Borkin et al. (2008) On the Southern Boundary of the Distribution of Hybrid Rana esculenta...
A zoogeographic "common grave" from which the most interesting directions for further searches for undescribed populations of pool frogs and edible frogs emerge. Its PDF copy is available here. Borkin L. Ya., Bezman-Moseiko O. S., Mazepa G. A., Zinenko A. I., Korshunov A. V., Lada G. A., Shabanov D....
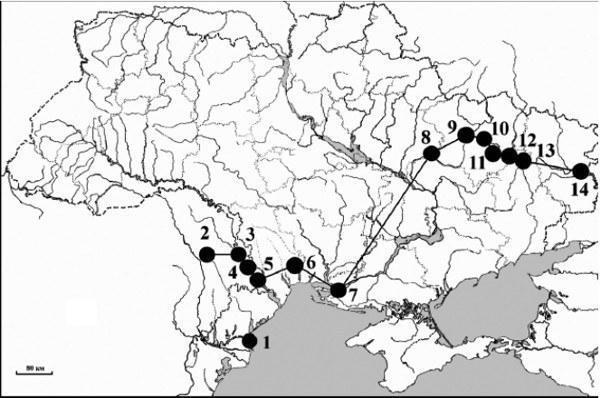
Borkin L. Ya., Bezman-Moseiko O. S., Mazepa G. A., Zinenko A. I., Korshunov A. V., Lada G. A., Shabanov D. A., Litvinchuk S. N., Rozanov Yu. M. On the southern boundary of the distribution of hybrid Rana esculenta (Ranidae, Anura, Amphibia) in Ukraine and Moldova: DNA flow cytometry data // Proceedings of the Ukrainian Herpetological Society. – No. 1, 2008. – Pp. 3–8. UDC 595.6, 597.94 ON THE SOUTHERN LIMIT OF THE RANGE OF HYBRID RANA ESCULENTA (RANIDAE, ANURA, AMPHIBIA) IN UKRAINE AND MOLDOVA: DNA FLOW CYTOMETRY EVIDENCE L. Ya. Borkin1, O. S. Bezman-Moseiko1, G. A. Mazepa2, A. I. Zinenko3, A. V. Korshunov2, G. A. Lada4, D. A. Shabanov2, S. N. Litvinchuk5, Yu. M. Rozanov5 1 Zoological Institute of the Russian Academy of Sciences, Universitetskaya Embankment, 1, St. Petersburg, 199034 Russia; E-mail: lacerta@zin.ru 2 V. N. Karazin Kharkiv National University, Svobody Square, 4, Kharkiv, 61077 Ukraine; E-mail: d_sh@i.ua 3 Museum of Nature of V. N. Karazin Kharkiv National University, Trinklera St., 8, Kharkiv, 61022 Ukraine; E-mail: zinenkoa@yahoo.com 4 Tambov State University, Internatsionalnaya St. 33, Tambov, 392000 Russia; E-mail: esculenta@mail.ru 5 Institute of Cytology of the Russian Academy of Sciences, Tikhoretsky Ave., 4, St. Petersburg, 194064 Russia; E-mail: slitvinchuk@yahoo.com On the Southern Limit of Range of Hybrid Rana esculenta (Ranidae, Anura, Amphibia) in Ukraine and Moldova: DNA Flow Cytometry Evidence. Borkin L. J., Bezman-Moseiko O. S., Mazepa G. A., Zinenko A. I., Korshunov A. V., Lada G. A., Shabanov D. A., Litvinchuk S. N., Rozanov J. M. — Using DNA flow cytometry, the distribution of the hybridogenetic species Rana esculenta in Moldova and Ukraine was established. According to the data obtained, the southern boundary of the distribution of R. esculenta can be traced from west to east through the following localities: Vilkovo (Danube River delta), Vulcanesti (Moldova), Doibany, Tiraspol, and Kitscany (Dniester River), Berezovo (Tiligul River), Golaya Pristan District (Dnieper River), Russky Orchik (Kharkiv Province, Ukraine), and along the middle reach of the Seversky Donets River from the north-eastern part of Kharkiv Province (Ukraine) to Rostov Province (Russia). In the overwhelming majority of cases, R. esculenta was represented only by diploid individuals, and the population systems were of RE type. Only in Russky Orchik was the REL system reliably detected. Records of R. esculenta in Kherson Province are geographically isolated from populations inhabiting the northern and central parts of Ukraine. However, this large distributional gap appears to be associated more with insufficient sampling of the area than with the relict nature of these southernmost populations. Key words: Rana esculenta, genome size, Ukraine, Moldova On the Southern Limit of Range of Hybrid Rana esculenta (Ranidae, Anura, Amphibia) in Ukraine and Moldova: DNA Flow Cytometry Evidence. Borkin L. J., Bezman-Moseiko O. S., Mazepa G. A., Zinenko A. I., Korshunov A. V., Lada G. A., Shabanov D. A., Litvinchuk S. N., Rozanov J. M.