
Shabanov, Biriuk, Korshunov, Kravchenko (2017) Distribution of Various Types of Population Systems of Water Frogs in the Siverskyi Donets Basin
Shabanov D. A., Biriuk O. V., Korshunov O. V., Kravchenko M. O. Distribution of Various Types of Hemiclonal Population Systems of the Hybridogenetic Complex of Water Frogs (Pelophylax esculentus complex) in the Siverskyi Donets Basin // Current State and Protection of Natural Complexes...
Shabanov Dmytro Andriiovych1, Biriuk Olha Viktorivna1, Korshunov Oleksii Vladyslavovych1, Kravchenko Maryna Oleksandrivna1,2 1V. N. Karazin Kharkiv National University, 61022, Ukraine, Kharkiv, Svobody Square, 4; shabanov@karazin.ua; 2H. S. Skovoroda Kharkiv National Pedagogical University DISTRIBUTION OF VARIOUS TYPES OF HEMICLONAL POPULATION SYSTEMS OF THE HYBRIDOGENETIC COMPLEX OF WATER FROGS (PELOPHYLAX ESCULENTUS COMPLEX) IN THE SIVERSKYI DONETS BASIN Water frogs are a group of animals that deserve the exceptional attention of researchers and conservationists. This is related not only to their relatively high biomass and powerful influence on biogeocoenotic processes. It is precisely on the example of water frogs that one can observe insufficiently studied mechanisms for overcoming the sterility of interspecific hybrids, interspecific recombination, non-Mendelian inheritance, and the existence of unusual types of biosystems, etc. The Central European hybridogenetic complex of water frogs, Pelophylax esculentus complex, is formed by two parental species, the pool frog, Pelophylax lessonae (Camerano, 1882) and the marsh frog, Pelophylax ridibundus (Pallas, 1771), as well as their various hybrids, to which a species-like name is applied: the edible frog, Pelophylax esculentus (Linnaeus, 1758). Representatives of the parental species hybridize readily, producing viable hybrids. However, the genomes of P. lessonae and P. ridibundus, although consisting of the same number of chromosomes (n=13), have acquired such differences that their normal conjugation during meiosis has become impossible. It is precisely this phenomenon that causes the hybrid sterility of many interspecific hybrids. In various plants and animals, two main pathways for overcoming such sterility are known: tetraploidization (as a result of which each chromosome acquires a homologous pair) or transition to clonal, i.e. non-recombinant reproduction (various forms of asexual reproduction, parthenogenesis, gynogenesis, etc.). The third pathway is the rarest: hemiclonal reproduction of hybrids. In the typical case, in the germ line cells of P. esculentus, elimination of one of the parental genomes and endoreduplication (doubling without subsequent division) of the other parental, clonal genome occurs (Plötner, 2005; Dedukh et al., 2015). In the typical case, reproduction of P. esculentus involves hybridization of hybrids with representatives of that parental species whose genome they eliminate. We will explain what was stated in the previous paragraph, using the letter L to denote the genome of P. lessonae, and R for the genome of P. ridibundus; clonality is denoted by enclosing the genome symbol in parentheses. Hybrids arise as a result of hybridization of representatives of parental species (gametes are shown in rectangular cells: . In hybrids, gametes carrying the clonal genome are formed: . Upon crossing of hybrids with a representative of the parental species whose genome is eliminated, all offspring consist of hybrids: . Hybrid reproduction can thus proceed over many generations. Over most of the range, P. esculentus most frequently transmits R genomes clonally (Plötner, 2005). Over part of the P. esculentus range, both diploid and triploid hybrids occur, which can exist in two genomic compositions: LLR and LRR. The following variant of triploid gametogenesis can be considered typical: triploids transmit to gametes the genome represented by two copies: (Vinogradov et al., 1990). From the above it is clear that reproduction of P. esculentus takes place not in populations of any single species, but in special systems in which representatives of parental species and various hybrids jointly participate in reproduction, and in which, from generation to generation, both recombinant and clonal genomes are transmitted. These biosystems are termed hemiclonal population systems, HPS (Shabanov et al., 2009). We use a notation for HPS types in which letters denote the frog forms comprising them: L and R for parental species, and E for P. esculentus.
