
V. Reptilia sensu lato-09. Snakes
Fragments for now... This section is intended to cover the systematics of lepidosaurs, their distinctive features, and the smallest of the modern reptilian orders.
Unfortunately, a description of the major lizard groups is not yet available — only their classification is provided. As in other cases, groups that students are expected to know for the examination on this topic are highlighted in brick-red.
Suborder Ophidia (Serpentes) — Snakes
Snakes are a highly successful group of reptiles. Consider the reticulated python: it is a large, beautiful, and remarkably refined animal. Throughout the long evolutionary history of our ancestors, large snakes preyed upon them, and venomous members of this group posed a constant threat. The human brain possesses a dedicated population of neurons that detects serpentine movement and relays the signal — that something resembling a snake is present — directly to motivational centres. Some people fear snakes; others (like the author of these lines) are fascinated by them; virtually no one is truly indifferent. The emotional response to snakes is biologically determined, while its valence (positive or negative) depends largely on one's level of knowledge about snakes and the nature of prior experience with them.
This is the skeleton of a viper (Vipera sp.). It clearly illustrates the characteristic features of typical snakes. The body is elongated and the number of vertebrae is greatly increased. Limbs and their girdles are absent (in some snakes, rudiments of the hind limbs are retained). The thoracic cage is open ventrally; body shape is maintained by powerful musculature that produces lateral flexion of the vertebral column and drives rib movements.
The skull is kinetic, composed of movable units (in "typical" snakes). Its architecture is adapted for swallowing large prey, and in some snakes — such as this viper — also for injecting venom into prey.
Many lineages within Squamata have lost their limbs: amphisbaenians, several groups of limbless lizards, slow-worms, and skinks. Yet none of these groups acquired the ability to force their way between obstacles. This is not surprising. If a fossorial reptile pushes its passage through the substrate with its head, the skull will evolve into a compact, consolidated structure. It is likely that the ancestors of snakes burrowed through loose litter permeated by roots and stems through which they had to squeeze, and in doing so they evolved the capacity to flatten their bodies. Those lineages that specialised in subterranean life subsequently acquired consolidated skulls, while those that exploited the advantages of cranial kinesis became highly successful predators.
One line of evidence that snakes formerly led a fossorial lifestyle is found in comparisons of snake eyes with those of lizards: several muscles and their associated nerves have been lost in snakes. This may be interpreted as follows: the ancestors of snakes had no need for functional eyes, which began to degenerate, and later — as the mode of life shifted — eyes became necessary once more.
Worm-like snakes have the following appearance.
This Burmese python (Python bivittatus), engulfing a rat, exploits the mobility of the skull and thoracic skeleton.
Over evolutionary time, many snakes came to hunt prey that was quite large relative to their own body size.
The absence of limbs in snakes necessitates rather specialised modes of locomotion. The primary mode is, of course, lateral undulation.
Precise neuromuscular control of body movements enables effective arboreal locomotion, as demonstrated by this ratsnake.
In slender snakes, their flexible, muscular body easily supports its own weight.
Other modes of progression also exist — for example, rectilinear locomotion driven by the intercostal musculature.
Still more exotic is the "sidewinding" gait of snakes adapted to locomotion across loose sand.
The flexible and powerful body of snakes is ultimately capable of precise and refined movements. In the photograph below one can observe the combat ritual of male rattlesnakes: they pit their strength against one another by entwining their bodies. Similar tournaments are known in our own vipers.
Precise control of trunk movements requires considerable autonomy of the spinal cord. There exists a cruel pastime: upon encountering a snake, killing it — for instance, with a spade (individuals who commit this act frequently regard themselves as heroes in doing so). The decapitated body of a snake continues to writhe for a considerable time, and the severed head will even strike (as demonstrated in a disturbing video of a decapitated rattlesnake head).
One of the foremost evolutionary acquisitions of snakes is the ability to swallow large prey. In the photograph, a rock python (Python sebae) is swallowing an antelope. This is arduous "work": the python proceeds by alternately "walking" the premaxillary and dentary bones, in effect pulling itself over the carcass.
