
Three Articles on Interspecific Recombination
Contagious Genes, or the Horizontal Turn: DNA can be acquired not only from parents. Tracing the genealogy of any microorganism, one can see that a "river" of genetically heterogeneous information flows through it. The entire biosphere is reflected in this stream! Wolbachia: strangers...
Contagious Genes, or the Horizontal Turn: DNA Can Be Acquired Not Only from Parents "...All living matter stands before us as a single whole, as one enormous organism, borrowing its elements from the reservoir of inorganic nature, purposefully governing all processes of its progressive and regressive metamorphosis, and ultimately returning everything borrowed to dead nature." S. N. Vinogradsky, Lecture before the Imperial Family, 8 December 1896 The journal Nature publishes not only articles on accomplished achievements, but also essays in which the authors attempt to chart further paths for the development of science. One of the most recent belongs to Nigel Goldenfeld and Carl Woese, American scientists who predict yet another revolution in biology. From their perspective, new data on the exchange of genetic information between members of different species will bring about a paradigm shift in biology. The subject concerns horizontal gene transfer — the transmission of hereditary information from organism to organism outside the genealogical (vertical, from ancestors to descendants) sequence. The phenomenon itself has long been known: for example, genes conferring antibiotic resistance are transferred in this way from one pathogenic microorganism to another. For many years it was viewed as a kind of blunder of nature. Enthusiasts hoped to explain with it the phenomenon of evolution; pragmatists sought in it natural technologies for genetic engineering… Only the recent avalanche of molecular-biological data has made it possible to understand how widespread this phenomenon is in nature. For decades, microbiology was dominated by the principle enshrined as a dogma by Robert Koch: the microorganism under study must be grown in pure culture. Frequently, the descendants of a single bacterial cell are used for cultivation. Microorganisms grown under such conditions prove identical, which is regarded as a criterion for their correct isolation. Alas, such conditions are entirely unnatural! What is natural is precisely the functioning of microorganisms in a complex environment, the informational exchange with which includes the transfer of DNA fragments. Under such conditions, for the two principal groups of "microbes" — bacteria and archaea (or archaebacteria) — it is altogether difficult to speak of a species-specific genome (the set of hereditary information). Thanks to horizontal transfer, this genome is constantly replenished with new fragments, and thanks to its own evolution loses much of what has been acquired. Tracing the genealogy of any microorganism, one can see that a "river" of genetically heterogeneous information flows through it. The entire biosphere is reflected in this stream! How is horizontal transfer of genetic information detected? Sometimes one can observe the original sequence in one organism and its copy in another. More often, however, indirect evidence must be used — identifying DNA segments that differ in the ratios of different nucleotide pairs or in the frequency of use of different triplets for encoding the same amino acids (these features are, in general, species-specific). Finding in someone's genome a stretch of text that differs from its surroundings in the frequency of "letters" (nucleotides) or "words" (triplets), one can hypothesize that a borrowed sequence is at hand. Over time, the host will correct the text, bringing it into conformity with its own "style."
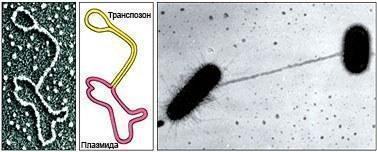
Thus, the work of modern genetic engineers creating genetically modified organisms — and thereby generating a wave of protests and alarming stories — turns out to be an ordinary matter for the Earth's biosphere. What is most interesting is that horizontal transfer is characteristic not only of microorganisms. For example, recent studies by researchers at Berkeley of the genomes of rice and millet (cereals separated by at least 30 million years of independent evolution) showed that horizontal transfer of genetic information had taken place between these species. Virtually identical transposons were found in these plants — segments of DNA capable of moving within and between genomes! Most often transposons transfer information within a species, but they can also cross the species barrier, since the boundary between transposons and viruses is quite conditional. The reorganization of the genome that occurs during speciation may be associated with the activity of modified transposons. Hot-headed thinkers see in horizontal transfer an explanation for progressive evolution. New traits arise like this: the wind blows, brings a virus with a piece of new information, and — lo and behold! — the organism acquires a new beneficial property. Thus the outstanding palaeobotanist V. A. Krasilov considers that in the epoch of the origin of flowering plants, it was precisely viruses that carried "blocks" of genetic information responsible for flower formation from one gymnosperm to another. Alas, this is not easy to believe. The reason is that the design of any organism is a difficult compromise between modularity and monolithism. Any creature inhabiting the Earth is simultaneously a coherent system and a conglomerate of relatively independent traits. Those properties of an organism that are bound to all the others in a tight knot of interrelations cannot be "caught" from the wind. This is precisely why the role of horizontal transfer in the evolution of highly organized and highly integrated groups decreases. Perhaps one day genetic engineers will get around to testing Krasilov's idea and will transfer the "flower-formation genes" into the genomes of pines, ginkgos, or cycads? Most likely, the foreign fragments will be unable to integrate into the developmental regulatory system of a plant characterized by a different mode of reproduction. Even when a major novelty in highly integrated animals turns out to be associated with some product of horizontal transfer, it should not be regarded as a direct consequence of the capture of foreign information. Let us give one example. In mammals, the development of the placenta (the organ providing the physiological connection between the embryo and the mother) requires, among other things, the activity of the Peg10 gene. This gene is extraordinarily similar to one of the