Shabanov (2012) The Hybridogenetic Complex of Water Frogs as a Model for Studying Multilevel Selection
Shabanov D.A. The hybridogenetic complex of water frogs as a model for studying multilevel selection // Bulletin of the Vasyl Stefanyk Precarpathian National University. Series - Biology. - Ivano-Frankivsk: Publishing House of the Vasyl Stefanyk Precarpathian National University, 2012. - Issue ...
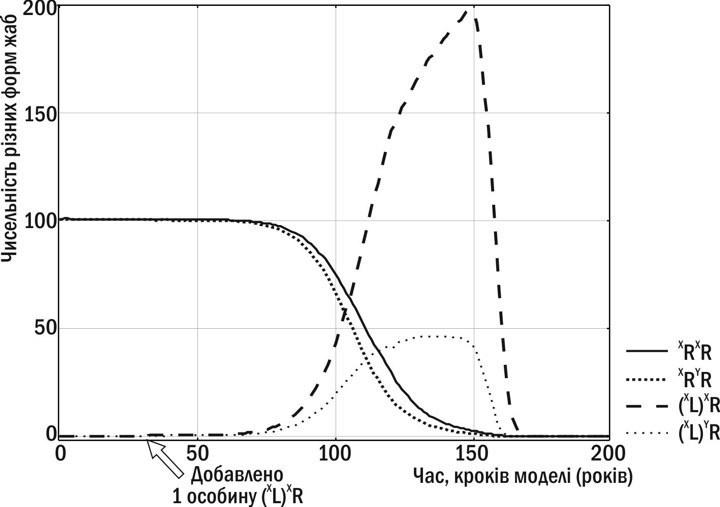
THE HYBRIDOGENETIC COMPLEX OF WATER FROGS AS A MODEL FOR STUDYING MULTILEVEL SELECTION Shabanov D.A. V.N. Karazin Kharkiv National University, Department of Zoology and Animal Ecology, e-mail: d.a.shabanov@gmail.com The observation and modelling of the evolution of the hybridogenetic complex of water frogs (Pelophylax esculentus complex) reveal phenomena that, for their explanation, require drawing upon diverse conceptual frameworks. Among these are the concepts of the "selfish gene", the "selfish individual", and group selection. These concepts do not contradict one another but are complementary (in the sense of Niels Bohr's complementarity principle). Consequently, this hybridogenetic complex constitutes an apt model for the study of multilevel selection. Key words: Pelophylax esculentus complex, hemiclonal inheritance, hemiclonal population systems, "selfish gene", "selfish individual", group selection, multilevel selection. D. A. Shabanov. Hybridogenetic complex of water frogs as a model for multilevel selection studies. Monitoring of hybridogenetic complex of water frogs (Pelophylax esculentus complex) and modeling of its evolution reveal phenomena that may be explained through the concepts of "selfish gene", "selfish individual" and "group selection". In fact these concepts are not contradictive and complement each other (in terms of the complementarity principle by Niels Bohr). That is why this hybridogenetic complex is a good model for multilevel selection studies. Key words: Pelophylax esculentus complex, hemiclonal inheritance, hemiclonal population systems, "selfish gene", "selfish individual", group selection, multilevel selection. Introduction One of the most contentious issues in theoretical biology, which has generated vigorous scholarly debate in recent decades, is the determination of the level at which natural selection operates. Since the time of Charles Darwin, it was held that the individual organism constitutes the unit of selection (4). The architects of the Modern Synthesis emphasised populations as the arena in which changes in allele frequencies occur, linked to the survival and reproduction of organisms. In explaining the origin of altruism, Darwin suggested that it results from the enhanced survival of human groups rather than of individual altruists (3). V. C. Wynne-Edwards termed this view group selection and attempted to explain a wide range of organismal traits through it (18). Wynne-Edwards' arguments were substantially refuted by G. C. Williams (16). The crux of the matter is that in situations where "group interests" require one course of action while individual interests dictate another, individuals that pursue individual interests will increase the proportional contribution of their alleles to the future gene pool of the population. For example, if a population exists under conditions of acute resource scarcity, one might suppose that for the population's stable persistence it would be advantageous if all individuals reduced their reproductive expenditure. Yet any individual that does not reduce (or indeed increases) its reproduction will augment the proportion of its descendants in future generations. Natural selection will favour precisely such individuals. After some time the pendulum swung in the opposite direction. Through the work of J. B. S. Haldane, W. D. Hamilton, J. Maynard Smith, and, to a considerable degree, the brilliant populariser of their collective views R. Dawkins, the concept of the "selfish gene" gained broad currency, viewing genes as the principal units of selection (5). From this perspective, organisms are merely vehicles through which genes replicate themselves. The principal argument in favour of this concept was the notion of kin selection and inclusive fitness of the gene, developed by W. D. Hamilton in the course of his research into the evolution of altruistic behaviour in social insects (13). At present, no consensus has been reached on this contentious question. A considerable number of biologists, particularly molecular biologists, regard the "selfish gene" concept as definitively established. They are opposed by specialists in classical disciplines who work with organisms and consider this concept a reductionist oversimplification. Their views may be termed the concept of the "selfish individual". In recent times, disputes concerning which level of natural selection is primary have intensified markedly. The doyen of evolutionary biology and eminent myrmecologist E. O. Wilson began reviving ideas of group selection. These ideas found full expression in the monograph "The Social Conquest of Earth", published in 2012 (17). These views are grounded in a detailed description of the transitional stages in the evolution of sociality in insects, as well as in the results of modelling evolutionary processes. Such