The wonders of sexual reproduction. Column in ComputerreOnline #48
The mechanisms of gene transmission and alteration differ among asexual species, the parasexual Penicillium, humans and other animals with a typical sexual cycle, and the truly extraordinary Pushtun frogs
Discussion of the latest "evolutionary" columns has drawn attention to the topic of the evolution of evolution. Some readers find it hard to believe that different groups of organisms evolve (develop adaptations) in different ways. I will give examples from my own work.
Among the evolutionary acquisitions that accelerate evolution, the largest seem to be sexual reproduction and cultural inheritance. We have talked about cultural inheritance. The successes of our species are testimony to the power of this mode of generating adaptations. A much older invention is sex, more precisely the haplo‑diploid life cycle with meiosis. I will now explain what this means.
Exchange of genetic information accelerates the evolution of sexually reproducing organisms enormously compared with asexual species (perhaps I will someday build a model and substantiate this claim with it). But obtaining genetic information "from the outside" requires a special life cycle. Imagine: a cell with a certain set of chromosomes (molecular complexes carrying genetic information) repeatedly receives new ones. How to reduce the excess chromosomal load?
The simplest solution is achieved by a parasexual process. It is characteristic of many fungi, such as Penicillium or Aspergillus – the molds in our kitchens. Hyphae (thread‑like fungal bodies) of different genetic individuals come into contact and exchange nuclei. Different nuclei can fuse, forming hybrid nuclei with a double set of chromosomes. These nuclei, like the others, divide by mitosis. Mitosis is the most common division of nuclei and cells; our bodies are also formed by this division method.
During mitoses of hybrid nuclei in fungi, step by step, elimination (removal) of excess chromosomes occurs. Sometimes recombination is observed – exchange of segments between paired chromosomes, creating new combinations of genetic material. The different nuclei that result after simplification of the hybrid nucleus have different chromosomal parts by origin.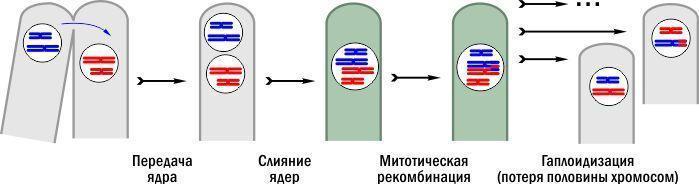
Scheme of the parasexual process typical for many fungi
In the illustration, hyphae where nuclei have a haploid (single) chromosome set are shown in gray, and diploid (with two chromosome sets) are colored greenish. However, life is more complex than the scheme. Diploid fungal nuclei become haploid gradually, after many mitoses, and the same hyphae can contain different nuclei.
Greater complexity is achieved by organisms that use a special type of division – meiosis – to reduce the chromosome set. A comparison of mitosis and meiosis is shown in the diagram (do not be alarmed: this is standard school material!). Meiosis consists of two successive divisions. In the first, chromosomes separate between daughter cells; in the second, as in mitosis, chromosomes split into chromatids. The result is four genetically unique cells.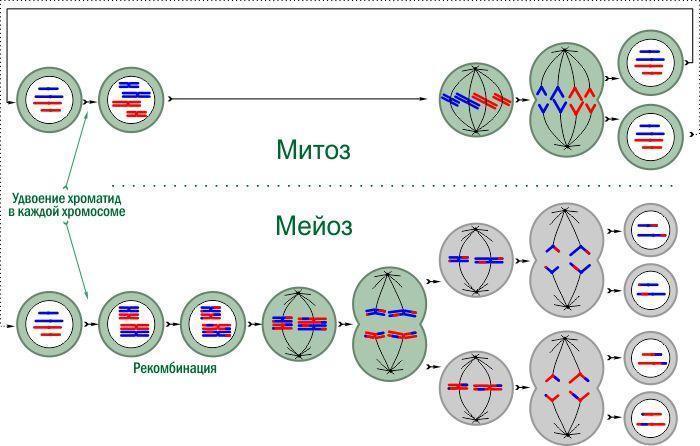
Comparison of mitosis and meiosis. Mitosis – a cycle; some of the cells formed in it may proceed to meiosis. Chromosomes may consist of a single chromatid or, after duplication of genetic material, of two. Haploid cells are gray, diploid – greenish.