widely distributed transposons. This fact can be interpreted in two ways. A naive individual, convinced that every trait is the projection of a particular gene, will conclude that the emergence of placental mammals (and ultimately of ourselves) is the consequence of "infection" with the "placenta gene." From there it is a short step to belief in a genetic Demiurge who governs evolution by periodically launching new viruses and new progressive qualities into circulation. A more mature interpretation presents itself differently. New functions developed in the course of evolution can be connected not only with old segments of the genetic text that perform their own tasks, but also with new, free ones. These new genes may turn out to be pieces carried in from outside. Just as an experienced driver, repairing a car that has broken down on the road, may fashion a required part from some fragment found by the roadside. The function of that fragment in the repaired structure was not contained in it originally — it arose as a consequence of its particular position in the new system. This interpretation is supported by the fact that the transposon closely related to the "placenta gene" in any other location and context does not induce the formation of anything similar. On the contrary, horizontal transfer serves not as a driver, but rather as a brake on evolution! Organisms that have acquired a relatively isolated genome evolve morphologically, physiologically, and behaviourally far more rapidly than microorganisms tightly woven into the unified network of the biosphere. Evolution is directed from a continuum of genomic possibilities toward individual genotypes. Why did biologists understand this so late? There are several reasons. First, biology had to reach a technical level permitting the registration of the very fact of the diversity of pathways for transmitting hereditary information. Second, the study of supra-organismal systems is psychologically difficult for researchers who are themselves organisms (see the sidebar). Third, the spread of new understanding is impeded by the interpretation of the organism as the embodiment (realization) of a genetic program. The metaphor of hereditary information as a program has become so deeply embedded in the consciousness of modern biologists and non-biologists that many of them find it simply difficult to grasp its implications. And what is the organism, if not the embodiment of the plan stored in its genes? That which uses those genes! Hereditary information is not the true essence of the organism, but a library it uses. For most of the history of life, this library was shared by the most diverse creatures, and only later did some of them acquire their own isolated "bookshelves." When we speak of humans or our closest relatives, the higher animals, we are dealing with relatively isolated species. Each of them is characterized by a certain commonality of genetic program, occupying a definite place in ecosystems. The discreteness of bacteria and archaea is of a different nature. It is more closely associated with the discreteness of possible modes of life, of ecological niches. Thus, there is every reason to agree with the authors of the essay in Nature. We stand on the threshold of a new understanding of life around us. The concepts of organism, species, genome, and hereditary information require serious clarification. At the same time, this approach should not be regarded as entirely new. It was this that Alexander von Humboldt was groping toward in his Cosmos, that Sergei Nikolaevich Vinogradsky expressed in his lecture before the royal family, that James Lovelock embodied in the Gaia theory, that the academician of the Russian Academy of Sciences Georgii Aleksandrovich Zavarzin is attempting to develop under the name of "naturalistic microbiology"… But in any case, the development of biology will yet yield many more surprises. Sidebar: How to Encompass the Boundless We ourselves are organisms, and we perceive the other systems around us by analogy with ourselves. However natural this approach is for us, it is not the only possible one… The spring before last, the author of these lines found himself during the spawning of moor frogs near a small spawning pond — a pool among oak-covered hills. It was twilight — the "hour between the dog and the wolf." Several thousand male frogs had gathered in the shallows. At this time they are transformed by their bright blue breeding coloration. The call of a male moor frog resembles the gurgling of water boiling in large bubbles. The polyphonic chorus of several thousand males merged into an incredible roar. This united voice of thousands of individuals spread across the surrounding hills, summoning the unhurried, spawn-swollen females… Approaching the spawning aggregation, a female finds a mate and together with him deposits her portion of spawn. Those females that arrive later than others are forced to spawn on a continuous field of egg masses. Some of their eggs are fertilized by the milt not of "their own" male, but of others — those that were fathers of the adjacent clutches. A little time will pass, and tadpoles will emerge from the eggs; their hatching enzymes will liquefy the mucous egg membranes. Even if, during the course of their development, the water level in the spring pond drops, the liquefied spawn will flow downhill toward the receding water. Let tens of thousands of individuals (and a tadpole, like an egg, is an individual!) dry out on the margins of the pond — hundreds of thousands will enter the water, where they will continue to develop. As they grow, they will exchange diverse chemical and physical signals. When necessary, those tadpoles that have outpaced the others will slow the growth of the laggards in order to reduce competition for limited resources. In other cases, on the contrary, the growth of larvae of different ages and kinships will be synchronized — the population itself will choose the strategy of its optimal development. So it was that the twilight, the calling of frogs, and fatigue shifted the perception of the person who had approached the spawning aggregation. He felt that he was standing not before a certain number of organisms, but before an entity of a higher order — a population. A potentially immortal living object was at one of the key stages of its annual cycle. The production of new individuals justifies the losses of existing ones, for the being of the population is realized only through their flow and renewal. It would seem that a person cannot be noticed by this entity — he is, after all, at a lower level of being than it.

