Походження статі, роздвоєності статей та геміклонального успадкування. Формулювання задачі. Колонка для Комп'ютерри #130
Поява статевого розмноження. Поява статевого розділення. Екологічна диференціація статей. Відмова від статевого процесу та перехід до клонального відтворення. Поява геміклональності. Можливий відмова від геміклональності. Завдання цієї колонки полягало...
Україна — велика Врадиевка. Вибрані місця з листування з російськими та проросійськими друзями
Походження статі, роздвоєності та геміклонального успадкування. Постановка задачі
Загадка сексу. Cui prodest: ген, індивід, група?
Колонка для Комп’ютерри #129
Колонка для Комп’ютерри #130
Колонка для Комп’ютерри #131
Лише-ліше почавши співпрацювати з мережею «Комп’ютерра», я написав колонку, де підвів розмову до теми походження роздвоєності. Ця колонка була четвертою за рахунком. Там я обґрунтовував, чому роздвоєність, широко поширену серед високоорганізованих живих організмів, пояснити не так просто. У сьомій колонці я пояснив, чому широко відомі (і дуже цікаві) ідеї Віґена Геодакяна не дозволяють пояснити походження (саме походження, а не успіх) роздвоєності. У тих статтях я пообіцяв найближчим часом продовжити розмову на цю тему… і не виконав свого обіцянки. Тоді мені здавалося, що залишилося лише трохи доопрацювати майже готову модель. На жаль, за минулі два з половиною роки я так і не зміг впоратися з цією задачею. Але до теми повернутися все ж треба, і я це зроблю — починаючи з цієї, сто тридцятої за рахунком колонки. Тепер я спробую опрацювати тему більш ґрунтовно.
Перший крок, який треба зробити, — задати класифікацію типів популяційного відтворення. Зверніть увагу: мова йде не про типи розмноження (їх класифікації розроблені давно). Розмноження — це процес, який відбувається на рівні організмів, а на рівні популяцій відбувається відтворення. Однорідне популяційне відтворення може здійснюватися завдяки принципово різним способам розмноження.
Поясню цю думку на прикладах.
— Амеба протей ділиться навпіл у результаті базового типу ділення у всіх ядерних організмів, мітозу. Спочатку ділиться ядро (з утворенням двох ідентичних дочірніх ядер), потім — цитоплазма клітини.
— Малий морський багатощетинковий черв’як Ctenodrilus monostylos втратив здатність до статевого процесу і розмножується за допомогою архітомії. При цій формі вегетативного розмноження тіло розділяється на передню і задню половини, кожна з яких доращує недостаючу частину.
— Скельові ящірки роду Darevskia (названого так на честь описавшого їх розмноження І. С. Даревського) розмножуються шляхом партеногенезу (застаріла назва — «девственне розмноження»). Партеногенез, за прийнятою класифікацією, — одна з форм статевого розмноження; перед утворенням яйцеклітини хромосоми в клітині‑попередниці подвоюються шляхом ендомітозу (подвоєння хромосом без ділення клітини), і утворена яйцеклітина може розвиватися без запліднення.
— У наїзників (перепончатокрилих комах), заражених паразитичною бактерією Wolbachia, бактерії зупиняють перше ділення мейозу (клітинного ділення, в ході якого утворюються статеві клітини), приводячи до паразитарно індукованого партеногенезу.
— Сріблясті карасі у багатьох водоймах розмножуються шляхом гіногенезу, при якому ікринка, відкладена самкою карася, активується сперматозоідом будь‑якої карпової риби. Ядро сперматозоїда в ікринку не потрапляє, і розвиток потомства забезпечується ядром яйцеклітини.
Ілюстрації до згаданих прикладів клонального відтворення: амеба Amoeba proteus; архітомія поліхети Ctenodrilus monostylos; скельова ящірка Darevskia unisexualis; наїзник Trichogramma і його яйце, заражене бактеріями Wolbachia (світлі точки); сріблястий карась Carassius gibelio
Описані форми розмноження відрізняються одна від одної практично фундаментально. А з точки зору популяційного відтворення вони дуже схожі, оскільки породжують клони — сукупності генетично ідентичних особин. У всіх цих випадках дочірні особини генетично не відрізняються від материнських (строго кажучи, це твердження не абсолютно вірне, але для подальшого викладу це не суттєво). Популяції, що складаються з клонів, принципово відрізняються від популяцій зі статевим відтворенням. Фактично це «пучки» з незалежних клональних ліній, здатних розгалужуватись і вмирати. До речі, функціонування клональних популяцій, механізми, завдяки яким підтримується їх приналежність до одного виду, і взагалі розуміння того, чим є вид у клональних організмів, розроблені сучасною наукою цілком недостатньо…
Отже, одні й ті ж типи відтворення можуть розвиватися на різній основі. Щоб їх порівнювати, їх треба виділити і позначити. Класифікація типів популяцій за механізмами їх відтворення, яку я пропоную, показана на схемі. Ця класифікація є результатом деталізації тієї, що я публікував кілька років тому. Однак перед тим, як перейти до її обговорення, зроблю важливе уточнення.
