Maro et al. (2008) Can the developmental conditions of tadpoles Bufo bufo...?
Attempt to understand the ontogenetic strategies of the common toads and a hypothesis about the mechanism of their switching. Pdf of the article here. Maro A. N., Shabanova A. V., Shabanov D. A. Can the developmental conditions of Bufo bufo tadpoles determine the rates of post‑metamorphic growth and maturation of toads? // Voprosy herpetologii. Mat...
For representatives of the genus Bufo, including the common toad (Bufo bufo (Linnaeus, 1758)), the formation of isolated local populations (breeding groups) closely linked to spawning water bodies is characteristic. Such breeding groups are characterized by a high degree of uniqueness. To study the reasons for such uniqueness, it is of interest to examine the consequences of artificial colonization of common toads in previously unoccupied habitats. Such colonization was carried out in 1994 with the participation of one of the authors of this work. This paper presents the results of morphometric study of 9 breeding groups of common toads (Table 1). All studied individuals were caught during spawning. Sampling sites are located in Zmievskyi (Nos. 1–3 in Table 1), Kharkivskyi (Nos. 4–8) and Chuhuiivskyi (No. 9) districts of Kharkiv region. Age of toads was determined by the number of growth rings in bone (Smirina, 1989). On a freezing microtome transverse sections of finger phalanges were prepared and stained with hematoxylin‑eosin according to Erlich. The number of lines of fusion in finger phalanges was determined for 52 individuals. In addition, this work uses results of long‑term marking (by finger‑phalange amputation) and recapture of common toads performed in the Iškov pond (No 3).
Toads taken from Iškov pond (No 3) in 1994 were introduced into two water bodies, Nos. 4 and 6. Subsequently, toads colonized and formed new breeding groups in neighboring water bodies independently. Pond 4 is the lower one in a system of connected ponds, therefore colonization of toads proceeded against the water flow. Ten years after introduction, toads only began to occupy the third of these ponds (No 5). From pond 6 colonization proceeded downstream through a system of seven ponds (pond 7 is the sixth of them) and occurred in the first years after introduction. Then toads crossed a watershed and colonized a pond in a neighboring ravine (No 8). The overall colonization sequence is: 3 → 4 → 5 and 3 → 6 → 7 → 8. Field observations of toad movement during spawning allowed us to state (Maro, Shabanov, 2005) that colonization occurs when individuals heading to spawn fall into another water body located on their route. In addition, tadpoles can be carried by water currents. Comparison of toads from long‑existing and recently formed populations showed significant differences in their size structure (Table 1, Fig. 1). For example, comparison of size distributions of toads from ponds 3 (“parental”) and 8 by the Kolmogorov‑Smirnov test yields significant differences (p < 0.001 for both females and males). Comparing combined data from long‑existing populations (Nos. 1–3) with those formed as a result of artificial colonization (Nos. 4–8) also shows significant differences (p < 0.005 for females and p < 0.001 for males). Breeding groups in recently colonized habitats are characterized by relatively larger females and the participation of very small males belonging to size classes that do not take part in breeding in long‑existing populations. This suggests that in newly settled habitats male common toads mature faster, while females grow faster. Skeletochronological studies confirmed this hypothesis (Fig. 2). Fig. 1. Size structure of the studied toad samples. Characteristics of each population are shown on the plot by a cross. Designations: a — mean body length of females and males; b and c (bold lines) — variability range encompassing 50 % of individuals for females and males respectively (upper and lower quartiles); d and e (thin lines) — minimum and maximum size values for females and males respectively.
Table 1. Sampling sites, sample size (N) and features of size structure of studied common toad samples (L — mean body length, s — standard deviation of body length).
No.
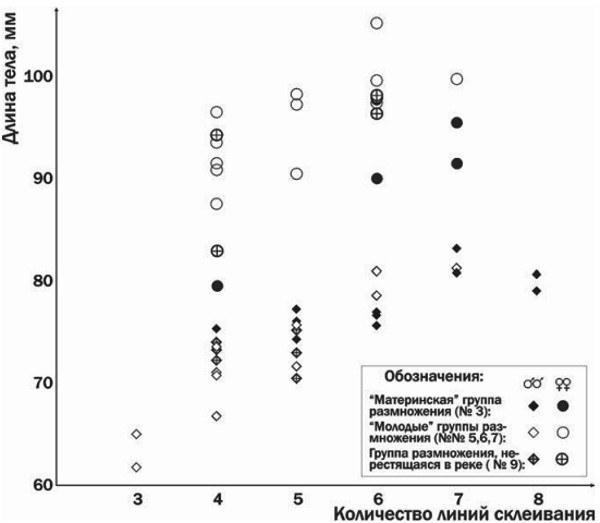
Fig. 2. Relationship between body length and age (number of lines of fusion in finger phalanges) for toads from different breeding groups
As seen in Fig. 2, females from young breeding groups, although younger (having fewer lines of fusion) than females from the parental breeding group, are either similar in size or even larger. Among males from recently colonized habitats, both individuals comparable in size and age to those from the parental group and substantially younger and smaller ones are recorded during spawning. In the skeletochronological studies shown in Fig. 2, the number of resorbed lines of fusion in the medullary cavity was not assessed, and therefore the values presented in Fig. 2 represent lower estimates of each individual’s age. It should be noted that toads from the parental breeding group are characterized by a larger endosteal cavity. This may be a consequence of relatively longer development before reaching sexual maturity, leading to resorption of a greater number of lines of fusion. In this case, the age difference between toads from long‑existing and recently colonized habitats may be even greater than shown in Fig. 2. Supporting this hypothesis are, among other things, the results of long‑term marking (by finger‑phalange amputation) of common toads in the “parental” breeding group (Iškov pond, No 3 in Table 1), shown in Fig. 3.