Thanks to meiosis, fertilization can become a permanent part of the life cycle: the fusion of gametes (sex cells) forms a zygote. Fertilization doubles the chromosome set, while meiosis reduces it.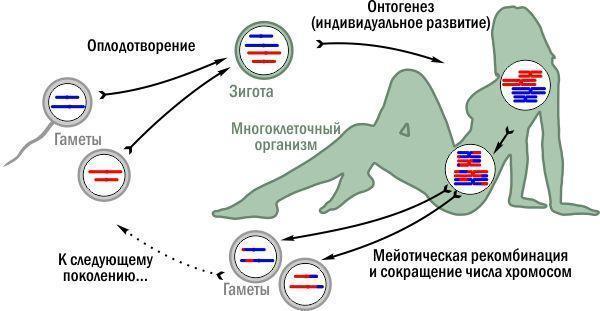
Typical haplo‑diploid life cycle with meiosis. Haploid stages are gray, diploid – greenish.
The diagram shows a life cycle in which a multicellular organism develops in the diploid phase. This is not the only possible solution. In some archaic species, multicellular stages are haploid, and meiosis occurs immediately after fertilization. In plants, cycles are typical where multicellular organisms arise in both diploid and haploid phases. In any case, the haplo‑diploid life cycle with meiosis (now you understand what this means) is the evolutionary mainstream. Yet after its emergence, experiments with reproductive modes did not cease.
I will start with an example close to me. I study hybridisation of green frogs (details here). Without going into details, two different species of green frogs interbreed, producing hybrids (Pelophylax esculentus). In diploid hybrids, before meiosis, one parental chromosome set is eliminated in the cells that will become gametes. This is somewhat analogous to chromosome elimination in the parasexual process of fungi, but in fungi any chromosome in each pair may be removed, whereas frogs lose an entire set.
Then, in the cells that have become haploid, the chromosome set is duplicated by endomitosis ("internal" mitosis, chromosome duplication without division). This yields a diploid cell with two identical chromosome sets. Recombination occurs between them, which in the typical case changes nothing: identical chromosomes exchange identical segments. The result are gametes carrying the chromosome set of one parental species unchanged, clonally. Because only one genome is transmitted clonally, this reproduction is called semi‑clonal (hemiclonal). Besides green frogs, it has been found in some other animal groups.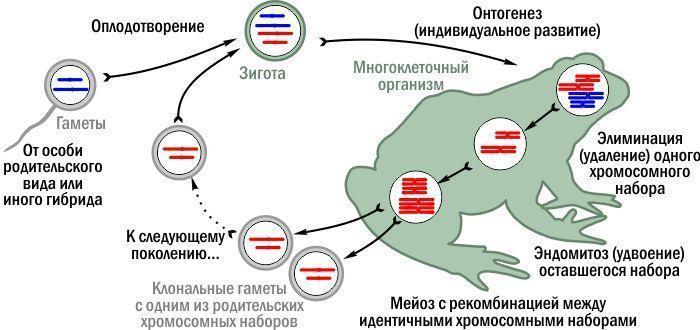
Semi‑clonal inheritance in interspecific hybrids of green frogs. Chromosome colour indicates their origin from different species.
Note: in semi‑clonal green frogs the mechanism of transmitting hereditary information between generations changes. The consequence is a special way of their evolution!
Now it is time to report that in some regions where hybrid frogs live (for example, where I work) there are not only diploid individuals but also triploid ones – with three chromosome sets.
Not long ago it was taken as an axiom that triploid animals are incapable of sexual reproduction: their chromosomes cannot pair during meiosis. Yet triploid frogs have found tricks to bypass this limitation. Here I will describe the variant that can be considered typical.
Unlike the two previous figures, I will not draw the full reproductive cycle of triploids. It has been studied for Western Europe and differs across regions; we have not yet firmly established the mechanisms of triploid formation in our study area, but we know which gametes they most often produce.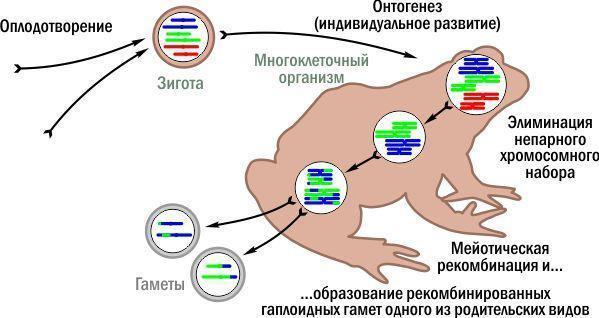
"Typical" gamete formation in triploid frogs. Red chromosomes belong to one parental species, blue and green to the other. Triploid stages appear reddish.
Simply by forgoing endomitosis (duplication of the remaining sets), triploid frogs can produce haploid gametes that are not clonal but recombined – as the parental species would produce. If endomitosis does occur, the gametes become diploid. If a diploid and a haploid gamete fuse at fertilisation, a new triploid arises.