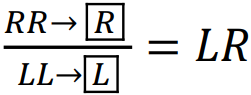
The marker p corresponds to the presence of polyploids (triploids) in the HPS, while markers f or m indicate the case where a given form is represented only by females or males, respectively.
If a given form is represented by only one genomic composition, this can be denoted by the corresponding subscript.
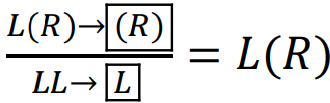
Examples of the described notation: for example, R‑E‑Ep‑HPS are characteristic of the Homilsha Forests NNP, while R‑LLREpf‑HPS occur in the Svyati Hory NNP. The examples cited concern the Siverskyi Donets basin. The study of water frogs in this region has a long history, but unfortunately in many cases it is impossible to determine which forms of frogs are discussed in works carried out prior to the development of reliable identification methods. The application of flow DNA cytometry, which allows distinguishing frog forms by genome size, led to the discovery in the Siverskyi Donets basin of a region of triploid distribution (Borkin et al., 2004).

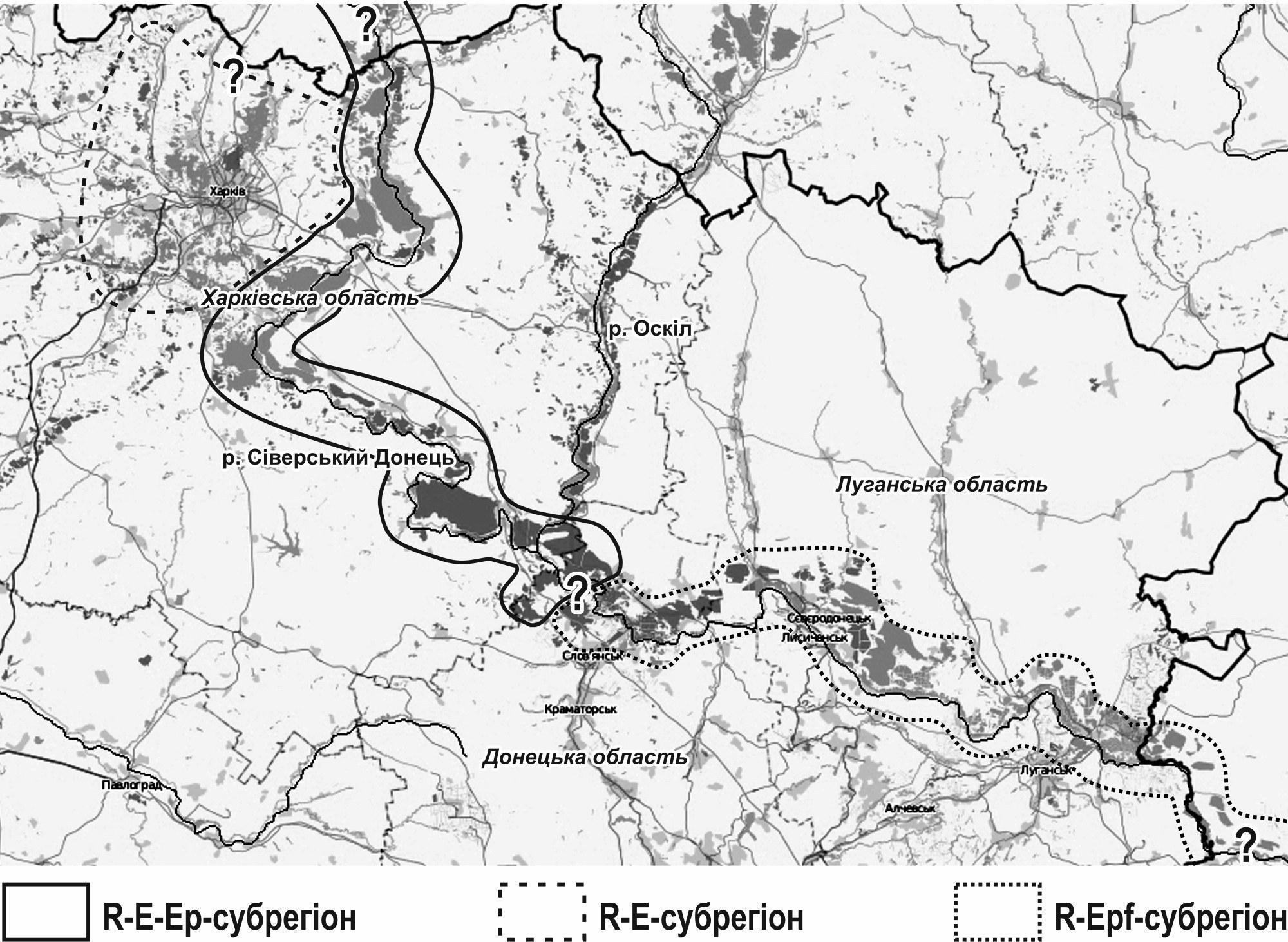
Shortly thereafter it was shown that the diversity of HPS composition in this region is linked to the diversity of gametogenesis of P. esculentus occurring there. Among diploid hybrids there are those that transmit the genomes of one or the other parental species, i.e. L(R) and (L)R, as well as those that produce a mixture of germ cells with genomes of both parental species: (L)(R) (Borkin et al., 2005). The complexity of the HPS described in the Siverskyi Donets is unprecedented. For example, on the territory of the Homilsha Forests NNP the following occur: — representatives of one of the parental species: ♀♀ and ♂♂ RR; — diploid hybrids: ♀♀ and ♂♂ L(R), (L)R, and (L)(R); — triploid hybrids: ♀♀ and ♂♂ LLR and LRR; — rare immature tetraploids LLRR and representatives of the other parental species, LL. Paradoxically, sexually mature P. lessonae have not been found in the Siverskyi Donets basin (Korshunov, 2010); all L genomes are transmitted here through hybrids. Over time, this should lead to degradation of L genomes through the action of “Müller's ratchet,” a consequence of the cessation of genetic recombination. Presumably, this does not occur owing to recombination of L genomes during gametogenesis in LLR hybrids (in accordance with the scheme presented above). The circumstances described above led to the description of the Siverskyi Donets diversity centre of Pelophylax esculentus complex (Shabanov et al., 2009). This centre consists of parts that differ from one another. Two descriptions of the geographical structure of this centre have been made. The first of these descriptions is based on an analysis of the HPS composition of water frogs of the Left-Bank Forest-Steppe of Ukraine (Shabanov, 2015). In this region, 5 subregions are distinguished. — Subregion I is inhabited by ordinary populations of P. ridibundus. It comprises portions of the Siverskyi Donets basin where there are no habitats meeting the biogeocoenotic requirements of P. lessonae and P. esculentus, as well as the Oskil basin, a left tributary of the Siverskyi Donets. — Subregion II