positions provoked fierce criticism from R. Dawkins (12). On the whole, it may be considered that 2012 witnessed a resurgence of interest in debates concerning the primary level of selection. Further advancement in the study of the problem outlined above requires the introduction of new arguments. In the author's view, it is desirable to investigate this problem using new model systems that go beyond the bounds of the traditionally employed examples (and, if possible, do not concern the evolution of altruistic behaviour, as that is an unduly complex question). This paper discusses the criteria that should be demanded of an adequate model for investigating which level of selection is primary. This discussion affords the opportunity to propose whether the notions of the "selfish gene", the "selfish individual", and group selection are genuine alternatives. Finally, the paper substantiates why the hybridogenetic complex of water frogs constitutes a suitable model for the study of multilevel selection. Main Body Alternative or complementary concepts? It should be noted that the very designation of the "selfish gene" concept is not entirely accurate. According to this view, evolutionary advantage accrues to those variants of genes (alleles) whose effect most efficiently increases the proportion of their copies in subsequent generations. It is for this reason that these views are better termed the concept of the "selfish allele", henceforth SA. By analogy, we shall employ the abbreviation SI for the concept of the "selfish individual", and SG ("selfish group") for the concept of group (or interdeme (2)) selection. How is one to determine which of the concepts (SA, SI, or SG) is correct in any given case? A considerable portion of the disputes between proponents of these views stems from the fact that all of these concepts describe a large number of processes adequately. Let us consider a simple example. We consider two groups consisting of individuals that reproduce and transmit their genes to descendants. One group grows more rapidly because the individuals comprising it reproduce (and transmit genes) more efficiently than individuals of the other group. The processes in this example are equally well described by the concepts SA, SI, and SG; observation of such processes does not permit selection of a more adequate explanation. But this is not always the case. In the Introduction, brief consideration was given to the example in which selection at the group level would favour reduced fecundity, while selection at the individual level would favour increased fecundity. In such cases it becomes possible to determine which level of selection proves more effective. Observation of model biosystems upon which selection at different levels operates in opposing directions affords the possibility of choosing between competing concepts. In the example under consideration, the registration of a decline in fecundity constitutes evidence for SG, while its maintenance at the previous level constitutes evidence for SI or SA (the two concepts yield identical predictions). Thus, in order to resolve the problem under discussion, cases should be sought in which selection at different levels is oppositely directed. For instance, meiotic drive (2) may be regarded as a classic example of SA in action - a case in which a particular allele increases the probability of its entry into gametes. In a number of examples it has been demonstrated that such alleles do indeed spread in nature, disrupting the reproduction of organisms and the perpetuation of populations. Thus, at least in some cases, application of the SA concept is indispensable. There is also a well-known example that cannot be explained from the perspective of SA. The fact is that in organisms with sexual reproduction and meiosis occurring during gametogenesis, each allele has (in the typical case) only a 50% probability of entry into gametes. This mechanism considerably reduces the effectiveness of selection at the gene level and, notwithstanding this, is widespread throughout the biosphere. It can be explained only at the level of SG (the Vicar of Bray hypothesis and other hypotheses) or SI (the Red Queen hypothesis and others) (10). Since modern science is aware of phenomena explicable only from the perspective of one of the enumerated concepts, it can be established that these concepts are not alternatives. These concepts are complementary (in the sense of Niels Bohr's complementarity principle), that is, they appear to be mutually contradictory but in reality explain different aspects of the process under study (1). Features of Pelophylax esculentus complex. The hybridogenetic complex of Central European water frogs, Pelophylax esculentus complex, consists of the pool frog, Pelophylax lessonae (Camerano, 1882), the marsh frog, Pelophylax ridibundus (Pallas, 1771), and their hybrids. For these hybrids, a species-like name is applied: the edible frog, Pelophylax esculentus (Linnaeus, 1758) (15). In the typical case, during gametogenesis in P. esculentus, one of the genomes (haploid chromosome sets) of a parental species - either P. lessonae or P. ridibundus - passes into the gametes they form. Let us consider this with a simple example. In water bodies situated in the city of