— Based on DNA flow cytometry, we identified green frog localities with hybrid Rana esculenta. We found or confirmed some southernmost localities of the species situated in Moldavia and Ukraine. According to our data, the southern distributional border of R. esculenta is demarcated as follows from the west to the east: Vilkovo (Danube River Delta), Vulcanesti (Moldova), Doibani, Tiraspol', and Kitscani (Dniestr River), Beryozovo (Tiligul River), Golaya Pristan District (Dnieper River), Russky Orchik (Kharkov Province, Ukraine) as well as along the middle part of Seversky Donets River in north-eastern Ukraine and Rostov Province in adjacent Russia. The majority of samples contained diploid hybrid frogs from the RE kind of population systems. The REL system was identified in Russky Orchik. The records of R. esculenta in Kherson Province of Ukraine are geographically far isolated from populations in northern and central parts of Ukraine. However, this large gap seems to be resulted from inadequate frog sampling and species identification rather than from relic nature of the southernmost records. Introduction In recent decades, the hybridogenetic Rana esculenta complex has attracted close attention from researchers in many countries. It turned out to possess unusual evolutionary-genetic mechanisms whose study is of great importance for understanding speciation processes, not only among amphibians. These include, in particular, hybridization, non-Mendelian (meroclonal) inheritance, polyploidy, and diverse types of population systems incorporating both unisexual (either males or females) and bisexual hybrids. The distribution of the hybridogenetic complex across the territory of the European part of the former USSR has been insufficiently studied. This is largely due to the unreliability of species identification using traditional diagnostic characters, since the ranges of variation in body proportions and coloration of the hybrid edible frog (Rana esculenta Linnaeus, 1758) and the parental species (Rana lessonae Camerano, 1882 and Rana ridibunda Pallas, 1771) overlap considerably, especially in triploid individuals (Planer et al., 1994; Lada et al., 1995; Schruer, 1996; Borisovsky et al., 2001; Ruchin et al., 2005). The mass occurrence of polyploids (3n and 4n) was recently discovered in north-eastern Ukraine and Rostov Province of Russia (Borkin et al., 2004, 2006; Borkin et al., 2005). In light of the above, questions arise regarding the reliability of published data obtained using external morphological characters. At present, adequate identification of species within the hybridogenetic complex is possible using biochemical methods such as protein electrophoresis (see Mezhzherin et al., 2007) and DNA cytometry (see Borkin et al., 1987; Borkin et al., 2004). Over recent decades, we have examined large numbers of green frogs from many regions of the European part of the former USSR. The overwhelming majority of samples were studied by DNA flow cytometry, which allows not only accurate species identification of each individual but also determination of ploidy level and, for males, gamete type. In cases of polyploidy, the genomic composition of each individual can also be determined. Taking these data into account, we have been able to substantially refine the distribution of R. esculenta. In the present paper we report materials characterizing the southern boundary of the range of this hybrid species in Ukraine and Moldova. Materials and Methods Species identification and ploidy level of frogs were determined by genome size as measured by DNA flow cytometry (Borkin et al., 2004). After anesthesia, erythrocytes were taken directly from the heart. Test cells were mixed with reference cells and analyzed together, so that both the cells under study and the reference cells were stained and measured under identical conditions. Erythrocytes of grass frogs (R. temporaria) collected in Leningrad and Pskov Provinces of Russia served as the reference standard. Details of this method have been described previously (Vinogradov et al., 1990; Rozanov, Vinogradov, 1998; Borkin et al., 2001). Results and Discussion Among the 1,497 green frogs examined from 131 localities in Moldova and Ukraine, 774 individuals from 76 sites proved to be hybrids. The southernmost localities were (Fig. 1): the town of Vilkovo (Kiliya District, Odessa Province, Ukraine), the village of Vulcanesti (Nisporeni District, Moldova), the village of Doibany 2nd (Dubossary District, Transnistria, Moldova), the city of Tiraspol (Tiraspol District, Transnistria, Moldova), the village of Kitscany (Slobodzeia District, Transnistria, Moldova), Berezovsky