is located in the Dnipro basin, in that part where there are water bodies suitable for P. esculentus and P. lessonae situated adjacent to forested areas. Another part of the Left-Bank Forest-Steppe in the Dnipro basin belongs to Subregion I. The most complex HPS in Subregion II are R‑E‑L‑HPS; R‑E‑HPS, L‑E‑HPS and populations of P. ridibundus also occur. It includes an isolated (presumably relict) enclave with R‑E‑L‑HPS in the Rusky Orchyk nature reserve on the boundary of Kharkiv, Poltava, and Dnipropetrovsk oblasts. — Subregion III is located in the basins of the Mzha and Udy rivers, right tributaries of the Siverskyi Donets, as well as the Lopan River (tributary of the Udy) and the Kharkiv River (tributary of the Lopan). R‑E‑HPS and populations of P. ridibundus are distributed here. — Subregion IV is the most diverse in terms of the number of frog forms registered there (listed above for the Homilsha Forests NNP). It extends along the Siverskyi Donets to Donetsk Oblast. The most complex systems are R‑E‑Ep‑HPS. In certain HPS some frog forms are absent; for example, in the pond of the village Zamulivka (formerly Zhovtneve), Vovchansk district, an R‑E‑Epf‑HPS occurs where triploid hybrids are represented only by females. — Subregion V is located below Subregion IV along the Siverskyi Donets, on the territory of Donetsk and Luhansk oblasts and, presumably, Rostov Oblast of the Russian Federation. R‑LLREpf‑HPS are characteristic of it. The objects of study by the authors of this work were HPS near the village of Brusivka in Donetsk Oblast and the city of Kreminna in Luhansk Oblast, but other similar systems also exist in this region (Shabanov, 2015). On the territory of Subregion V there remain areas corresponding in HPS composition to those of Subregion IV; for example, an R‑E‑Ep‑HPS (presumably relict) has been recorded near the village of Yasnohirka near Kramatorsk. The second description of the geographical structure of the Siverskyi Donets diversity centre of water frogs is associated with the study of gametogenesis and mating systems in HPS (Biriuk et al., 2016; Biriuk, 2017). Based on these characteristics, three groups of HPS are distinguished. — HPS of Group I consist of P. ridibundus and diploid P. esculentus. They correspond to Subregion III in the previous classification (Shabanov, 2015). — HPS of Group II consist of P. ridibundus and triploid P. esculentus, and correspond to Subregion V. — HPS of Group III are geographically located between the two preceding groups, consist of P. ridibundus, di- and triploid P. esculentus, and correspond to Subregion IV in the previous classification. The most interesting is the mating system described for Group II HPS, i.e. R‑LLREpf‑HPS. Females with genotype LLR typically produce haploid egg cells L (Dedukh et al., 2015). They mate with males of P. ridibundus, which produce spermatozoa with genotype R. It has been suggested (Biriuk et al., 2015) that triploid zygotes arise as a result of doubling of the L genome occurring after fertilization. Other explanations of the registered phenomenon are also possible. Among them (besides, naturally, the presence of unregistered P. esculentus