
Ecology: the biology of interaction. IV-13. Gause's competitive exclusion principle
In 1931–1935, the young Soviet biologist Georgii Franzovych Gause attempted to experimentally verify the competition equations derived by V. Volterra. Gause understood that for experiments in population biology, the simplest organisms are well suited: they require a very small amount of space...
IV-13. Gause’s competitive exclusion principle
The more similar two species are, the more their resource needs overlap and the stronger the competition between them. The sharpest competition is intraspecific. Yet individuals of the same species, although they compete intensely for resources, jointly form the next generation. What happens if competition between two different species reaches the same intensity as intraspecific competition?
In 1931–1935 the young Soviet biologist Georgy Frantsovych Gause attempted to experimentally test the competition equations derived by V. Volterra. Gause realized that for experiments in population biology the simplest organisms are most suitable: they require a very small amount of environment, and their generational turnover is much faster than that of multicellular animals. Gause’s experiments, which made his name famous, were performed on the ciliate Paramecium caudatum and its closest relatives (fig. IV-12.1).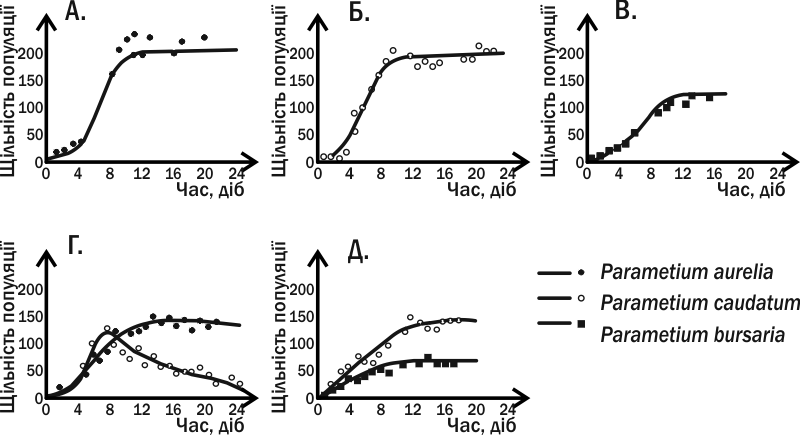
Fig. IV-13.1. Dynamics of the abundance of three ciliate species in G.F. Gause’s experiments. In experimental variants A., B. and C. the species were cultured singly; in cases G. and D. two ciliate species were introduced into the tube simultaneously.
Gause cultured three Paramecium species in tubes to which he added oat flour and periodically removed waste. Yeasts and bacteria grew on the oat flour and served as food for the ciliates. All three Paramecium species developed successfully in monoculture under these conditions. In mixed cultures P. aurelia always completely displaced P. caudatum. However, when P. aurelia and P. bursaria were cultured together the two species could coexist. Gause found that these two species differ in their feeding mode: the former fed on bacteria in the bulk liquid, the latter on yeast cells near the bottom.
Based on these observations the principle of competitive exclusion, or Gause’s rule, was formulated: two species occupying the same ecological niche cannot stably coexist in the same habitat; coexistence is possible only through niche partitioning.
The competitive exclusion rule is one of the possible explanations for the extraordinary species diversity in nature. If two populations in the same habitat cannot occupy the same niche, they must divide it. Over time each such population specializes in its own niche and may give rise to a new species.
Testing Gause’s rule in natural conditions is quite difficult, but some examples of its operation are known to biologists. Since the late 19th century, across much of Europe the broad‑clawed crayfish (Astacus astacus) has been displaced by the narrow‑clawed crayfish (A. leptodactylus). The niche of the narrow‑clawed crayfish overlaps that of the broad‑clawed, and the winning species is more fecund. The roach (Rutilus rutilus) displaces the common rudd (Scardinius erythrophthalmus) and the perch (Perca fluviatilis) in many water bodies. The adult niches are separated, while those of the juveniles overlap; the roach fry prove more competitive. However, the existence of many cases of competitive displacement does not prove that it always occurs. During the period in which biologists have studied Gause’s rule, both examples of its operation and situations where it fails have been found.
For example, in groups of planktonic algae niche partitioning is not observed. This is due to rapid environmental fluctuations, where transient conditions favor one species or the other, and also to intense predation pressure (zooplankton and various filter feeders) that prevents competitive exclusion from becoming the dominant factor in group formation. Yet in some cases competitive displacement does occur in planktonic assemblages. This happens during so‑called “water blooms,” when planktonic algae escape predation that normally limits their numbers. As a result of a bloom, only a few algal species become massive, each differing in its lifestyle, leading to competitive exclusion.