Forest (Berezovsky District, Odessa Province, Ukraine), the villages of Podtemnoye and Rybalche (Golaya Pristan District, Kherson Province, Ukraine), the locality of Russky Orchik (Zachepilivka District, Kharkiv Province, Ukraine), the village of Velikaya Gomolsha (Zmiyiv District, Kharkiv Province, Ukraine), the village of Kreydyanka (Balakliya District, Kharkiv Province, Ukraine), the settlement of Chervony Shakhtyor and the village of Yaremovka (Izium District, Kharkiv Province, Ukraine), the city of Svyatogorsk (Svyatogorsk District, Donetsk Province, Ukraine), the village of Stanichno-Luganskoe (Stanichno-Lugansk District, Luhansk Province, Ukraine). [IMG_1] Fig. 1. The southern boundary of the distribution of Rana esculenta in Ukraine and Moldova. 1 — Vilkovo; 2 — Vulcanesti; 3 — Doibany 2nd; 4 — Tiraspol; 5 — Kitscany; 6 — Berezovsky Forest; 7 — Golaya Pristan District; 8 — Russky Orchik; 9 — Velikaya Gomolsha; 10 — Kreydyanka; 11 — Chervony Shakhtyor; 12 — Yaremovka; 13 — Svyatogorsk; 14 — Stanichno-Luganskoe. In the PhD dissertation of N. M. Suryadnaya (2005, p. 277 and Fig. 7.9), a list of records of R. esculenta in Ukraine is presented. In addition to the main cluster of localities in the northern and central parts of the country, this author also indicated two districts that are geographically isolated from each other and from the main body of known sites for the species: the Danube River delta (Vilkovo) and the lower reaches of the Dnieper River (the villages of Staraya Zburyevka, Zabarino, and Gerojskoe in Golaya Pristan District of Kherson Province). A similar distribution pattern was shown for the pool frog R. lessonae (except for the locality Gerojskoe). Identification of both species was carried out by external morphology and partly by chromosome morphometry. Our DNA flow cytometry data confirm the occurrence of R. esculenta in these areas. It should also be noted that hybrids from the Danube River delta had been detected earlier using protein electrophoresis (Gather et al., 1991; Mezhzherin et al., 2007). For the territory of Moldova, several authors (see Borkin et al., 1997) recorded two species of green frogs: R. ridibunda and R. "esculenta" (= R. lessonae). Interestingly, in the PhD dissertation of V. E. Tofan (1966) the latter species was listed for the village of Suklei in Tiraspol District (Transnistria), the village of Karbuna in Novo-Anensky District, the villages of Bykovets and Chuteshty in Kalarash District, and the village of Filipeny in Leovo District (Moldova). However, diploid hybrids had previously been found by us only near the village of Vulcanesti in Nisporeni District (Borkin et al., 1997). Rana lessonae was not found there. In 2006, in a sample of 36 individuals caught in the city of Tiraspol, two diploid hybrids were identified; all other individuals proved to be R. ridibunda. A targeted survey conducted by us in 2007 revealed a fairly wide distribution of R. esculenta in the Slobodzeia, Tiraspol, and Dubossary Districts of Transnistria; however, R. lessonae could not be found. All hybrid specimens (n = 31) were also diploids. Thus, these data allow us to trace the southern boundary of the distribution of R. esculenta along the lower reaches of the Dniester River. In 2007, on the Tiligul River in Berezovsky Forest (Berezovsky District, Odessa Province, Ukraine), 10 individuals were caught, 9 of which proved to be diploid hybrids and one was R. ridibunda. A noteworthy feature of this record is that it does not belong to the catchments of the Dniester or Dnieper rivers and occupies a geographically intermediate position between them. It should be noted that reliable records of numerous hybrids in Transnistria and the Tiligul River basin geographically connect R. esculenta from such distant areas as the deltas of the Danube and Dnieper rivers. In 2007, we were also able to confirm the occurrence of R. esculenta (3 diploids) at two localities (Podtemnoye and Rybalche) in Golaya Pristan District of Kherson Province. Furthermore, based on acoustic and external morphological characters, this