
Vegetina, Biryuk, Shabanov (2014) Comparison of spermatogenesis stability
Veherina A. O., Biryuk O. V., Shabanov D. A. Sravneniye ustoychivosti spermatogeneza mezhvidovykh poluklonalnykh gibridov, Pelophylax esculentus, i obitayushchego sovmestno s nimi odnogo iz roditelskikh vidov, Pelophylax ridibundus. – Pratsi ukrainskoho herpetolohichnoho ...
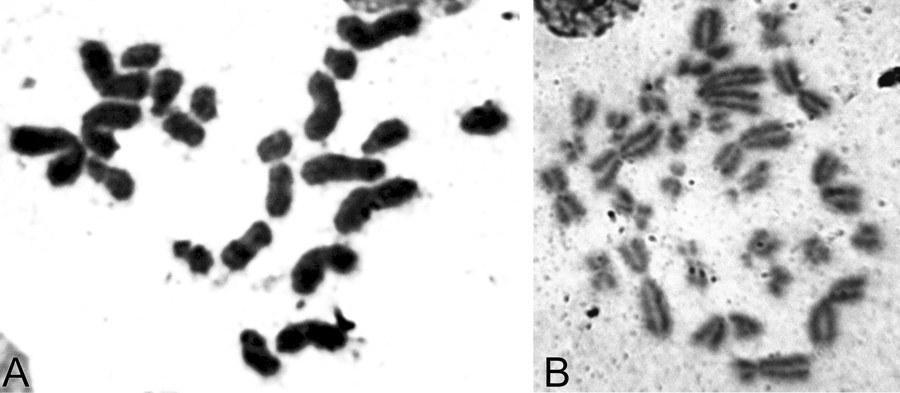
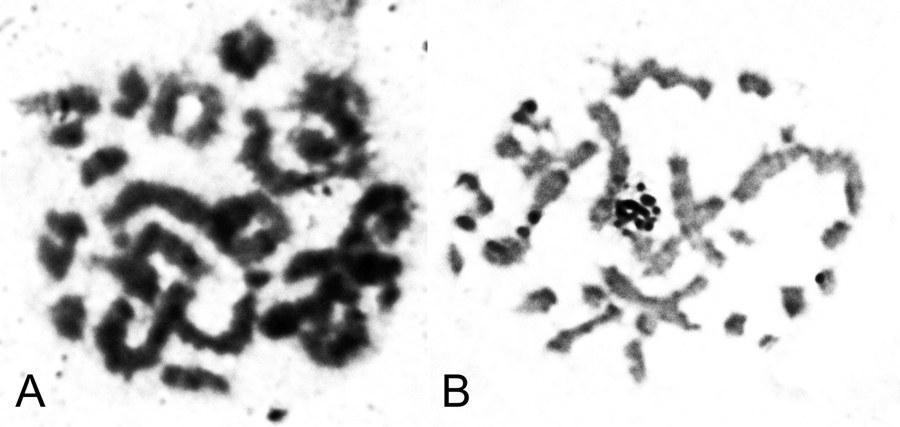
UDC 597.851(576.354.4) Comparison of spermatogenesis stability in hemiclonal interspecific hybrid Pelophylax esculentus and parental species Pelophylax ridibundus (Amphibia, Anura) A.O. Vegerina, O.V. Biriuk, D.A. Shabanov V.N. Karazin Kharkiv National University, Svobody Sq., 4, Kharkiv, 61077 Ukraine E-mail: anastasia.vegerina@gmail.com; mykhailova.o.v@gmail.com; d.a.shabanov@gmail.com Comparison of spermatogenesis stability in hemiclonal interspecific hybrid Pelophylax esculentus and parental species Pelophylax ridibundus (Amphibia, Anura). Vegerina A.O., Biriuk O.V., Shabanov D.A. — The article presents the results of karyological study of male germline cells of interspecific hybrid Pelophylax esculentus and one of its parental species P. ridibundus . Aneuploid cells were detected in both, hybrids and parental species individuals, but the average frequency of spermatocytes I with the wrong number of bivalents is significantly higher in P. esculentus . The range of changes in the normal cells proportion among the individualsin the sample of hybrids varies significantly wider than among P. ridibundus . We have concluded that the comparative instability of hemiclonal inheritance mechanism leads to difficulties in spermatogenesis and reduction of the interspecific hybrids reproductive efficiency. Key words: Pelophylax esculentus , spermatogenesis, abnormalities, aneuploidy, hemiclonality. Introduction The hybridogenetic complex of green frogs ( Pelophylax esculentus complex) includes two parental species: pond frog, Pelophylax lessonae (Camerano, 1882) and marsh frog, Pelophylax ridibundus (Pallas, 1771), as well as their hybrids (Berger, 1964). For a number of reasons, the name analogous to the species name is used to denote interspecific hybrids — edible frog, Pelophylax esculentus (Linnaeus, 1758). As a rule, in P. esculentus , only one parental genome is transferred to the gametes, which is inherited clonally without recombination over generations (Tunner, 1974). Such reproduction is called hemiclonal (Plötner, 2005). Typically, P. esculentus inhabit and reproduce together with representatives of one or both parental species, forming hemiclonal population systems, HPS (Shabanov, Litvynchuk, 2010). For the region named the Seversky-Donets center of diversity of green frogs, HPS consisting of diploid and triploid P. esculentus , as well as P. ridibundus are characteristic (Shabanov, Litvynchuk, 2010). P. lessonae is absent in this region, and all its genomes are transmitted through interspecific hybrids, P. esculentus . Hemiclonal reproduction of hybrids is ensured by a specific type of gametogenesis, which is essentially abnormal (compared to the typical one characteristic of representatives of the parental species). The genomes of the parental species of the P. esculentus complex differ so much that typical meiosis in a hybrid is impossible. The adaptation that allows bypassing this difficulty is that one of the parental genomes is removed from the germline cells before meiosis begins. In a series of cytogenetic studies using fluorescence microscopy, Austrian authors H. Tunner and S. Heppich-Tunner showed that the removal of the non-clonal genome occurs during mitotic divisions of germline cells. After that, the clonal genome is duplicated, and this process occurs before the cells enter meiosis, or in