in these HPS that produce diploid gametes) are blocking of the last meiotic division and polar body separation, or fusion of the egg cell with the polar body after fertilization. In describing the subregions of the Left-Bank Forest-Steppe of Ukraine by HPS composition of water frogs, it was noted that the Siverskyi Donets diversity centre of Pelophylax esculentus complex includes Subregions III and IV (Shabanov, 2015). A diversity centre is a territory in which an elevated, compared to other territories, number of forms of the group under consideration is registered (Shabanov et al., 2009). Currently, in our view, it can be considered that the unusual reproduction mode of triploids in the HPS of Subregion V provides grounds for including this subregion in the Siverskyi Donets diversity centre as well, since here the reproduction of water frogs takes place by a greater number of modes than elsewhere. The inclusion of Subregion III in this centre is justified, in our view, by its proximity (both geographical and in terms of diploid P. esculentus composition) to Subregion IV (Shabanov, 2015) and its sharp distinction from the water frog HPS inhabiting the Dnipro basin (Mezhzherin et al., 2010). We propose to combine the two descriptions listed above and distinguish three subregions within the Siverskyi Donets diversity centre of Pelophylax esculentus complex (Fig. 1). The origin of the entire diversity centre is presumably related to the R‑E‑Ep‑subregion. The designation of this (and other) subregions indicates not what HPS inhabit their entire territory, but what HPS inhabit the more favourable habitats. The frog forms distributed here could have reached the territory of Ukraine from Belgorod Oblast of the Russian Federation, descending along the Siverskyi Donets. The basis for this assumption is that E‑HPS were registered in Belgorod Oblast (Lada, 1998), which were also described in the R‑E‑Ep‑subregion. Unfortunately, the E‑HPS in the R‑E‑Ep‑subregion (HPS of Iskiv Pond and Koriakov Ravine) have transitioned to another state; the study of their transformations is an interesting task (Makaryan et al., 2016). An important problem in the study of this subregion is the determination of mechanisms preserving a significant number of L genomes and maintaining a relatively constant proportion of triploid hybrids in R‑E‑Ep‑HPS. The R‑E‑subregion is located in the basins of the Mzha and Udy rivers. The composition of its population systems has been studied, among other methods, by simulation modelling (Kravchenko, Shabanov, 2010; Shabanov, 2015). It has been shown that the HPS characteristic of this subregion can maintain stability owing to differences in the intra-population ontogenetic strategies of the frogs composing them (Shabanov et al., 2014; Usova et al., 2015; Shabanov et al., 2015). Fig. 1. Subregions of the Siverskyi Donets diversity centre of Pelophylax esculentus complex, the hybridogenetic complex of water frogs The R‑Epf‑subregion is located on the territory of Donetsk and Luhansk oblasts. It harbours not only R‑LLREpf‑HPS but also R‑E‑Epf‑HPS and, naturally, populations of P. ridibundus. It can be assumed that