In some cases the prey is immobile and need not be killed beforehand. Observe how the African egg-eating snake (Dasypeltis scabra) swallows a quail egg. Once the ventrally directed hypapophyses of the cervical vertebrae crack the shell, the snake directs the liquid contents into the stomach and ejects the now superfluous shell membranes.
Mammals differ from most other tetrapods in being able to chew and breathe simultaneously. This is made possible by the separation of the nasopharynx from the oral cavity by the secondary bony palate. However, mammals cannot breathe and swallow simultaneously, whereas snakes can. Observe the oral cavity of this python.
Dorsally, two rows of tooth-bearing bones are visible: the maxillary and palatine tooth rows. Between the palatine teeth, a conical recess is visible into which the choanae (internal nares) open. Anterior to the choanae are the paired pits of the Jacobson's organ, the olfactory organ accessing the oral cavity.
In the centre of the "floor" of the oral cavity, a conical protrusion is visible: the glottis. When the snake's mouth is closed, this protrusion fits into the corresponding palatal recess, and the snake breathes air passing through the nostrils. It is worth noting that in snakes the nostrils are oriented such that the exhaled airstream is directed posteriorly rather than anteriorly, so as not to disturb prey being approached.
When swallowing prey, the snake extrudes a corrugated trachea from the glottis. The snake may simply extend this "air duct" to the corner of the mouth and continue breathing calmly while engulfing large prey.
But how is this large prey killed in the first place? One option is constriction. In this photograph, a boa (probably a garden boa) constricts a lizard (probably an anole). Swallowing its prey will not be particularly difficult for this snake...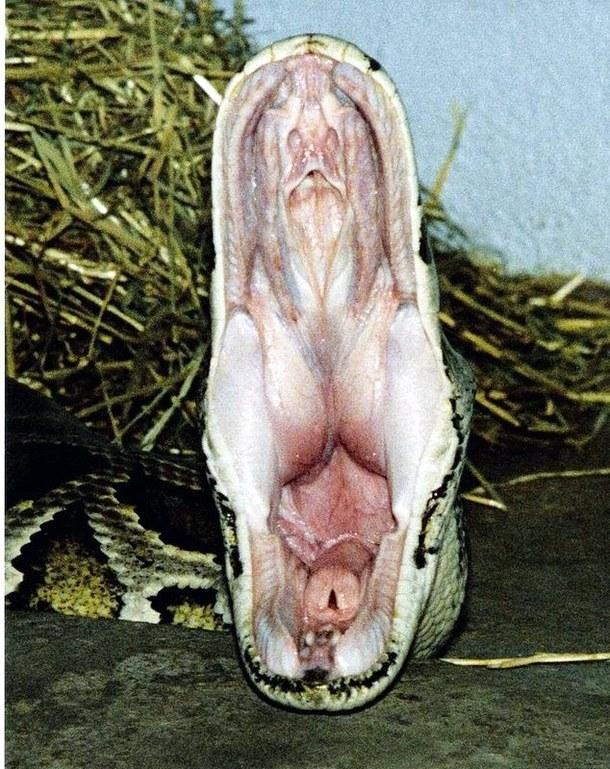
...whereas for this rock python engulfing an antelope, it is a considerable challenge. It must, of course, breathe throughout the process.
Certain evolutionary lineages of snakes have, however, devised a more refined solution for killing prey. Teeth are used (for grasping and retention) and saliva (for lubrication) during prey restraint and ingestion. The teeth introduce saliva into the prey's body, and the enzymes contained in saliva accelerate digestion. This is, incidentally, a very serious challenge. Digesting a large prey item before it putrefies within the snake's body represents a biochemical tour de force. If the temperature drops after a snake has fed well, the snake may be forced to regurgitate incompletely digested food (losing a reserve of digestive enzymes in the process). Any intensification of digestion is therefore an important and beneficial adaptation.