Класифікація, яку ми будемо обговорювати, стосується лише популяцій з постійним способом розмноження особин. Окрім них, відомо безліч випадків складних життєвих циклів або чергування способів розмноження. У багатьох видів статеве розмноження чергується з безстатевим або розмноження з заплідненням чергується з партеногенезом. Так, дафнії і тлі протягом літа утворюють клональні лінії партеногенетичних самок, а перед відходом на зиму у них з’являється роздвоєне покоління. Самці і самки виробляють запліднені яйця, які чекають наступної весни.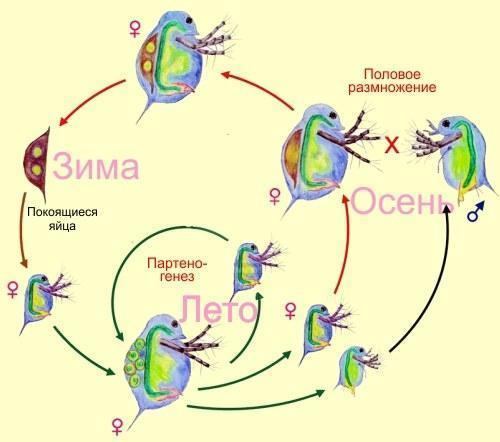
Циклічний партеногенез у дафній
Строго кажучи, такі способи відтворення не вписуються в запропоновану класифікацію. Однак завжди можна сказати, наприклад, що дафнії і тлі поєднують підтипи відтворення IIIa і IIb. Проте чергування поколінь — у будь‑якому випадку окрема тема. Відклали його до кращих часів, перейдемо до розгляду типів популяцій з постійним характером відтворення.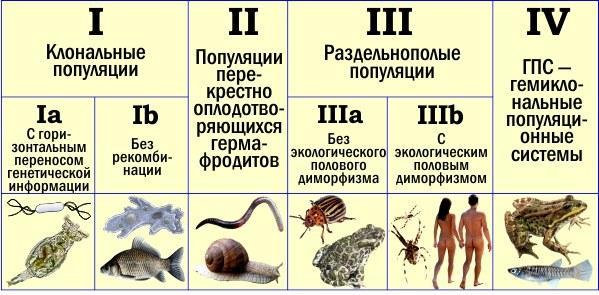
Класифікація типів популяційного відтворення (у видів з постійним характером розмноження особин). Детальні пояснення — у тексті
Тип I представлений вже згаданими клональними популяціями. Він ділиться на два підтипи залежно від того, чи ізольовані клональні лінії одна від одної генетично.
Для підтипу Ia характерна здатність до горизонтальної передачі генетичної інформації («вертикальною» вважається передача всередині генеалогічної послідовності, від предків до нащадків, а «горизонтальною» — між неродичними організмами). Найбільш різноманітні бактерії належать саме до цього типу. Горизонтальний перенос генів у прокаріот (доядерних організмів) був відкритий на прикладі передачі стійкості до антибіотиків. Останнім часом стало ясно, що такий перенос — широко поширене явище. Для бактерій нехарактерні ізольовані видові генофонди. «Куски» генетичного матеріалу передаються від «виду» до «виду» за допомогою різних механізмів.
