V. Reptilia sensu lato-01. History of reptiles. Phylum Synapsida
Reptiles evolved through several evolutionary branches. The most diverse are the diapsids, but other groups also hold exclusive interest
It seems that the systematics of no other vertebrate group has undergone such changes or faced such difficulties as the systematics of reptiles. One of the authors of this textbook taught for many years a completely logical reptile systematics, which was built on morphological and paleontological data and seemed understandable. Then, thanks to molecular phylogenetics, biology received a huge influx of new data, and it became clear that we had understood reptile phylogeny far too inadequately.
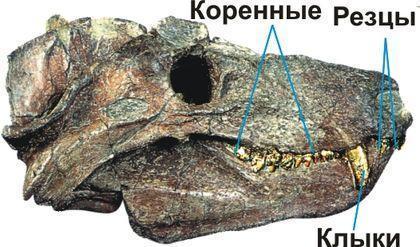
The problem that destroyed the whole system was the placement of turtles in the system. For many years the main criterion in the reptile section (in the broad sense) for grouping was the structure of the temporal regions of their skulls. The primitive condition – a solid, anapsid skull, with a temporal region formed by a continuous shield of covering bones, inherited from the pectoral‑finned fishes. This condition was characteristic of the ancestors of reptiles, their most primitive representatives, a separate group that received the name parareptiles, and, in a somewhat modified form, in modern turtles. Because of this, turtles were included among the parareptiles. In other groups, in order for the musculature that moves the jaws to have space to expand during contraction (and also simply to lighten the head), temporal openings – the temporal fenestrae – appeared in the skull. Three types of fenestration (opening formation) in the skull were distinguished. For the line that eventually gave rise to mammals, the formation of a single temporal opening is characteristic. Such a skull is called synapsid.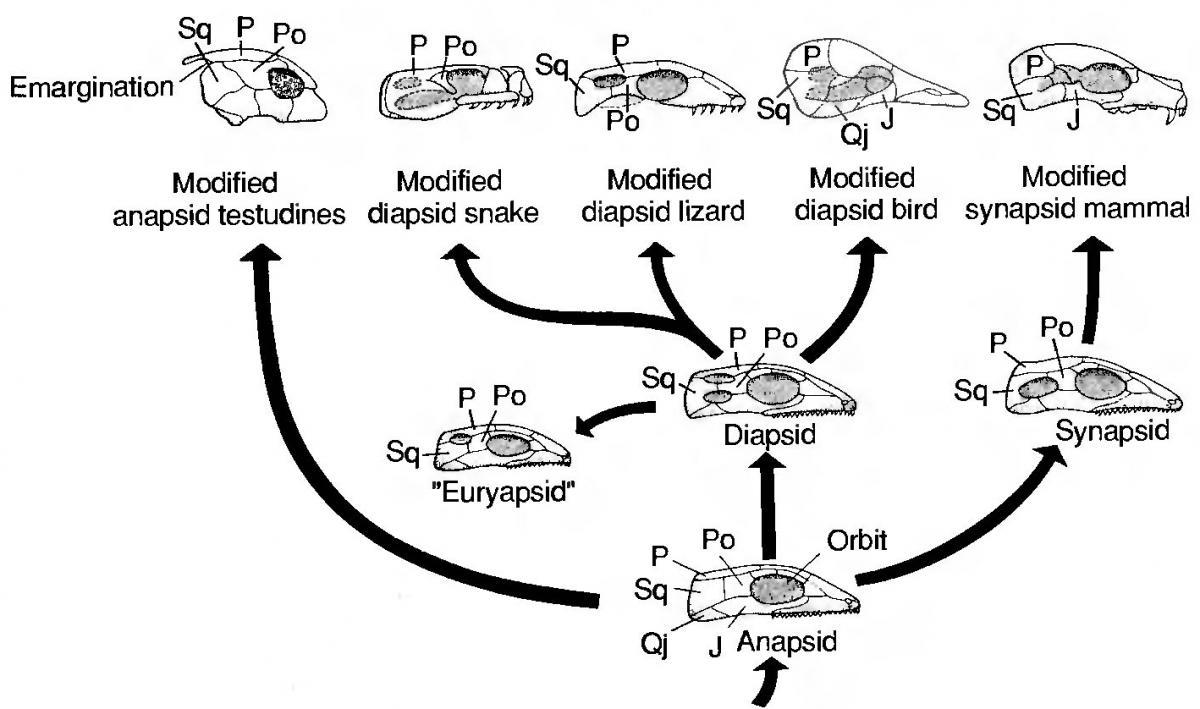
The great majority of reptiles belonged to groups in which two temporal windows formed (such a skull is called diapsid). Crocodylians and gatoriids retained the diapsid skull; lizards lost the lower ventral arch that closed the lower window, and snakes generally lost the arches that closed the windows. Birds, which also belong to the archosaur group characterized by a typical or modified diapsid skull, on the contrary, lost the arches that closed the temporal openings from above.