Some types of interactions can involve not only individuals from different populations but also relatives within a single population. For instance, intraspecific and interspecific competition are similar in that both limit the resources available to an individual. However, their effects on niche breadth are opposite. Intraspecific competition widens ecological niches: all individuals lack optimal resources for the species, and some are pushed to the periphery where only partially suitable resources exist. According to Gause’s rule, interspecific competition, on the contrary, narrows niches. Thus the observed niche widths in nature result from a balance of two opposing processes—expansion through intraspecific competition and contraction due to interspecific competition.
An interesting example of competitive exclusion was studied by Thomas Park with flour beetles. Two beetle species from the family Tenebrionidae (Tribolium confusum and T. castaneum) were kept in flour containers. These species competed for food (flour) and also could feed on each other, with beetles of both species primarily consuming individuals of the competing species rather than conspecifics (this combination of competition and predation is called antagonism). Park succeeded in finding conditions under which either the first or the second species consistently won. However, the most intriguing results were obtained under intermediate conditions that did not guarantee the advantage of either species (Table IV-13.1).
Table IV-13.1. Proportion of victories of the two flour beetle species in Thomas Park’s experiments
|
Table IV-13.1. Proportion of victories of the two flour beetle species in Thomas Park’s experiments |
“Climate” (experimental conditions) |
|
|
Tribolium confusum |
Tribolium castaneum |
|
|
Tribolium castaneum |
0 |
100 |
|
100 |
14 |
86 |
|
86 |
71 |
29 |
|
29 |
90 |
10 |
|
10 |
87 |
13 |
|
13 |
100 |
0 |
IV-13. Gause’s competitive exclusion principle
The more similar two species are, the more their resource needs overlap and the stronger the competition between them. The sharpest competition is intraspecific. Yet individuals of the same species, although they compete intensely for resources, jointly form the next generation. What happens if competition between two different species reaches the same intensity as intraspecific competition?
In 1931–1935 the young Soviet biologist Georgy Frantsovych Gause attempted to experimentally test the competition equations derived by V. Volterra. Gause realized that for experiments in population biology the simplest organisms are most suitable: they require a very small amount of environment, and their generational turnover is much faster than that of multicellular animals. Gause’s experiments, which made his name famous, were performed on the ciliate Paramecium caudatum and its closest relatives (fig. IV-12.1).
Fig. IV-13.1. Dynamics of the abundance of three ciliate species in G.F. Gause’s experiments. In experimental variants A., B. and C. the species were cultured singly; in cases G. and D. two ciliate species were introduced into the tube simultaneously.
Gause cultured three Paramecium species in tubes to which he added oat flour and periodically removed waste. Yeasts and bacteria grew on the oat flour and served as food for the ciliates. All three Paramecium species developed successfully in monoculture under these conditions. In mixed cultures P. aurelia always completely displaced P. caudatum. However, when P. aurelia and P. bursaria were cultured together the two species could coexist. Gause found that these two species differ in their feeding mode: the former fed on bacteria in the bulk liquid, the latter on yeast cells near the bottom.
Based on these observations the principle of competitive exclusion, or Gause’s rule, was formulated: two species occupying the same ecological niche cannot stably coexist in the same habitat; coexistence is possible only through niche partitioning.
The competitive exclusion rule is one of the possible explanations for the extraordinary species diversity in nature. If two populations in the same habitat cannot occupy the same niche, they must divide it. Over time each such population specializes in its own niche and may give rise to a new species.
Testing Gause’s rule in natural conditions is quite difficult, but some examples of its operation are known to biologists. Since the late 19th century, across much of Europe the broad‑clawed crayfish (Astacus astacus) has been displaced by the narrow‑clawed crayfish (A. leptodactylus). The niche of the narrow‑clawed crayfish overlaps that of the broad‑clawed, and the winning species is more fecund. The roach (Rutilus rutilus) displaces the common rudd (Scardinius erythrophthalmus) and the perch (Perca fluviatilis) in many water bodies. The adult niches are separated, while those of the juveniles overlap; the roach fry prove more competitive. However, the existence of many cases of competitive displacement does not prove that it always occurs. During the period in which biologists have studied Gause’s rule, both examples of its operation and situations where it fails have been found.
For example, in groups of planktonic algae niche partitioning is not observed. This is due to rapid environmental fluctuations, where transient conditions favor one species or the other, and also to intense predation pressure (zooplankton and various filter feeders) that prevents competitive exclusion from becoming the dominant factor in group formation. Yet in some cases competitive displacement does occur in planktonic assemblages. This happens during so‑called “water blooms,” when planktonic algae escape predation that normally limits their numbers. As a result of a bloom, only a few algal species become massive, each differing in its lifestyle, leading to competitive exclusion.
