development made it possible to provide terrestrial development of the egg...\n\nIn these and many other cases, the evolutionary acquisition achieved at a certain stage is preserved in the future in the arsenal of the group in question. This is because, generally speaking, since the time fish emerged onto land, water has remained water and land has remained land (although in reality both water and land have changed quite significantly). However, the biotic, living environment changes simply swiftly. Here it is impossible to solve the problem of adaptation to any conditions once and for all. Just as the predator adapts to catch and eat prey — snap, and the prey have already changed, and new techniques must be developed! As soon as the host learns to defend against parasite attacks — look, the parasites are no longer those of yesterday, and the defense methods that worked quite recently turn out to be ineffective. In this continuously changing environment, even to remain in place (to preserve one's role in the ecosystem), one must continuously \"run\" (evolve).\n\nRegarding sexual reproduction, the Red Queen hypothesis is as follows. Between interacting species, primarily between parasites and their hosts, there is a continuous evolutionary \"arms race\" that requires constant change. Since the sexual process accelerates the potential rate of evolution, this promotes the success of organisms with sexual reproduction. Note: the discussion here concerns primarily immunological evolution, the manifestations of which may be imperceptible to an external observer!\n\nLet us consider this hypothesis from the standpoint of its \"urgency\" (sensu Protopopov, i.e., its position on the \"short-term — long-term\" axis). The Red Queen describes relatively short-term advantages of sexual reproduction, at least compared to Muller's ratchet and the \"vicar of Bray.\" The thing is that part of this hypothesis is an explanation of why the environment to which adaptation is required changes rapidly and requires rapid changes from the organisms in question. On the other hand, the advantage described by the Red Queen will hardly manifest in the very first generation, whose numbers would have to be halved due to the \"double cost of sex.\"\n\nThe Red Queen is not merely a speculative construct; this hypothesis has received experimental confirmation! Read about this in Elena Naimark's news item on \"Elements.\"\n\nThe roundworm Caenorhabditis elegans, a classic model organism in developmental genetics, has two sexes — self-fertilizing hermaphrodites and males.\n[IMG_1]\nMating of a male and hermaphrodite Caenorhabditis elegans\n\nStrictly speaking, reproduction of hermaphrodites in Caenorhabditis is not clonal, since their offspring result from fertilization of their own eggs by their own sperm. During gamete formation, the genetic material is randomly reduced by half. Since for each locus (position in chromosomes) gametes carry only half of the parent's alleles (versions of a particular gene), the offspring of a hermaphrodite will receive two copies of the same parental allele for half of the loci. After several cycles of self-fertilization, its offspring will become completely homozygous (carrying identical alleles at each locus), and their reproduction will become equivalent to clonal (there will be nothing left to lose).\n\nThe offspring of hermaphrodites are hermaphrodites (and rarely-rarely, due to non-disjunction of sex chromosomes, — males). Half of the offspring from crossing males and hermaphrodites are males, half are hermaphrodites. This simplifies assessment of the proportion of crossings with recombination in the worm population: it is proportional to the proportion of males.\n\nIf worms develop under conditions that do not require rapid evolution from them, hermaphrodites outcompete males, since their reproductive rate turns out to be twice as high (either each hermaphrodite leaves one brood of offspring, or two individuals — hermaphrodite + male). Therefore, under stable conditions, the proportion of males in the population decreases to a low level.\n\nHowever, the greater the danger from parasites threatening the worms, the higher the proportion of males. Among the experiments described in Naimark's article, the most convincing is this one. The experimenters infected worms in the experimental population with bacteria, whose culture at each stage was collected from the corpses of their kin from the previous generation, killed by infection. This promoted the rapid evolution of bacteria, adapting to overcome the hosts' defense mechanisms. And under these conditions, the proportion of males (and, correspondingly, the proportion of offspring carrying recombined genomes) steadily increases.\n\n\"These results unequivocally prove that the low level of recombination, dependent on the number of crossing individuals, directly determines resistance to infections. Under conditions of the constant presence of infectious agents and their continuous development, asexual reproduction (self-fertilization) inevitably leads to extinction. In this context, parasites can be replaced by any stressful agent that changes over time. Clearly, such agents can be predators and competitors, as well as abiotic factors — they are rarely constant over a number of generations in animals and plants\" (Elena Naimark).\n\nIt seems these results help find an answer to Williams' balance argument, which I wrote about in the third of the \"sexual\" columns. This problem consists of finding an explanation for why populations with sexual reproduction are not displaced by clonal mutants, which reproduce twice as intensively. It turns out that, at least in some cases, clonal lineages can be destroyed by parasites. Having optimally adapted to a particular genotype, parasites can wipe out all representatives of the clone possessing this genotype, and the reproductive intensity of the clone will have no significance in this situation: however many individuals appear, the parasites will kill them all. And about why clones sometimes nevertheless persist, we will have to talk later...