As a consequence, the dentition of many snakes has become specialised for the injection of venom into prey. A process of biochemical refinement of the venom is set in motion. Ultimately, the venom becomes so potent that it kills still-living prey or even serves to defend the snake against predators.
The photograph below shows a viper fang. It is covered by a dermal fold that retracts easily during a strike.
Venom delivery is a sophisticated operation demanding virtuosic mastery of the venom apparatus.
The evolution of the snake skull has been directed toward the development of an increasingly kinetic cranium, and subsequently toward providing mobility of the venom fangs (in viperids).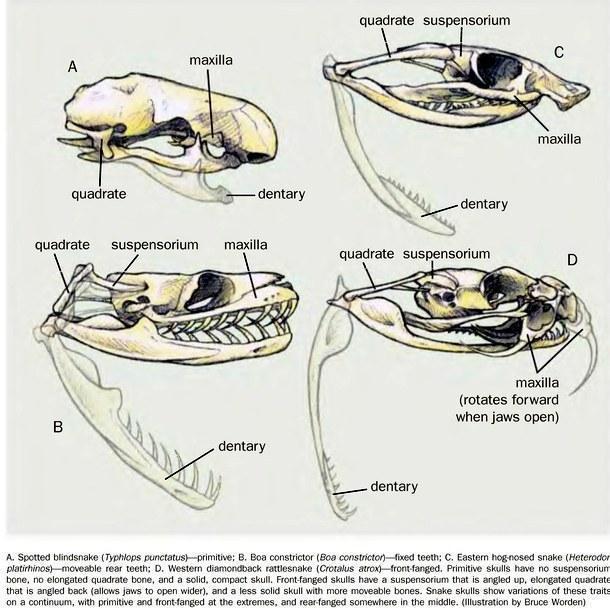
The following slide illustrates the evolution of venom-conducting dentition. From aglyphous snakes (Aglypha, not shown here) are derived opisthoglyphous snakes (Opisthoglypha) — with rear grooved fangs (such as the colubrid shown at the top) — and proteroglyphous snakes (Proteroglypha) — with front fixed-grooved fangs (sea snake and cobra); finally, certain proteroglyphous forms, such as the viper shown at the bottom, may be assigned to the solenoglyphous grade (Solenoglypha), bearing hollow, erectile fangs. The maxilla in solenoglyphous snakes is shortened and highly mobile. In advanced proteroglyphous and especially solenoglyphous snakes, the groove conducting venom becomes an enclosed tubular channel within the fang.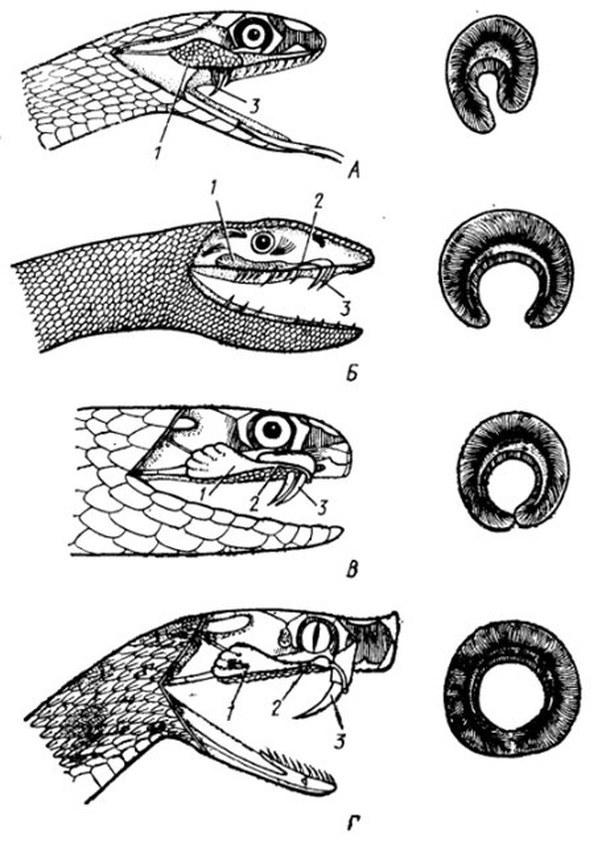
Snakes lack a middle ear; sound is conducted to the inner ear through the body tissues. This feature is shared with earless varanids and constitutes further evidence for the fossorial origins of snakes. Many snakes possess reasonably good vision and olfaction. The principal olfactory organ is of the contact type — the Jacobson's organ (vomeronasal organ). By flickering their tongue in the air and touching it to objects, snakes adsorb odorant molecules onto its surface; they then bring the tongue tips into contact with the Jacobson's organ and analyse the collected chemical stimuli.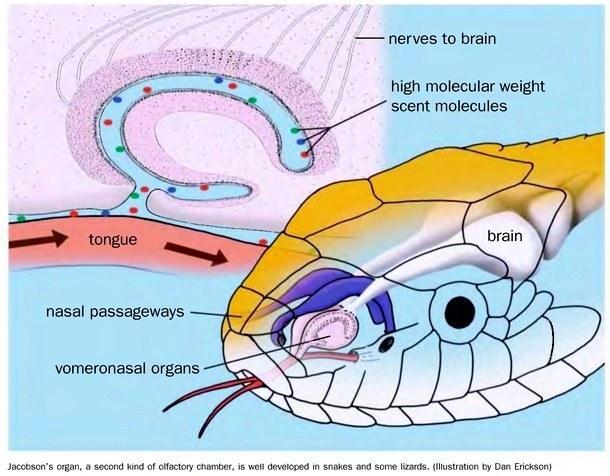