the R‑Epf‑subregion extends beyond the borders of Ukraine onto the territory of the Russian Federation, since triploid LLR females were also found in Rostov Oblast (Borkin et al., 2006). It is clear that R‑LLREpf‑HPS are distributed not across the entire territory shown in Fig. 1, but only in suitable habitats. It should be emphasised that identification of triploid female P. esculentus by external characteristics is unreliable (Korshunov, 2010); besides high-technology methods, erythrocyte size determination can be used to confirm the diagnosis of triploids (Bondareva et al., 2012). It is indisputable that establishing the reproduction mechanisms of triploid hybrids in this region is an urgent scientific problem. The R‑Epf‑HPS themselves are unique and insufficiently studied natural biosystems that require careful conservation. According to our data (Korshunov, 2010), the most important condition for the preservation of the unusual water frog HPS is the maintenance of the biogeocoenoses characteristic of them, which include the water bodies where frogs reproduce and the forested areas situated adjacent to them. Biriuk O. V. Cytogenetic features of spermatogenesis in diploid and triploid hemiclonal interspecific hybrids of Pelophylax esculentus complex: abstract of thesis for Cand. Biol. Sci. degree, speciality 03.00.15 – genetics. – Kyiv, 2017. – 22 p. Bondareva A. A., Bybyk Yu. S., Samylo S. M., Shabanov D. A. Cytogenetic features of erythrocytes of water frogs from the Siverskyi Donets diversity centre of Pelophylax esculentus complex // Visnyk KhNU im.
V. N. Karazina. Ser.: biol. – 2012. –15 (1008) – P. 116–123. Borkin L. Ya., Zinenko A. I., Korshunov A. V. et al. Mass polyploidy in the hybridogenetic complex Rana esculenta (Ranidae, Anura, Amphibia) in Eastern Ukraine: Proc. First Conf. of the Ukrainian Herpetological Society – Kyiv: Zoomuzei NNPM NANU, 2005. – P. 23–26. Korshunov O. V. Ecological regularities of distribution of Pelophylax esculentus complex in habitats of the upper Siverskyi Donets River basin: abstract of thesis for Cand. Biol. Sci. degree, speciality 03.00.16 – ecology. – Dnipropetrovsk, 2010. – 20 p. Kravchenko M. A., Shabanov D. A. Modelling of transformations of hemiclonal population systems of water frogs (Pelophylax esculentus complex; Amphibia, Ranidae) using recurrent difference equations // Visnyk Kharkivskoho natsionalnoho universytetu imeni V. N. Karazina. Seriia: biolohiia. – 2010. – Iss.12 (No.920). – P. 70–82. Lada, G. A. On the necessity of preserving unique “pure” populations of the diploid edible frog (Rana esculenta Linnaeus, 1758) in Belgorod and Kharkiv oblasts: Problems of protection and rational use of natural ecosystems and biological resources. – Penza, 1998. – P. 333–335. Makaryan R. M., Biriuk O. V., Korshunov O. V. et al. Composition of water frog tadpoles (Pelophylax esculentus complex) in Iskiv Pond (Homilsha Forests NNP): State and biodiversity of ecosystems of the Shatsk NNP and other protected areas. – Lviv: SPOLOM, 2016. – P. 61-65. Usova E. E., Kravchenko M. A., Shabanov D. A. Intra-population ontogenetic strategies in water frogs (Pelophylax esculentus