\n\nBut do these experiments explain the origin of sex? That the Red Queen has explained the sex phenomenon is agreed upon by such authorities as Alexander Markov or Matt Ridley. However, it is said that Alexander Kondrashov (the author of Kondrashov's axe, mentioned in the previous column) continues to consider the sex phenomenon as still lacking its explanation.\n\nThe experiments about which Naimark writes are undoubtedly convincing. However, don't forget: they cannot prove that the \"forces\" of the Red Queen would suffice to overcome the double drop in fertility caused by the transition from clonality to gonochorism at the time of the origin of sexual reproduction.\n\nIn the case of Caenorhabditis elegans, we are dealing with a species in which sexual reproduction is already perfectly formed. The fact that it is degenerate in hermaphrodites due to self-fertilization is a secondary effect. By the way, when analyzing the described experiment, one must bear in mind that offspring resulting from self-fertilization turn out to be more or less homozygous (having identical alleles), and worms appearing from crossing males and hermaphrodites turn out to be much more heterozygous (having different versions of homologous alleles at many loci). Increased heterozygosity can promote increased resistance by itself, without accounting for the consequences of recombination.\n\nIn any case, Caenorhabditis produces both eggs and sperm. The haploid-diploid life cycle with fertilization and meiosis is fully formed in these worms. The Red Queen can increase the proportion of worms with sexual development. At the same time, I doubt that it could ensure the evolution from clonal reproduction to sexual reproduction, stepping over the twofold loss in reproductive efficiency.\n\nAnd now I must make a confession. As I wrote back in the column before last, I have my own version of the solution, I just was not in a hurry to explain it. To appreciate the way out of the theoretical dead end, you needed to be led into this dead end. I also will not have time to give the full substantiation of the solution in this column, but I will be able to outline it.\n\nThe perceptive (no joke) Nick Lane, quoted in the epigraph, does not separate sex and its \"double cost.\" He is wrong. The decrease in reproductive efficiency is connected not with sexual reproduction as such, but with the presence of males, which themselves do not leave offspring. What John Maynard Smith called the \"double cost of sex\" would more correctly be called the \"double cost of gonochorism.\" Hermaphrodites do not pay this price!\n\nLet me remind you of the illustration I gave when starting the conversation about the evolution of recombination.\n[IMG_2]\nClassification of types of population reproduction and transitions between them\n\nPay attention to the red, green, and red-green arrows. We have been speaking all this time about the \"red-green\" transition — I→IIIa. But, as I wrote when explaining this table, I consider another variant much more probable: first the \"green\" emergence of the sexual process in cross-fertilizing hermaphrodites I→II, and then the \"red\" transition to gonochorism — II→IIIa.\n\nWhy do most authors consider precisely the transition I→IIIa? Tradition. The habit of considering gonochorism as the typical case of sexual reproduction. The inertia of thought, predetermined by Williams' balance argument. Finally, explaining how gonochorism defeats hermaphroditism is also a difficult task...\n\nI am quite confident that the sexual process, as well as the haploid-diploid cycle with fertilization and meiosis, arose in cross-fertilizing hermaphrodites. It was they who transitioned from non-homologous recombination during horizontal transfer to ordered homologous recombination during meiosis. It was they who assembled the mechanism of sexual reproduction from individual previously existing molecular blocks. And since they did not have to pay the double cost of sex, the drop in their reproductive efficiency did not block this fateful process.\n\nRemember, in the previous column I compared the reproductive efficiency of clonal and gonochorous organisms? In the case of transition I→IIIa, if the advantage of gonochorous organisms was less than twofold (!), clonal organisms won. Whereas in transition I→II, only a small advantage of hermaphrodites is sufficient. Look.\n\n\"Compe-\ntition\" 2\n\nType of repro-duction\n\nProducers\n\nOff-spring\n\nSur-vival\n\nWill survive\n\nTotal\n\nResult\n\nVariant A\n\nClonality\n(type I)\n\nIndividual 1\n\n100\n\n25%\n\n25\n\n50\n\nLoss\n\nIndividual 2\n\n100\n\n25\n\nVariant D\n\nCross hermaph-roditism\n(type II)\n\nPartner 1\n\n100\n\n26%\n\n26\n\n52\n\nVictory\n\nPartner 2\n\n100\n\n26\n\nThe reasons for the transition from clonal reproduction to cross-fermaphroditism can be imagined quite well. The development of new adaptations was accelerated thanks to the \"vicar of Bray\" and Bell's shore, and the action of the Red Queen made this process endless. Getting rid of deleterious mutations was facilitated thanks to escaping from the power of Muller's ratchet and the action of Kondrashov's axe. The spread of hermaphrodites was promoted by Williams' lottery. And there was no need to pay Maynard Smith's \"double cost of sex.\"\n\nOne question remains. But how could hermaphrodites transition to the separation of male and female roles? After all, this required payment of the \"double cost of gonochorism\" — a twofold decrease in reproductive efficiency.\n\nI have an answer to this question. I will give it in the next column.\n\n←\nDmytro Shabanov\n→\n\nWill eight indirect hypotheses describing the advantages of sexual reproduction suffice to explain the origin of sex?\nIs the Red (a.k.a. Black) Queen truly the queen of evolutionary problems? Don't rush to answer — don't forget about hermaphrodites!\nThe victory of stability over optimality, or Why hermaphrodites lose to males and females\n\nColumn for Kompyuterra #133\nColumn for Kompyuterra #134\nColumn for Kompyuterra #135"}