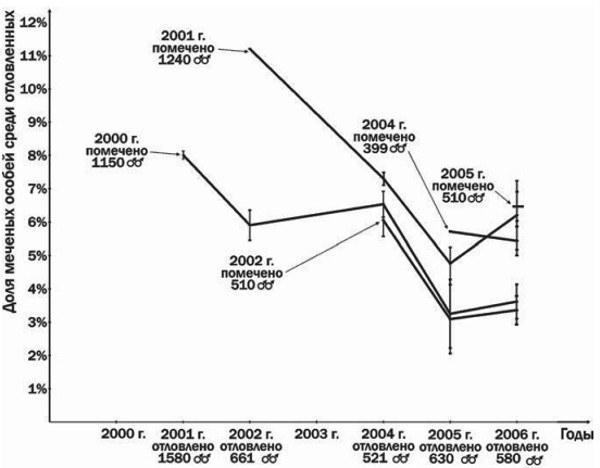
Fig. 3 shows that males participating in spawning in 2000 and 2001 constituted a substantial part of the breeding group even after 6 or 5 years, respectively. In addition, the results presented in Fig. 3 indicate a decline in the size of the Iškov pond breeding group over the observation period. This may be a consequence of the Iškov pond being drained during the 2000 spawning of common toads (Ačkasova et al., 2001). Consequently, the breeding group of common toads in Iškov pond is characterized by low growth rates combined with multi‑year (up to 6–7 times) participation in spawning.
Fig. 3. Results of marking and recapture of male common toads from Iškov pond. Vertical bars indicate the maximum and minimum return estimates, depending on the interpretation of insufficiently distinct marks. For each year the number of captured and marked individuals is given.
The presented data provide a basis for the hypothesis that in the parental population males reach spawning at a later age but are more competitive. In a competitive environment, small males will have low chances of fertilizing a female. A slower growth of females in a competitive environment may also be adaptive, because in amphibians and fish the largest and usually slower‑growing individuals do not reach the maximum age (Mina, Klevézal, 1976).
Thus, it can be assumed that development of toads in newly colonized habitats promotes an increase in their numbers, whereas in long‑established habitats it enhances their competitiveness. This difference in individual development strategies can be compared with the r‑ and K‑strategies described by R. MacArthur and E. Wilson (Pianka, 1981). It should be noted that here we are not comparing strategies of different species, but demographic strategies of populations and ontogenies of individual members of the same species.
Features of the common toad population spawning in the channel of the Northern Donets River at a site with relatively strong current (No 9) suggest that in flowing water the influence of high tadpole density on post‑metamorphic ontogeny of toads is reduced. The described breeding group has existed for a long time and is characterized by high abundance and density of spawning toads, but has a size structure typical of “young” breeding groups: it combines small males and large females (Fig. 1). One of the significant factors of interaction among tadpoles that determines their future ontogeny may be the release into the water of various substances influencing their growth and development (Schwartz, 1972 and other works).
Based on the above, we propose the following hypothesis. The rate of post‑metamorphic growth and the age at sexual maturity of common toads is determined by tadpole density in the water bodies where they develop. Low tadpole density promotes toad development that maximizes fecundity in the coming years. Males in such populations participate in breeding at a relatively young age and at small sizes; females exhibit high growth rates and probably relatively short lifespans. Where tadpole density is high, the pattern of post‑metamorphic development of toads reflects maximization of total productivity over the whole life in a competitive environment. In such habitats toads develop and grow more slowly, but live longer and become ready to spawn prepared for intense competition. In particular, male readiness for sexual selection is manifested by reaching sexual maturity at larger sizes.
Further studies, currently underway, can confirm or refute the proposed hypotheses.
The authors thank E. M. Smirina, S. M. Liapkova and V. G. Ishchenko for valuable criticism, but do not hold them responsible for the assumptions expressed in the article. The authors also thank A. V. Korshunov, M. A. Kravchenko and G. A. Mazepa for assistance in field work.
Ačkasova I. V., Dyachenko L. A., Mykota Ya. Yu., Shabanova H. V., Shabanov D. A., 2001. Size structure of the population of gray toads (Bufo bufo)// Biologia ta Valiologia. 36. scientific works. Kharkiv: KhDPU. Issue 4. pp. 41–46.
Maro A. N., Shabanov D. A., 2005. Mechanisms of colonization of the gray toad (Bufo bufo (L. 1758); Amphibia, Anura) and features of its populations in recently colonized habitats // Materials of the First Conference of the Ukrainian Herpetological Society. K.: Zoological Museum of the NNP of the NAS of Ukraine. pp. 107–110.
Mina M. V., Klevézal G. A., 1976. Animal growth. Moscow: Nauka. 291 p.
Pianka E., 1981. Evolutionary ecology. Moscow: Mir. 400 p.
Smirina E. M., 1983. Post‑mortem age determination and retrospective body‑size assessment of the gray toad (Bufo bufo)// Zool. J. 62(3): 437–444.
Schwartz S. S., 1972. Metabolic regulation of growth and development of animals at the population and organismal levels // Izvestija AN USSR. Series Biol. No 6. pp. 142–151.