Kharkiv, representatives of P. ridibundus and P. esculentus cohabit. Designating the genome composition of their genotypes, we shall use the letter L for the P. lessonae genome and R for the P. ridibundus genome. It can be indicated that in this locality frogs of genotype RR and LR reside. Clearly, representatives of P. ridibundus produce gametes bearing the R genome. The case of P. esculentus is more complex. In Kharkiv, diploid P. esculentus are prevalent (in certain other localities, for example in the Siverskyi Donets centre of water frog diversity, triploid individuals are also widespread (7, 11, 14)). During gametogenesis, one of the genomes - the recombinant genome (in the case under discussion, genome R) - of such frogs is eliminated (destroyed). The other genome, the clonal genome (in this case, genome L), is doubled through endoreduplication. Thereafter, gametes are produced bearing identical copies of the parental L genome. This mode of inheritance is termed hemiclonal (semiclonal) (15). To designate the clonal genome, its symbol will be placed in parentheses. Individuals that have received two identical clonal genomes from their parents are not viable. In this case, we can establish that in the localities described, three types of crosses are possible: (1) RR x RR -> RR (2) (L)R x (L)R -> (L)(L) ->dagger (3) (L)R x RR -> (L)R The first and second types of cross do not affect the ratio of P. ridibundus to P. esculentus, while the third leads to an increase in the proportion of P. esculentus in the locality. It can be verified that during joint reproduction, representatives of the parental species and the water frog hybrids form distinctive systems in which both clonal and recombinant genomes are transmitted - hemiclonal population systems, hereafter HPS (11). HPS are characterised by transformations of their composition caused by internal and external factors; these transformations can be studied both through observation of natural systems and through the application of simulation modelling (8, 9). Levels of selection in P. esculentus complex. In light of the above, it is possible to determine at which levels selection operates to condition the features of HPS transformations in P. esculentus complex. In the examined example of population systems in the city of Kharkiv, significant transformations of HPS do not occur. This is most likely attributable to the fact that under conditions of a large city, representatives of P. ridibundus possess higher viability than P. esculentus. But this is not always the case. What occurs when the viability of representatives of the parental species and the hybrid frogs is equal can be determined through simulation modelling (9). In the example shown in Fig. 1, the genotype designations also indicate the sex-chromosome affiliation of the genomes. It should be noted that the sex-determination system in water frogs is analogous to that in humans: the male sex is heterogametic, the female is homogametic. [IMG_1] Fig. 1. Transformations of HPS in P. esculentus complex according to simulation modelling data (9). The entry of a single P. esculentus individual bearing a female clonal genome - (XL)XR - into a P. ridibundus population leads to a situation in which all individuals in the HPS carry the clonal genome, reproduction of the HPS ceases, and the HPS perishes. The hemiclonal inheritance of a genome (in the case shown in Fig. 1, genome XL) may be regarded as the limiting case of meiotic drive, in which not a single allele but an entire genome passes into the gametes. The state into which the HPS shown in Fig. 1 has entered is detrimental from the perspective of both SG and SI. The possibility of precisely such a course of events not only follows from the results of modelling but is also, to some degree, supported by the results of field observations. However, the transmission of clonal genomes does not invariably lead to disruptions in HPS reproduction. Figure 2 shows what will occur if clonal genomes belonging to different parental species are transmitted within an HPS. In this case, the HPS may enter a stable state. We have grounds to suppose that those HPS that find themselves in an unstable state change or disappear, while those in which a stable set of genomes has formed spread. In this case, these processes constitute an example of group selection (incidentally, one not associated with altruism). The most important argument advanced by R. Dawkins (6) against group selection is that a group lacks a genotype and cannot transmit its properties by inheritance. In the case under consideration, this argument does not apply. The analogue of the individual's genotype at the group level is its gene pool. The most important characteristic of the HPS gene pool is the set of clonal genomes that are transmitted within it. The "reproduction" of groups occurs during the dispersal of juveniles, in the course of which the daughter group may receive a relatively representative sample of clonal genomes from the gene pool of the parental group. [IMG_2] Fig. 2. Transformations of HPS in P. esculentus complex according to simulation modelling data (9). An HPS in which different clonal genomes are transmitted enters a stable state and reproduces normally. The effectiveness of group selection