For two marine reptile groups, the ichthyosaurs and the synapsid‑like reptiles (plesiosaurs, together with other closely related groups) an euryapsid skull was characteristic. It had one opening, but not like the synapsid one, rather similar to the upper opening of a diapsid. On this basis it was assumed that euryapsids are a side branch of diapsids.
Based on the fenestration scheme of the temporal part of the skull described here, reptiles were divided into subclasses. And here the possibilities for molecular phylogenetics opened.
What relationships between turtles, gatoriids, lizards (and other lepidosaurs) and crocodylians could be expected on the basis of such views? Gatoriids, lepidosaurs and crocodylians are diapsids. They should have been relatively close. Turtles and mammals should have been phylogenetically much more distant from diapsids, and by a large margin.
Surprisingly, turtles ended up on a branch between different diapsids. Most likely they split from the common branch with crocodylians after the gatoriid and lepidosaur lines diverged… How can this be explained? In any case, the structure of the temporal region of the skull ceased to be a reliable guide.
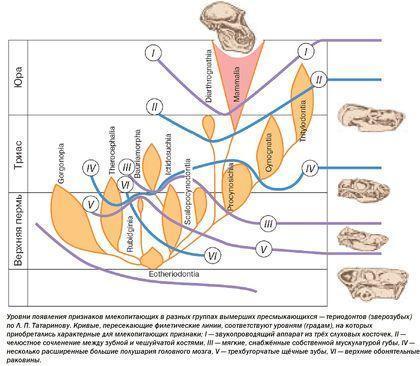
There is no universally accepted reptile systematics at present. For teaching purposes a very simplified provisional system is given below. In it turtles are combined (based on paleontological data) with synapsids. Another “problematic” group – the ichthyosaurs – is treated as a separate infraclass of eureptiles.
As in other cases, the groups that students are supposed to imagine are highlighted in brick colour.
Class “Reptilia” (sensu lato) – Reptiles (in the broad sense, including synapsids)
Phylum “Synapsida” – Synapsids
† Order Caseasauria – Caseasaurs
† Order Pelycosauria – Pelycosaurs
Order “Therapsida” – Therapsids
† Suborder Biarmosuchia – Biarmosuchians
† Suborder Dinocephalia – Dinocephalians
† Suborder Anomodontia – Anomodonts
† Suborder Gorgonopsia – Gorgonopsians
† Suborder Therocephalia – Therocephalians
Suborder “Cynodontia” – Cynodonts
Phylum Sauropsida – Sauropsids
† Subclass Parareptilia – Parareptiles
† Order Pareiasauromorpha – Pareiasauromorphs
Subclass “Eureptilia” – Eureptiles (True reptiles)
† Infraclass Ichthyosaurs – Ichthyosaurs
Infraclass Pantestudines – Pantestudines
† Order Placodontia – Placodonts
† Order Eosauropterygia – Eosauropterygians
† Order Plesiosauria – Plesiosaurs
Order Testudines – Turtles
Infraclass Lepidosauria – Lepidosaurs
Order Rhynchocephalia – Rhynchocephalians
Order Squamata – Lepidosaurs
Infraclass “Archosauria” – Archosaurs
Order Crocodylia – Crocodylians
† Order Pterosauria – Pterosaurs
† Suborder Rhamphorhynchoidea – Rhamphorhynchoids
† Suborder Pterodactyloidea – Pterodactyloids
Superorder “Dinosauria” – Dinosaurs
Order “Saurischia” – Saurischians
Suborder “Theropoda” – Theropods
† Suborder Sauropodomorpha – Sauropodomorphs
† Order Ornithischia – Ornithischians
† Suborder Thyreophora – Thyreophorans, armored dinosaurs
† Infraorder Stegosauria – Stegosaurs
† Infraorder Ankylosauria – Ankylosaurs
† Suborder Cerapoda – Cerapods
† Infraorder Pachycephalosauria – Pachycephalosaurs
† Infraorder Ceratopsia – Ceratopsians
† Infraorder Ornithopoda – Ornithopods
Note: the presented systematics is highly simplified! Most likely it lacks a considerable number of small groups that played a huge role in evolution and the origin of important clades.
In this systematics the symbol † marks groups whose all members and descendants are extinct. Brackets are used for groups that do not include all their descendants (i.e., they are paraphyletic). Thus, mammals are not included in “Synapsida”, and birds are not included in “Eureptilia”. Obviously, the group we label “Theropoda” (which is most likely ancestral to birds) in the sense we present here is extinct. However, some descendants of this group probably still exist.