species was identified in the vicinity of the villages of Kokhany (Golaya Pristan District) and Peschanivka (Tsyurupynsk District). At the latter locality, some specimens of green frogs were identified by us as R. lessonae. Earlier, in other parts of the same province (Kherson, Golaya Pristan, and Tsyurupynsk), only R. ridibunda had been recorded by us. The following cytometric data concern north-eastern Ukraine. This region, as is now well established, is characterized by a wide distribution of R. esculenta. Its unique feature (within the European part of the former USSR) is the mass occurrence of polyploidy among hybrids found along the middle reach of the Seversky Donets River over a stretch of 480 km (Borkin et al., 2004, 2006; Borkin et al., 2005). Of several new localities surveyed by us, two deserve special attention in the context of this paper. At the Russky Orchik locality (Kapinernoye Lake, the villages of Zarechye and Damansky), all three species of the R. esculenta complex were identified (all hybrids diploid), which confirms earlier data of V. I. Vedmederya (1984) and G. A. Lada (Borkin et al., 2004) based on external characters. At another locality (village of Yaremovka), 4 individuals were caught and classified by genome size as triploid hybrids of RRL (n = 3) and RLL (n = 1) types. Other new records, to be reported in a separate paper, did not extend beyond the previously known geographic boundaries. The most south-eastern record of R. esculenta was made by us in Russia, in Rostov Province, which borders the eastern part of Ukraine (Luhansk Province). The nearest previously known record of this species in Russia had been 320 km to the north (Borkin et al., 2006). For completeness, it is important to mention several additional records of green frogs in Ukraine that were identified by external characters as R. esculenta or R. lessonae: the village of Krymka (Pervomaysk District, Mykolaiv Province; R. esculenta — Kuzmin, 1999), the village of Alexandrovka (Voznesensk District, Mykolaiv Province; R. lessonae — our data), and the Black Forest near the village of Znamianka (Znamianka District, Kirovograd Province; R. esculenta, R. lessonae, and R. ridibunda — our data). Thus, summarizing the above, it can be concluded that the southern boundary of the range of the edible frog (and apparently also of the pool frog) runs parallel to the Forest-Steppe/Steppe boundary, shifting northward with increasing continentality of climate and penetrating far to the south along major rivers such as the Danube, Dniester, and Dnieper. In the context of population systems analysis, it should be noted that the overwhelming majority of samples located at the southern boundary of the range of R. esculenta belong to the RE type. The exceptions are populations from the Russky Orchik locality and, probably, Vilkovo (REL type). In all the southernmost habitats, hybrids were generally represented only by males; the exception is north-eastern Ukraine, where R. esculenta populations may include both sexes or only females. On the basis of available information (Suryadnaya, 2005, Fig. 7.9; Pisanets, 2007, Fig. 100; our data), there is a "gap" in the distribution of R. esculenta in Ukraine between the northern and central regions on one hand, and the extreme south on the other. However, in our view, this "cartographic disjunction" is due primarily to the absence of adequate sampling and correct identification rather than to the relict nature of the southern populations alone. This "blank spot" may therefore disappear in the future with more thorough study of the distribution of green frogs. The authors express their gratitude to V. G. Dyadichko (Odessa Branch of the Kovalevsky Institute of Biology of the Southern Seas, NAS of Ukraine) for assistance in collecting material. On the Russian side, the work was supported by grants NSh-4212.2006.4, RFBR 05—04–48403, and the Centre for Collective Use "Materials Science and Diagnostics in Advanced Technologies"; on the Ukrainian side, by grant INTAS Ref. Nr 05-1000008-8147. Borisovsky A. G., Borkin