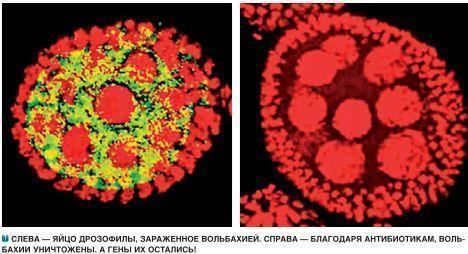
Nevertheless, when the entranced person approached one of the spawning aggregations, the calling frogs took fright, fell silent, and hastily hid themselves on the bottom. The voice of the population changed: it had noticed the stranger and responded to his presence… You will say that such an experience of contact with an entity of a higher order belongs to the sphere of interest of a psychiatrist, and not a biologist? You are, of course, right. But, believe it or not, such experience is very helpful in imagining the population as a unified whole, rather than a mere assemblage of separate individuals… And you would do better to try to imagine the planetary community of living organisms, bound together by horizontal transfer of genetic information, as the population of frogs is bound together by communal reproduction! Is it convenient to describe and study this community using traditional concepts and commonplace mental stereotypes? New tasks demand new modes of thinking! 1 At the same time, it is true, one of the arguments with which genetic engineers attempt to allay the fears of alarmed society — that "foreign genes" cannot possibly spread through the biosphere — loses credibility. It turns out that things are not so obvious. However, the inserts into the genomes of modified organisms are also products of the biosphere's development, just as the "native" genes of cultivated plants are. Back to the text Wolbachia: Strangers Within the Genome Kompyuterra has more than once mentioned the name of Craig Venter, the American multi-millionaire who amuses himself by sequencing the human genome (or more precisely, his own genome) and creating artificial life from modified mycoplasmas ("minimalist" bacteria). Recently, the staff of the Institute for Genomic Research he founded made yet another discovery — one less of a technological than of a worldview character. The article by the authorial collective (twenty names) reports the discovery of genes of the bacterium Wolbachia in the genomes of insects and nematodes. To appreciate the discovery, it is necessary to discuss the distinctive features of this bacterium — an endosymbiont and intracellular parasite of many invertebrate species. Let us review what was known about Wolbachia up to the present time. Wolbachia lives in the cytoplasm of the cells of its hosts: insects, nematodes, crustaceans, and other invertebrates. Sometimes the cells of insects are simply packed with these parasites! The effects of Wolbachia on its hosts can be highly diverse — sometimes it does not harm them, but for the most part it causes serious damage. For example, a typical manifestation of bacterial activity is the killing of the host's eggs containing male embryos (the so-called andricide, or male killing). Paradoxically, the hosts do not die out thereby. First, the reproductive potential of the population depends on the number of females, not males. Even a handful of male individuals can fertilize a multitude of females. Second, Wolbachia can take care of the reproduction of infected females by endowing them with the capacity for parthenogenesis — virgin reproduction. By disadvantaging males, Wolbachia in a sense helps females. Insects infected with this bacterium can double their lifespan and substantially increase their fecundity! It seems that individuals free of these bacteria can achieve the same level of viability, but in any event the removal of Wolbachia inflicts damage on lines that have become "addicted" to it. Why does Wolbachia wage war against males? The offspring of infected females will be infected, since Wolbachia will be able to pass into the eggs. But spermatozoa are too small for these bacteria. Males are "unnecessary" to the parasite, and so it kills them. But this is not the only strategy of Wolbachia. For some hosts (for example, woodlice, terrestrial crustaceans), it has been shown that the bacteria can keep males alive, simply transforming them into females, despite the male chromosomal complement. The offspring of transformed males will be the same — females with male genes forming a parthenogenetic clone infected with Wolbachia. In experiments, it was possible to transform such woodlice back into normal males by treating them with antibiotics and ridding them of the intracellular parasites. Another of the solutions used by Wolbachia to intensify its transmission is the control of the fertility of infected males. Their spermatozoa