In pythons, boas, vipers of the genus Bitis, and especially in pit vipers (subfamily Crotalinae), heat-sensitive pit organs develop on the head.
The eyes of snakes become regularly clouded. This occurs as the old epidermis — the cornified layer of the skin — separates from the underlying tissue.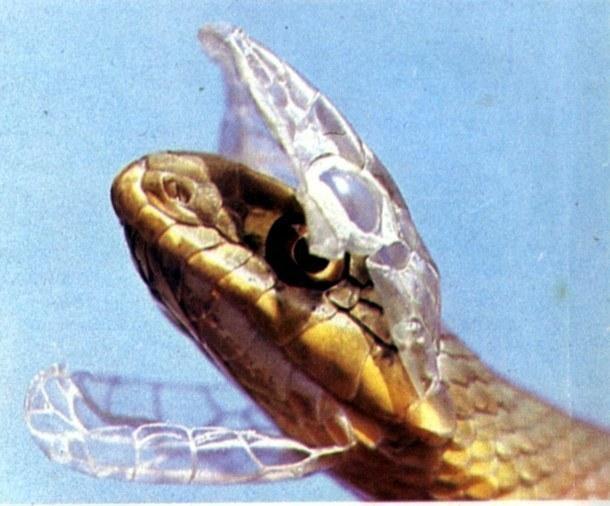
The clouded eyes will clear (when the old "skin" separates from the new layer), after which the old epidermis sloughs off the snake in a single piece, beginning from the mouth.
The result of ecdysis is the dry, intact shed skin (exuviae).
The coloration of snakes is highly diverse, often extraordinarily beautiful, yet generally entirely functional.
Developmental anomalies periodically give rise to bicephalic snakes, which sometimes survive for a considerable period.

In snakes, the full spectrum of reproductive modes is known: from oviparity through ovoviviparity to true viviparity. Common grass snakes (Natrix natrix) deposit their eggs in warm, sheltered locations (several females sometimes share a communal clutch site).
Pythons, such as this reticulated python (Malayopython reticulatus), guard their clutch and additionally warm it by periodically contracting and relaxing their musculature.
The hatchlings emerge fully capable of independent life. Frequently, after slitting the leathery eggshell with the egg tooth (a transient embryonic structure located at the tip of the snout, which soon falls off), the neonates remain within the egg for a period, breathing fresh air in the interim. The photograph below shows a neonate copperhead or some other pit viper.
And this shows parturition in a rattlesnake, which also belongs to the pit vipers.