Does the existence of triploid frogs seem like a curiosity to you? Do not rush. One of the advantages associated with sexual reproduction is precisely recombination, the continuous reshuffling of genes that allows the removal of deleterious mutations. Clonally transmitted chromosomes accumulate anomalies and gradually lose viability. This, for example, happens in diploid semi‑clonal frogs.
In our region, in the basin of the Seversky Donets River, one parental species of the hybrid frogs, Pelophylax lessonae, is absent. All its chromosome sets are transmitted through hybrids, but they do not degenerate. Apparently this is a consequence of recombination in triploids. Thanks to triploids, a new quality of hybrid evolution arises, allowing them to persist without the parental species!
My story has one final step.
At the end of the 20th century, German batrachologist Matthias Stöck began a series of extreme expeditions across Central Asia. He was there, where for a European to return alive is a great luck. In 1999 Stöck and his colleagues described the first triploid vertebrate species consisting of normally interbreeding males and females. These are the toads Bufo baturae, found in the arid highlands of Karakoram in Pakistan. Last year the results of thorough studies of the reproduction of these toads were published. If interested, an informal translation of Stöck and co‑authors' article is posted on my website.
Singing male of the Pushtun toad and a pair of toads spawning (see the egg strings?). Photo M. Stöck
A difficult question: how to name Bufo baturae in Russian? While writing this column I consulted the undisputed authority – Spartak Litvynchuk from St. Petersburg (thanks to him for the consultation!). He said that informally he and his colleagues call this species the Pushtun toad. Well, I will use this name as well – unofficially.
The Pushtun toad is of hybrid origin. From one parental species it inherited one chromosome set lacking a nucleolar organizer, from the other – two sets with nucleolar organizers. Without going into details, the nucleolar organizer is a region on certain chromosomes visible with specific staining. Let us assume that Pushtun toads have one "-" chromosome set and two "+" sets.
Gamete formation in females and males of Pushtun toads proceeds differently. Males implement the variant described above for triploid frogs: the "-" set is eliminated, and the two "+" sets undergo normal meiosis. The output are haploid sperm with one recombinant "+" set.
In females, before gametogenesis the "-" set is duplicated, producing cells with four sets: two "-" and two "+". Normal meiosis follows. In the "-" sets recombination occurs between identical copies and changes nothing, whereas in the "+" sets recombination generates new chromosome combinations. Females produce diploid eggs with one "-" set and one "+" set. Fertilisation by a "+" sperm restores the original genetic configuration.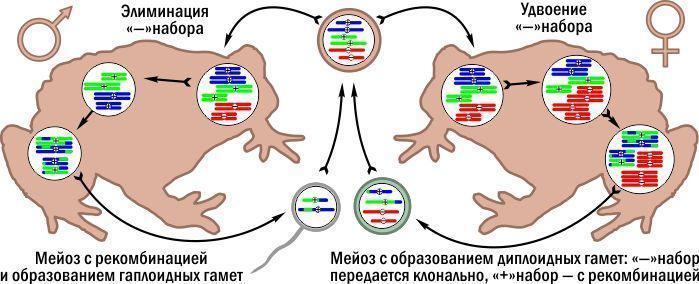
Reproduction of the Pushtun toad Bufo baturae. The "-" set (red) inherited from one parental species is transmitted clonally; the "+" sets from the other species are recombinant.
What amazes me most is that out of the 33 chromosomes in female Pushtun toads, only the 11 belonging to the "-" set are duplicated before meiosis. How can this be – 11 chromosomes are duplicated while the other 22 simply remain in the same nucleus? It is easier for me to believe that all chromosomes are duplicated, but then the extra copies of the "+" sets are eliminated. Either way, the Pushtun toad handles its genetic material differently from most inhabitants of this planet.
Stöck and co‑authors argue that the "-" set of the Pushtun toad should degenerate due to clonal transmission. They consider the species to have arisen recently and to be doomed to extinction. I doubt this forecast. Recall: hybrid green frogs found a way to "rejuvenate" their clonal sets in triploid individuals. I allow that something similar may occasionally happen in Pushtun toads.
It is evident to me that the features of evolution, modes of generating adaptations, mechanisms of gene transmission and alteration differ among asexual species, the parasexual Penicillium, humans and other animals with a typical sexual cycle, semi‑clonal green frogs and the truly extraordinary Pushtun toads. And do not think that the surprises one can encounter when studying reproductive modes of different species are exhausted!
By the way, the Pushtun toad is no longer alone. My colleagues from St. Petersburg found a similar species in the Pamir, and last year another one in the Western Trans‑Gimalaya. Let us wait to see what will be learned about the reproductive modes of these new finds.