Саме тому в стовпці Ia намальована якась бактерія. Під нею знаходиться зовсім інша істота — бделлоїдна коловратка. Це мікроскопічний черв’як — отже, еукаріотичний (ядерний) організм. Як у більшості інших тварин, і, зокрема, у нас з вами, у його клітинах знаходиться подвійний набір генетичної інформації, розташований на подвійній (диплоїдній) хромосомній системі (що містить ДНК клітинних структур). Особливість у тому, що у нас з вами один набір отриманий від батька, а інший — від матері, а бделлоїдні коловратки втратили статеве розмноження у досить далекому еволюційному минулому. Наші парні хромосоми містять схожі (з точністю до індивідуальних відмінностей) версії одного і того ж генетичного тексту. Парні хромосоми бделлоїдних коловраток перестали, по суті, бути парними: вони містять функціонально різний текст, який буквально набитий фрагментами, отриманими від інших організмів у результаті горизонтального переносу. Розмножуються ці дивовижні черв’яки шляхом партеногенезу.
Для бактерій, ймовірно, можлива ситуація, коли чужий фрагмент генетичної інформації вбудовується куди попало і виявляється корисним. Така вставка не є повністю випадковим текстом. Це результат відбору, що проходив у інших організмів, і він, можливо, пов’язаний з якимись цінними властивостями. Еукаріоти перестраюють свою генетичну інформацію інакше. Пам’ятаєте, у недавніх колонках про епігенетичну теорію еволюції я говорив, що розвиток високорозвинених організмів регулюється набагато складніше, ніж у бактерій? Якщо в ході індивідуального розвитку відбувається багатоступеневе взаємодія між генами і продуктами їх роботи, якщо активність генів регулюється складними генними мережами, то шанс, що випадковий кусок генетичного тексту, вбудований у випадкове місце, виявиться потенційно корисним, дуже малий.
Горизонтальний перенос зустрічається і у еукаріот, але в цілому для них характерний зовсім інший механізм рекомбінації, тісно пов’язаний зі статевим розмноженням. При заплідненні утворюється клітина з подвоєним набором хромосом. Значить, у процесі утворення статевих клітин набір хромосом має скоротитися вдвічі. Це відбувається під час ділення, яке називається мейозом. У мейозі парні хромосоми розпізнають один одного і обмінюються фрагментами в процесі, названому кросинговером (або просто перекриттям). Нова генетична інформація, яка несе сліди дії відбору, що відбувався у інших організмів, потрапляє при цьому не куди попало, а саме в те місце хромосоми, де вже існують механізми регуляції її активності.
Проте зараз ми говоримо про клональні популяції, для яких статевий процес і пов’язана з ним рекомбінація нехарактерні. Ми виділяємо підтип IIa, який характеризується відмовою від систематичного використання горизонтального переносу. Представники такого підтипу регулярно виникають від предків, що належали до наступних типів, але, ймовірно, можуть також походити і від популяцій Ia. На схемі показані амеба протей і сріблястий карась, розмноження яких ми вже обговорювали.
До типу II належать популяції крос‑оплоджуваних гермафродитів. На схемі зображені дощовий черв’як і виноградна равлик: це ті тварини, що при розмноженні утворюють пари з гермафродитних особин, кожна з яких передає своєму партнеру чоловічі статеві продукти. Такі істоти вже повністю використовують ключові переваги статевого розмноження і пов’язаної з ним рекомбінації (наслідки кросинговеру під час утворення статевих клітин), але ще не несуть навантаження роздвоєності.
Тип III. Роздвоєні. Поділений на два підтипи залежно від того, чи є значущі екологічні відмінності між самками і самцями. Можливо, розділяти цей тип на підтипи не варто було… Головна причина, чому це все ж зроблено, така. Є серйозні причини, у силу яких екологічний статевий диморфізм виявляється вигідним, і тут я цілком згоден з Віґеном Геодакяном. Однак очевидно, що роздвоєність має спочатку просто виникнути (підтип IIIa) — і лише потім між двома статями можуть накопичитися серйозні відмінності, що не стосуються безпосередньо сфери розмноження (підтип IIIb).
Підтип IIIa на схемі проілюстрований зображеннями колорадського жука і зеленої жаби. До речі, певний статевий диморфізм у зелених жаб є: самки у них (як і у багатьох інших жаб) більші за самців. Проте спосіб життя обох статей жаб приблизно однаковий; вони розміщені в IIIa саме для того, щоб показати, що певні відмінності між статями можуть спостерігатися і в цьому підтипі. До підтипу IIIb на схемі віднесені павуки‑хрестовики і люди.