relative to genic and individual selection is reduced, since the transmission of a representative sample from the gene pool is a stochastic and less stable process than the transmission of genes during reproduction. However, such selection also has its strengths: for example, within it, "acquired" traits may be inherited (clonal genomes maintained by selection that entered the HPS or arose through the evolution of previously existing genomes). From the foregoing it becomes clear that the transformations of HPS in P. esculentus complex constitute a complex process upon which selection may act at different levels. In certain cases (by no means fully described in this paper), selection at different levels may prove to be oppositely directed. This renders the hybridogenetic complex of water frogs an excellent model for the study of multilevel selection. Conclusions 1. The concepts of the "selfish gene", the "selfish individual", and group selection are not alternatives. They are mutually complementary (in the sense of Niels Bohr). 2. For the separate study of selection at a given level, model systems are required in which selection at different levels is oppositely directed. For example, if selection of alleles is opposed to selection of individuals and groups, and observation demonstrates that changes proceed in the direction determined by allele selection, such a situation is optimal for the study of allele selection. 3. The hemiclonal inheritance of genomes in Pelophylax esculentus may serve as an example of the "selfish gene" in action, while the transformations of unstable hemiclonal population systems of water frogs may serve as an example of group selection. Overall, the hybridogenetic complex of water frogs affords broad opportunities for the study of multilevel selection. Acknowledgements The arguments presented in this paper are the outcome of the author's collaborative work on the study of water frogs with numerous colleagues. Of particular importance to the author was his collaboration with M. O. Kravchenko, O. V. Korshunov, M. V. Vladimirova, G. A. Lada, S. M. Litvinchuk, L. Ya. Borkin, and S. Yu. Morozov-Leonov. The author extends sincere gratitude to all named and unnamed colleagues. References 1. Bohr N. Atomic Physics and Human Knowledge / Niels Bohr. - Moscow: Inostr. lit., 1961. - 152 p. 2. Grant V. The Evolutionary Process: A Critical Review of Evolutionary Theory / Verne Grant. - Moscow: Mir, 1991. - 488 p. 3. Darwin Ch. The Descent of Man, and Selection in Relation to Sex. Works / Charles Darwin. - Moscow: Publishing House of the USSR Academy of Sciences, 1953. - Vol. 5. - 1040 p. 4. Darwin Ch. On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life. Translated from the sixth edition (London, 1872) / Charles Darwin. - St. Petersburg: "Nauka", 1991. - 539 p. 5. Dawkins R. The Selfish Gene / Richard Dawkins. - Moscow: "Mir", 1993. - 317 p. 6. Dawkins R. The Extended Phenotype: The Long Reach of the Gene / Richard Dawkins. - Moscow: Astrel: CORPUS, 2010. - 510 p. 7. Study of population systems of water frogs (Rana esculenta complex) in Kharkiv Oblast: history, current status and prospects / D. A. Shabanov, A. I. Zinenko, A. V. Korshunov (et al.) // Bulletin of V. N. Karazin Kharkiv National University. Series: Biology. - 2006. - Issue 3 (No. 729). - P. 208-220. 8. Investigation of the stability of hemiclonal population systems of the hybridogenetic complex of water frogs using simulation modelling / M. A. Kravchenko, D. A. Shabanov, M. V. Vladimirova (et al.) // Bulletin of Dnipropetrovsk University. Biology. Ecology. - 2011. - Issue 19, vol. 1. - P. 51-64. 9. Kravchenko M. A. Modelling of transformations of hemiclonal population systems of water frogs (Pelophylax esculentus complex; Amphibia, Ranidae) using recurrent difference equations / M. A. Kravchenko, D. A. Shabanov // Bulletin of V. N. Karazin Kharkiv National University. Series: Biology. - 2010. - Issue 12 (No. 920). - P. 70-82. 10. Ridley M. The Red Queen: Sex and the Evolution of Human Nature / Matt Ridley. - Moscow: Eksmo, 2011. - 448 p. 11. Shabanov D. A. Water frogs: life without rules or a special mode of evolution? / D. A. Shabanov, S. N. Litvinchuk // Priroda. - 2010. - No. 3 (1135). - P. 29-36. 12. Dawkins R. The descent of Edward Wilson. Prospect, 24 May 2012 (Online). 13. Hamilton W. D. The genetical evolution of social behavior (I and II) / Hamilton W. D. // Journal of Theoretical Biology. - 1964. - V. 7 - P. 1-16; 17-52. 14. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine / L. J. Borkin, A. V. Korshunov, G. A. Lada (et al.) // Russian Journal of Herpetology. - 2004. - Vol. 11, No 3. - P. 194-213. 15. Plotner J. Die westpalaearktischen Wasserfrosche / Jorg Plotner. - Bielefeld: Laurenti-Verlag, 2005. - 161 p. 16. Williams G. C. Adaptation and natural selection: a critique of some current evolutionary thought / George Christopher Williams. - Princeton, 1966. - 307 p. 17. Wilson E. The Social Conquest of Earth / Edward Wilson. - N.Y.: Liveright Publishing Corporation, 2012. - 352 p. 18. Wynne-Edwards V.C. Animal Dispersion in Relation to Social Behaviour / Vero Corner Wynne-Edwards. - Edinburgh: Oliver & Boyd, 1962. - 653 p. Article received by the editorial board 15.10.2012; accepted for publication 22.10.2012.