lose the ability to fertilize the eggs of females uninfected or infected by other Wolbachia clones (let those not reproduce), but interact perfectly with eggs carrying the same infection! In short, the basic hypothesis of researchers of this phenomenon, termed cytoplasmic incompatibility, comes down to the idea that Wolbachia creates in spermatozoa a certain controlled defect, which can be corrected by the activity of bacteria residing in the egg. What a strategy! Recall science fiction films and books — could human imagination devise something similar? One of the consequences of the sophistication of Wolbachia's strategies was the so-called infectious speciation — the division of a host species of this bacterium into a multitude of genetically isolated parthenogenetic or bisexual lineages. As a result of independent evolution, such lineages can over time develop into distinct species. Is this not one of the secrets of the species richness of insects and nematodes — the most numerous classes of organisms on our planet? It is no coincidence that Wolbachia inhabits the cells of approximately 70% of extant animal species! Incidentally, under laboratory conditions Wolbachia lives in cultures of human cells, but under natural conditions these microorganisms have, fortunately, not made the transition to parasitizing mammals. The study of the relationships of this mind-boggling bacterium with the cells of its hosts may be the key to solving a broader problem — the origin of eukaryotic organisms. The cells of humans, as of all other animals, plants, and fungi, are powered energetically by the activity of mitochondria — relatively autonomous cellular organelles. Molecular studies have convincingly demonstrated that mitochondria are the descendants of certain alphaproteobacteria, one of the bacterial groups still flourishing today. Wolbachia belongs precisely to this group. Mitochondria live in the cytoplasm of our cells, reproduce by division, and even possess their own circular DNA and protein synthesis machinery. When any of us says "I" about our body, we imply not only the sum of organs, tissues, and cells, but also the endosymbionts inhabiting all those cells — descendants of once-independent organisms with their own evolutionary fate. However, there can be no question of the self-sufficiency of mitochondria — the greater part of the proteins they require are encoded in the nuclear genome. For a long time this circumstance was a favourite argument of opponents of the endosymbiotic origin of mitochondria. And the difference between Wolbachia and the bacterial ancestors of mitochondria was explained precisely by the claim that Wolbachia is incapable of transferring its genes into nuclei. This assumption proved unfounded. Let us return, however, to the discoveries of the staff of Venter's Institute. They found that even after treating Drosophila cells with an antibiotic that killed the Wolbachia inhabiting them, genetic tests for the presence of bacterial DNA in the cells of such Drosophila continued to yield positive results. Studies of this phenomenon showed that the Wolbachia genome can simply become incorporated into the host's genome. Fragments of the genetic information of this bacterium have been found in the cells of four insect species and four nematode species; the size of the fragments ranged from almost the complete genome (one megabase in length, one million base pairs) to relatively small pieces. The ability of viruses to integrate into the genome of their eukaryotic hosts and subsequently exit from it has been known for a long time. Kompyuterra has already reported that such viruses can transfer fragments of genetic information to new hosts from their previous "sites of residence." That bacteria are capable of a similar trick (at least with regard to the integration of their hereditary information into the host's genome) has been discovered for the first time. And can Wolbachia transfer blocks of the host's genetic information from place to place? Bear in mind that the "payload capacity" of a bacterium, possessing its own cell, is considerably greater than that of a virus — a conglomerate of a few molecules. If it can, then who (or what), properly speaking, governs the evolution of the overwhelming majority of species of terrestrial animals? However, let us dispense with mysticism. It is highly probable that what makes Wolbachia marvellous and enigmatic to us is precisely its relative degree of study.