Some types of interactions can involve not only individuals from different populations but also relatives within a single population. For instance, intraspecific and interspecific competition are similar in that both limit the resources available to an individual. However, their effects on niche breadth are opposite. Intraspecific competition widens ecological niches: all individuals lack optimal resources for the species, and some are pushed to the periphery where only partially suitable resources exist. According to Gause’s rule, interspecific competition, on the contrary, narrows niches. Thus the observed niche widths in nature result from a balance of two opposing processes—expansion through intraspecific competition and contraction due to interspecific competition.
An interesting example of competitive exclusion was studied by Thomas Park with flour beetles. Two beetle species from the family Tenebrionidae (Tribolium confusum and T. castaneum) were kept in flour containers. These species competed for food (flour) and also could feed on each other, with beetles of both species primarily consuming individuals of the competing species rather than conspecifics (this combination of competition and predation is called antagonism). Park succeeded in finding conditions under which either the first or the second species consistently won. However, the most intriguing results were obtained under intermediate conditions that did not guarantee the advantage of either species (Table IV-13.1).
Table IV-13.1. Proportion of victories of the two flour beetle species in Thomas Park’s experiments
Repeating the study under identical conditions, the experimenter recorded victories of one species or the other with a certain frequency. Changing the rearing conditions altered each species’ chances of winning, but across a fairly wide range of temperature and humidity both species could win with a certain probability. One constant remained: after some time only one species persisted in the environment, the other disappeared.
Differences between species of the same guild (a set of species that use the same resource) can be mediated by predation. A conspicuous difference from a neighbor can draw a predator’s attention away from the focal species! In Southeast Asia, mesh pythons live in human settlements, steal and eat goats, pigs and dogs, and very rarely attack humans. This is because humans do not externally resemble their quadruped relatives and are not perceived by pythons as prey.
Intraspecific competition leads to niche expansion, pushing the niche toward its fundamental limits and beyond (through evolution).
The degree of competition between species can be assessed by assuming a certain limiting similarity threshold. If the differences between neighboring guild members are roughly equal at comparable levels of intraspecific variability, this indicates a strong role of competition in niche differentiation.
George Hutchinson demonstrated similarity in the divergence of morphological traits among species of the same guild by comparing the sizes of oral parts (tightly linked to prey parameters) in coexisting insects, birds and mammals. He found that homologous structures of each successive species are 1.1–1.4 times larger than those of the preceding one, on average 1.28 times larger. This size difference translates into a body‑mass difference of about 2 (2.09) times. Hutchinson’s rule concerns the sizes of feeding structures and whole bodies across a wide range of animals: spiders, jumping beetles, lizards, salamanders, squirrels, bats, desert rodents, frugivorous pigeons. It was shown that with a coefficient of variation (CV) of 5.5 for two species with a size ratio of 1.28, niche overlap would be 1–2%. Species are distributed along resource gradients non‑randomly. A ratio close to 1.3 is used to describe differences in series of tape recorders, machines and bicycles! Perhaps it reflects aspects of human perception, whereby engineers and marketers create product classes that “naturally” differ from each other.
It should be noted that, unlike machines and bicycles, representatives of species with larger body sizes often pass through size classes typical of smaller species during ontogeny. Hutchinson’s rule does not account for allometric growth patterns.
A guild can be imagined as an n‑dimensional space occupied by the ecological niches of different species. Under conditions of diffuse competition each species interacts with many others, competing with them across various environmental factors. Whether a species succeeds or fails to establish in a guild primarily depends on its competitive and exploitative relationships with other species. Imagine a box filled with inflated balloons: each presses on the others. If a population of a certain species (a balloon) “loses,” i.e., cannot reproduce (pops), its disappearance shifts the equilibrium among the remaining populations (moving all other balloons).
Conversely, pairs of species that overlap strongly along one niche dimension may differ markedly along another, thereby reducing competition. This can be described as differential overlap, where diffusely competing species partition the niche by different parameters. For example, predators often partition resources temporally because their foraging activity also follows a daily rhythm. For herbivores, temporal partitioning is less common. Poikilothermic animals use the day more differentially than homeotherms.
The closer the total abundance of guild members approaches the environmental carrying capacity, the more important competition becomes. One may roughly assume that competition is sharper at higher trophic levels.
The degree of niche partitioning among plants is generally lower than among animals.
Since intraspecific competition proves stronger than interspecific competition, the evolution of metamorphosis or ecological sexual dimorphism can be viewed as adaptations that reduce intraspecific competition.