complex) // Visnyk Kharkivskoho nats. un-tu im. V. N. Karazina. Seriia: biolohiia. – 2015. – Iss. 25. – P. 223-238. Shabanov D. A. Evolutionary ecology of population systems of the hybridogenetic complex of water frogs (Pelophylax esculentus complex) of the Left-Bank Forest-Steppe of Ukraine: abstract of thesis for Dr. Biol. Sci. degree, speciality 03.00.16 – ecology. – Dnipropetrovsk, 2015. – 36 p. Shabanov D. A., Korshunov A. V., Kravchenko M. A. et al. Intra-population ontogenetic strategies of early and delayed maturation: definition illustrated by anuran amphibians // Vestnik Kharkovskogo natsionalnogo universiteta imeni V. N. Karazina, seriia “Biologiia”. – 2014. – Iss.22, No.1126. – P. 115-124. Shabanov D. A., Korshunov O. V., Kravchenko M. O. Which water frogs inhabit Kharkiv Oblast? Terminological and nomenclatural aspects of the problem // Biolohiia ta valeolohiia. – Iss. 11. – Kharkiv: KhDPU, 2009. – P. 116–125. Biriuk O., Shabanov D., Korshunov O. et al. Gamete production patterns and mating systems in water frogs (hybridogenetic Pelophylax esculentus complex) in North-Western Ukraine // J. of Zoology Systematics and Evolutionary Res. – 2016. – 54 (3). – P. 215–225. DOI: 10.1111/jzs.12132. Borkin L. J., Korshunov A. V., Lada G. A. et al. Mass occurrence of polyploid green frogs (Rana esculenta complex) in Eastern Ukraine // Russian J. of Herpetology. – 2004. – 11 (3). – P. 194–213. Borkin L. J., Lada G. A., Litvinchuk S. N. et al. The first record of mass triploidy in hybridogenic green frog Rana esculenta in Russia (Rostov oblast') // Russian Journal of Herpetology. – 2006. –13 (1). – P. 77–82. Dedukh D., Litvinchuk S., Rosanov J. et al. Optional endoreplication and selective elimination of parental genomes during oogenesis in diploid and triploid hybrid european water frogs // PLoS ONE. – 2015. – 10(4): e0123304. doi:10.1371/journal.pone.0123304. Mezhzherin S. V., Morozov-Leonov S. Yu., Rostovskaya O. V. et al. The ploidy and genetic structure of hybrid population of water frogs Pelophylax esculentus complex (Amphibia, Ranidae) of Ukraine fauna // Cytology and Genetics. – 2010. – Vol. 44, No 4. – P. 212–216. Plötner J. Die westpaläarktischen Wasserfösche. – Bielefeld: Laurenti-Verlag, 2005. – 161 S. Shabanov D., Usova O., Kravchenko M. et al. Sustainable coexistence of the parental species and hemiclonal interspecific hybrids is provided by the variety of ontogenetic strategies // Herpetological Facts Journal – 2015. – 2. – P. 35–43. Vinogradov A. E., Borkin L. J., Günther R., Rosanov J. M. Genome elimination in diploid and triploid Rana esculenta males: cytological evidence from DNA flow cytometry // Genome. – 1990. – Vol. 33, No. 5. – P. 619–627. Shabanov D. A., Biriuk O. V., Korshunov O. V., Kravchenko M. O. Distribution of various types of hemiclonal population systems of the hybridogenetic complex of water frogs (Pelophylax esculentus complex) in the Siverskyi Donets basin // Current State and Protection of Natural Complexes in the Siverskyi Donets Basin. Proc. scientific-practical conf. dedicated to the 20th anniversary of Svyati Hory NNP. – Sviatohirsk, TOV “Vydavnytstvo ‘Drukarski dvir’”, 2017. – P. 139-144.