Phylum “Synapsida” – Synapsids
We will consider three groups of synapsid animals. The first, the order Caseasauria – caseasaurs, is the most primitive. According to some views, they are not closely related to the other two groups; perhaps in this group the development of a single synapsid window proceeded differently than in the other groups. Representatives of this order are known from the Late Carboniferous and Permian. Undoubtedly, the caseasaur Cotylorhynchus, depicted in the next figure against a rear background, belongs here. These were Permian, fairly large herbivorous animals. Cotylorhynchus reached 6 m in length and was the largest known animal of its time. In front of Cotylorhynchus are Ophiacodon and Varanops – representatives of groups that are often placed in the next order, but sometimes are allied with caseasaurs.
Ophiacodon is a Late Carboniferous and Early Permian semi‑aquatic predator. The largest specimens of the genus exceeded 3 m in length. In some specimens the skull could be diapsid; in one individual the skull was diapsid on one side and synapsid on the other (!). Such asymmetry may indicate developmental instability of traits that later became much more stabilized in the group’s evolution. The family Varanopidae, to which Varanops belonged, consists of small (up to 1 m) Permian animals.
The order Pelycosauria is known from the Late Carboniferous and Early Permian. Carnivorous pelycosaurs belonged to the Sphenacodontia, and herbivorous ones to the Edaphosauria. Both groups are characterized by large crests formed by spine‑bearing processes of the vertebrae.
The best‑known sphenacodont is Dimetrodon. This is an Early Permian genus with several species.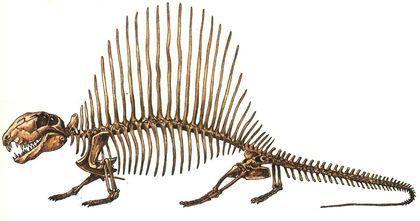

The largest dimetrodonts reached 4 m in length.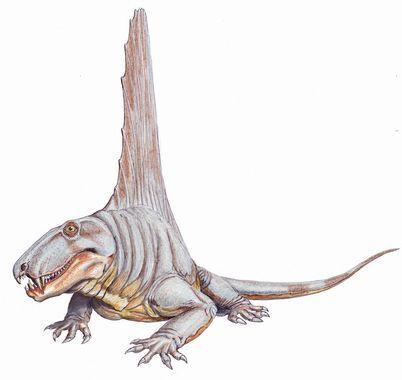
The largest edaphosaurids (Edaphosaurus) reached 3 m in length. Their crests also bore transverse processes that turned the whole “sail” into a complex three‑dimensional structure.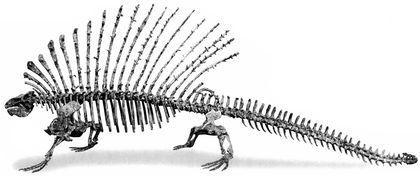
It is considered that sphenacodonts were the ancestors of later therapsids (and, consequently, also of us).
Dinomorphs (Dinomorpha) include fairly large Permian animals. Gorgodontia – carnivorous and herbivorous animals – are divided into two suborders: Dinocephalia (“terrible heads”) and Gorgonopsia. In both groups heterodonty developed – the division of teeth into functional groups. In many dinocephalians this division differed from that of mammals. This circumstance, together with the development of cranial thickening, gave them their name.
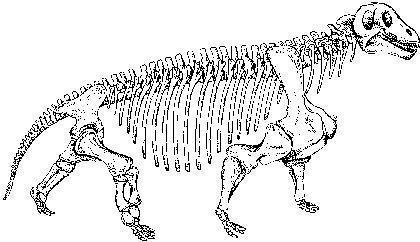
An example of a carnivorous dinocephalian is Titanophoneus (“titanic killer”), whose skeleton and skull are shown on the next two slides, and a reconstruction on the following one. This animal reached several metres in length.


Some dinocephalians were herbivorous. For example, this applies to the Tapinocephalidae. These large animals processed plant material with their front teeth, pushing it forward with the tongue from the palate. The forehead of tapinocephalids bore a spongy bony tissue. Earlier it was assumed that tapinocephalids could fight with horns; now this is doubted (their pachyostosis, bone overgrowth, was rather fragile).