L. Ya., Litvinchuk S. R., Rozanov Yu. M. Morphometric characterization of green frogs (Rana esculenta complex) of Udmurtia // Bulletin of Udmurt University. — 2000. — No. 5. — Pp. 70–75. Borkin L. Ya., Vinogradov A. E., Rozanov Yu. M., Tsaune I. A. Semiclonal inheritance in the hybridogenetic Rana esculenta complex: evidence from DNA flow cytometry // Doklady of the Academy of Sciences of the USSR. — 1987. — 295, No. 5. — Pp. 1261–1264. Borkin L. Ya., Zinenko A. I., Korshunov A. V. et al. Mass polyploidy in the hybridogenetic Rana esculenta complex (Ranidae, Anura, Amphibia) in eastern Ukraine // Proceedings of the First Conference of the Ukrainian Herpetological Society (October 10–12, 2005, Kyiv). — Kyiv: Zoological Museum of the National Museum of Natural History, NAS of Ukraine, 2005. — Pp. 23–26. Vedmederya V. I. Some data on frogs of the genus Rana in Kharkiv Province (based on materials of the Museum of Nature, KhSU) // Bulletin of Kharkiv University. — 1984. — No. 262. — Pp. 99–101. Kuzmin S. L. Amphibians of the Former USSR. — Moscow: KMK Scientific Press, 1999. — 298 pp. Mezhzherin S. V., Morozov-Leonov S. Yu., Nekrasova O. D., Kurtyak F. F. et al. Evolutionary-genetic aspects of semiclonal reproduction of the hybrid form Rana kl. esculenta (Amphibia, Ranidae) // Scientific Bulletin of Uzhhorod University. Biology Series. — 2007. — Issue 1. — Pp. 79–84. Pisanets E. M. Amphibians of Ukraine. A field guide to the amphibians of Ukraine and adjacent territories. — Kyiv: Zoological Museum of the National Museum of Natural History, NAS of Ukraine, 2007. — 312 pp. Rozanov Yu. M., Vinogradov A. E. Precision DNA cytometry: study of individual variability in animal genome size // Tsitologiya [Cytology]. — 1998. — 40, No. 8/9. — Pp. 792–799. Ruchin A. B., Borkin L. Ya., Lada G. A. et al. Morphological variability, genome size, and population systems of green frogs (Rana esculenta complex) in Mordovia // Bulletin of the Moscow Society of Naturalists. Biological Series. — 2005. — 110, Issue 2. — Pp. 3–10. Suryadnaya N. N. Green frogs of the fauna of Ukraine: morphological variability, karyology, and biology: PhD dissertation. — Kyiv, 2005. — 27 pp. Tofan V. E. Fauna of amphibians and reptiles of Moldova: PhD dissertation. — Leningrad, 1966. — 350 pp. Borkin L. J., Korshunov A. V., Lada G. A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in eastern Ukraine // Russ. J. Herpetol. — 2004. — 11, N 3. — P. 203–222. Borkin L. J., Lada G. A., Litvinchuk S. N. et al. The first record of mass polyploidy in hybridogenetic green frog Rana esculenta in Russia (Rostov Oblast') // Russ. J. Herpetol. — 2006. — 13, N 1. — P. 77–82. Borkin L. J., Litvinchuk S. N., Rosanov Y. M. Amphibians and reptiles of Moldavia: additions and corrections, with a list of species // Russ. J. Herpetol. — 1997. — 4, N 1. — P. 50–62. Borkin L. J., Litvinchuk S. N., Rosanov J. M., Milto K. D. Cryptic speciation in Pelobates fuscus (Anura, Pelobatidae): evidence from DNA flow cytometry // Amphibia—Reptilia. — 2001. — 22, N 4. — P. 387–396. Gbuther R., Plotner J., Tetzlaff I. Zu einigen Merkmalen der Wasserfrosche (Rana synkl. esculenta) des Donau-Deltas // Salamandra. — 1991. — 27, H. 4. — S. 246–265. Lada G. A., Borkin L. J., Vinogradov A. E. Distribution, population systems and reproductive behavior of green frogs (hybridogenetic Rana esculenta complex) in the Central Chernozem Territory of Russia // Russ. J. Herpetol. — 1995. — 2, N 1. — P. 46–57. Plotner J., Becker C., Plutner K. Morphometric and DNA investigations into European water frogs (Rana kl. esculenta Synklepton (Anura, Ranidae)) from different population systems // J. Zool. Syst. Evol. Research. — 1994. — 32. — P. 193–210. Schruer T. Morphologie und Ploidiegrade von Wasserfrdschen aus unterschiedlichen Populationssystemen in Nordost-Polen // Zeitschrift fur Feldherpetologie. — 1996. — 3. — S. 133–150. Vinogradov A. E., Borkin L. J., Gunther R., Rosanov J. M. Genome elimination in diploid and triploid Rana esculenta males: cytological evidence from DNA flow cytometry // Genome. — 1990. — 33, N 5. — P. 619–627