Нарешті, тип IV. Геміклональні (напівклони) популяційні системи, ГПС. Зазвичай їх не розглядають, як щось окреме від усіх інших типів популяцій (саме поняття ГПС ми з колегами придумали і ввели лише кілька років тому). На схемі — їстівна жаба (Pelophylax esculentus) і американська карпозуба риба роду Poeciliopsis, у якої вперше був описаний тип розмноження, характерний і для жаб. У таких організмів міжвидові гибриди передають у статеві клітини геном одного з батьківських видів як єдине ціле (майже як єдине ціле). Міжвидові гибриди живуть спільно з представниками батьківських видів, схрещуються з ними і спільно беруть участь у відтворенні. З покоління в покоління в таких системах передаються як клональні (практично незмінні), так і рекомбінантні, звичайні геноми.
Вичерпано чи перелік типів відтворення? Не знаю. З урахуванням тих ступенів свободи, за якими різняться перелічені типи, якихось інших їх поєднань я не знаю, але це не означає, що їх не може бути...
У розміщеній нижче таблиці показані результати порівняння різних типів популяцій за тими ознаками, які здаються мені суттєвими. Зверніть увагу, що тип IV виявляється цілком самостійним. Найменші відмінності цілком закономірно спостерігаються між підтипами IIIa і IIIb. На жаль, у цій колонці я встигну обговорити не всі графи таблиці: наприклад, розмова про рекомбінацію вимагатиме окремої колонки. Все ж сподіваюся, що напрямок для обговорення я задала.
Тип популяції
Ia
Ib
II
IIIa
IIIb
IV
Генеалогія
Лінійна
Сітчаста
Лінійна
«Подвійна ціна» статі
Ні (нащадок безпосередньо виробляють усі особи)
Так (нащадок безпосередньо виробляють лише самки)
Статевий відбір
Ні
Малоефективний
Ефективний
Є
Унікальність особини
Ні
Успадкована
Неуспадкована
Генофонд
Сукупність клонів
Рекомбінантний генофонд популяції
Рекомбінантний генофонд + напівклони
Ізоляція
Потік генів між клонами
Ізольовані клони
Потік генів всередині виду, ізольованого від інших видів
Потік генів всередині і між видами
Рекомбінація
Хаотична
Ні
Упорядкована внутрі‑видова
Упорядкована між‑ і внутрі‑видова
А тепер я зроблю те, заради чого, по суті, і писав цю колонку. Я повторю ту схему, що ви вже бачили, додавши в неї умовні позначення переходів між типами, які відбувалися в ході еволюції. Безперервні стрілки позначають ті еволюційні переходи, щодо яких ми знаємо, що вони відбувалися, або можемо це припускати з високою впевненістю (наприклад, перехід, показаний зеленим кольором, відбувався або так: I→II, або так: I→IIIa).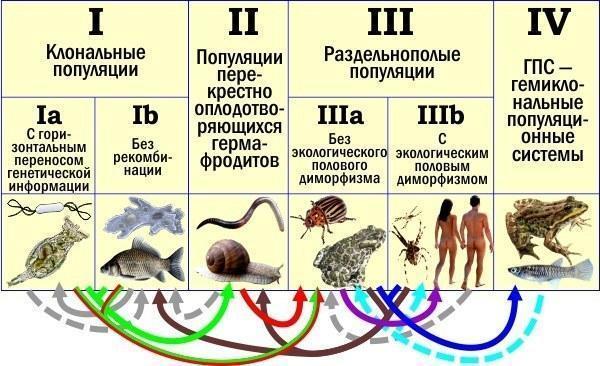
Я планую найближчим часом обговорити логіку наступних переходів.
Поява статевого розмноження: I→II (ймовірніше) або I→IIIa (зелені стрілки).
Поява роздвоєності: II→IIIa (ймовірніше) або I→IIIa (червоні стрілки).
Екологічна диференціація статей: IIIa→IIIb (фіолетова стрілка).
Відмова від статевого процесу і перехід до клонального відтворення: III→I або II→I (коричнева стрілка).
Поява геміклональності: III→IV (а що, чорт не жартує? Можливо, і II→IV) — синя стрілка.
Можливий відмова від геміклональності: IV→III (IV→II) — блакитна пунктирна стрілка.
Завдання цієї колонки полягало в тому, щоб задати схему, що дозволяє організувати обговорення щойно перелічених проблем. Я сподіваюся, що просто зможу повторити малюнок зі стрілками і за допомогою посилання надіслати пояснення всім, кому це потрібно, сюди, у цю колонку.
Згодні? Повірте, тут є про що розповісти і про що задуматися.