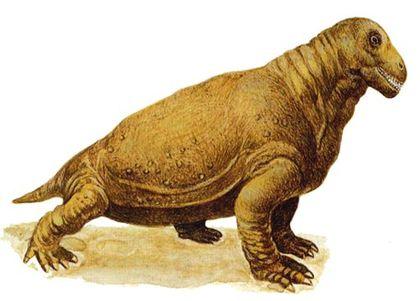
Among dinocephalians are also Estemmenosuchids – large animals with an impressive skull.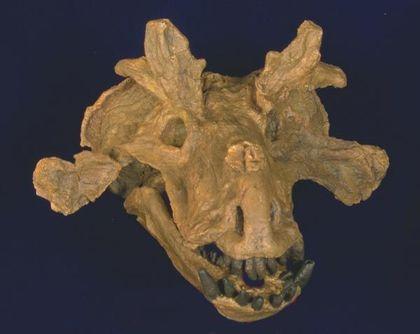
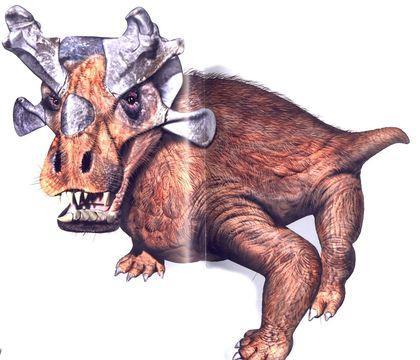

Gorgonopsians share the same division of teeth into functional groups that mammals inherited. Their chewing could not have been efficient (due to the lack of a secondary bony palate). One of the best‑known gorgonopsians is Inostrancevia, a serious predator that reached several metres in length.
It is likely that Inostrancevia was semi‑aquatic and could have hunted pareiasaurids (representatives of the parareptiles).
The skin of Inostrancevia was not dry and probably bore hair that served a tactile function.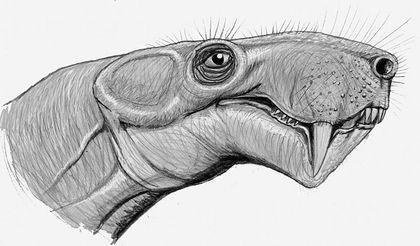
All dinomorphs were Permian animals, whereas representatives of the next order – the Anomodontia – passed from the Permian into the Triassic. Anomodonts were an extremely diverse group. The most successful among them were the dicynodonts, which survived until the end of the Triassic. The lower jaw of dicynodonts slid backward when the mouth closed, allowing them to bite off food (primarily plant material). Many dicynodonts lost almost all or all of their teeth and used a horny beak on the upper jaw for cropping.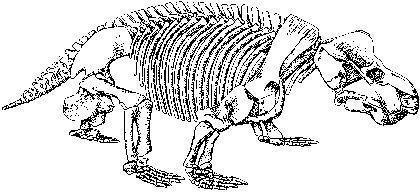
 (clickable)
(clickable)
Dicynodont skulls can be very expressive.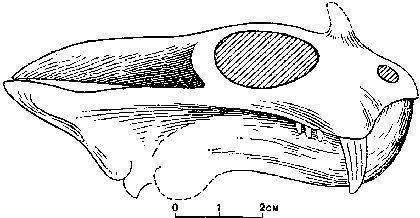
Finally, the infraclass Therapsida (literally “beast‑lizards”) includes the orders Therocephalia (“beast‑heads”) and Cynodontia (“dog‑toothed”). These organisms possessed a secondary bony palate and in many respects resembled mammals.
Speaking of therocephalians, one cannot omit the wonderful example of marital affection demonstrated by Volodymyr Prokhorovych Amalitsky. Amalitsky was the greatest researcher of Permian vertebrates at the end of the 19th – beginning of the 20th century; for example, he discovered Inostrancevia, which he named after his teacher A.A. Inostrantsev. He named a much more attractive therocephalian after his wife.
Therocephalians are a diverse group of Permian “reptiles”, many of which were small nocturnal animals. They are reasonably reconstructed as being fur‑covered.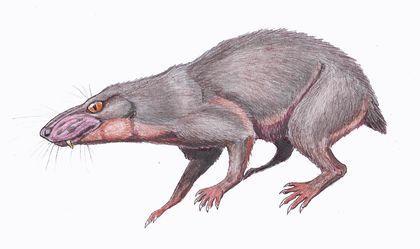
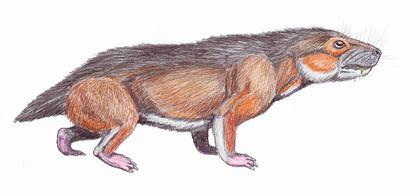
Finally, cynodonts are a group whose members would probably be hard to distinguish from mammals. Their teeth are not only differentiated; they acquire a complex shape that increases their functional efficiency.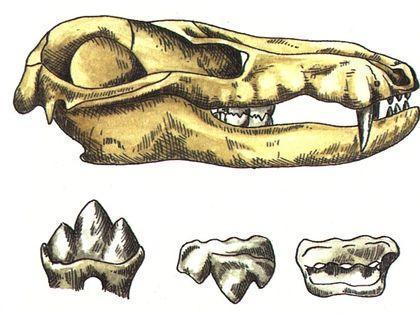
An important conclusion drawn from the evolution of therapsids is that over time the proportion of “mammal‑like” features characteristic of mammals increased in each of their branches.