Are you certain that in the living organisms beneath our feet even greater wonders do not lurk, as yet unnoticed by us? Recall the exceptional capabilities of Wolbachia — from andricide and the provision of cytoplasmic incompatibility to the feminization of males, the induction of parthenogenesis, and the extension of host lifespan. Presumably, the biochemical "keys" to these capabilities are encoded in its genome. And the hosts submissively incorporate this entire toolkit for hacking their species strategy into their own hereditary apparatus! Although… Perhaps the reason for Wolbachia's marvellous capabilities lies not so much in itself as in the organisms it infects? Perhaps the bacterium merely opens "secret doors" built into the hosts' design by a higher power? With respect to parthenogenesis or the extension of lifespan, this is not so difficult to believe. Feminization of males — more difficult, but also conceivable: after all, male embryos have the latent potential to become females. Andricide? Who knows… In what situation might such a capacity be advantageous for a species? Cytoplasmic incompatibility? It seems that realizing it without cooperation between parasite and host would be impossible. But even if the bacterium simply "chooses" among several alternative developmental strategies of the host, how does it make that choice, how does it switch off the other pathways? …Children on the younger side who attend school, and children on the older side who attend university, are told that an organism is the result of the realization of a particular genetic program, and that evolution is the process of change in gene frequencies in populations. Convincing, isn't it? The Glacier as a Time Machine Kompyuterra has already reported (see #675) that terrestrial microorganisms share a common gene pool — a pool of hereditary information. Presumably, over time this planet-wide gene pool gradually changes. New fragments of information are "invented," and some of the old ones are lost irretrievably. How irretrievably? Could they be preserved somewhere? Such a "refrigerator," where old genetic information would be protected from decay, could be a glacier. Recently, a number of reports have been published on the revival of bacteria contained in glaciers. But what is the limit of how long living organisms or their genetic material can be stored in ice? Generally speaking, almost any glacier can be compared to a river of ice. Under pressure, ice becomes fluid. In the "headwaters" it forms from snow and hoarfrost; in the "lower reaches" it melts or (as is typical for Antarctica) breaks off in chunks — icebergs — into the ocean. Therefore, almost everywhere on our planet the age of the oldest ice does not reach one million years. In Greenland, for example, record sections of ice aged 800,000 years were recently found, and bacteria were discovered in them, incidentally. But now an American-South Korean research team has reported the finding and investigation of ice samples that they consider the oldest on the planet — 8 million years old. These fragments were collected in the folds of the Transantarctic Mountains: geological traps have retained ice from the Miocene epoch! (Incidentally, if one examines the map of the sample collection points, it can be hypothesized that further down the glacier flow the ice may be even older.) This illustration from the article under discussion brings together a multitude of data: the geographical location of the study area and sample collection points (the age of the first sample is one hundred thousand years, the seventh approximately eight million); a photograph of the site, as well as optical and fluorescence images of rounded and filamentous bacteria. From the samples studied, DNA of thirty different microbial forms was isolated. They proved to be closely related to the sequences of known bacteria from very diverse groups and even kingdoms (both eu- and archaebacteria), but were not identical to modern forms.

Most importantly — it proved possible to restore them to life! Metabolic activity of the fossil bacteria was registered — for example, the incorporation into their composition of a nutrient medium containing "labelled" (radioactive) atoms. While samples from relatively "young" ice grew rapidly and formed new colonies, the growth of bacteria from the oldest samples was extremely slow and unstable. This is not coincidental: during the time they had spent in a frozen state, the organisms received an enormous radiation dose. It was even possible to determine the half-life (that is, the period of 50% degradation) of bacterial DNA under glacial conditions: it amounts to 1.1 million years. The result obtained means that global warming, by causing glaciers to melt, will introduce into the biosphere microbial material from the past. Could this have an adverse effect on the current state of affairs? Fortunately, the concern does not extend, for example, to microorganisms causing diseases of mammals — there is nothing for such organisms to do in Antarctic and Arctic snows. The fragments of genetic texts being returned to the biosphere should be considerably damaged. If one recalls that pieces of past genetic information are already stored in the genomes of the most diverse organisms, and imagines how many are being transferred and modified in the course of mutations, it becomes clear that the return of fossil information from melting glaciers should not exert any significant impact on the biosphere. But it is interesting nonetheless… D. Shabanov. Contagious Genes, or the Horizontal Turn // Kompyuterra, Moscow, 2007. – No. 7 (675) D. Shabanov. Wolbachia: Strangers Within the Genome // Kompyuterra, Moscow, 2007. – No. 34 (702). D. Shabanov. The Glacier as a Time Machine // Kompyuterra, Moscow, 2007. – No. 30 (698). — P. 12