females, it can occur during meiosis (Tunner, Heppich-Tunner, 1991). In triploid individuals, gametogenesis proceeds similarly, except for the duplication stage (Vinogradov et al., 1991; Plötner, 2005). As a result of such abnormal processes in hybrid frogs, compared to representatives of the parental species, reproduction faces a number of difficulties, including gonadal development disorders, larval development anomalies, mosaic individuals, and reduced viability and lifespan (Berger, 2008; Mikhailova et al., 2011). The peculiarities of gametogenesis in representatives of the Seversky-Donets center of diversity of green frogs have been studied using various methods, including flow DNA cytometry of sperm suspension (Borkin et al., 2005); obtaining metaphase plates from disrupted cells (Suryadnaya, 2003; Manilo et al., 2007); and electrophoretic analysis of germ cell suspension (Mezhzherin et al., 2007; Morozov-Leonov et al., 2009). Studies on oogenesis of P. esculentus have also been conducted using lampbrush chromosome markers and FISH techniques (Dedukh et al., 2015). Among other things, karyoanalysis in squashed preparations was used to study spermatogenesis (Mikhailova et al., 2011). Based on the results of the listed studies, it can be stated that in many cell lines of hybrids, gametogenesis, which can be considered normal for them (given its abnormality compared to the typical case of recombinant inheritance), is disrupted. The consequence of such disruptions is the appearance of a significant number of aneuploid and polyploid germline cells (most likely, the vast majority do not reach the stage of mature gametes). This means that in P. esculentus , due to hemiclonal inheritance, the gamete formation process is unstable. However, the results obtained are descriptive; a quantitative assessment of gametogenesis disorders that would allow evaluating the burden of hemiclonality has not been performed to date. Based on the foregoing, it can be stated that comparing the stability of spermatogenesis in diploid interspecific hybrids of green frogs ( P. esculentus ) and parental species of the hybridogenetic complex is of significant interest. In this regard, the task of this work was to describe and count abnormal mitotic and meiotic plates in the testes of P. esculentus hybrids and one of the parental species, P. ridibundus . For this, we used the karyoanalysis method on squashed cell preparations. Materials and Methods We studied a sample of 23 green frogs, including 10 diploid males of P. esculentus and 13 males of P. ridibundus from the Seversky-Donets center of diversity of the P. esculentus complex. The frogs were caught from June to October 2013 in the vicinity of the villages of Gaidary, Genievka, and Sukha Gomolsha in the Zmiiv district of the Kharkiv region, as well as within the city of Kharkiv (Timurovtsiv Street). The species affiliation of the frogs was determined by a complex of external characteristics (Shabanov et al., 2006). For individuals collected within the city, the genome composition was determined using flow DNA cytometry, performed by Yu.M. Rozanov and S.N. Litvynchuk at the Center for Molecular Genetics, Russian Academy of Sciences (Moscow). For karyological analysis, fragments of intestines and testes were taken from the animals. To indirectly determine ploidy in green frogs, blood smears were obtained and examined according to a published method (Bondareva et al., 2012). Each studied individual received an intraperitoneal injection of 0.04% aqueous colchicine solution (0.1–0.3 ml, depending on body weight) 24 hours before sacrifice. The materials obtained during dissection were washed, cut into fragments, and placed in a hypotonic solution (0.07 M KCl) for 20 minutes. Then, the solution was replaced with Carnoy's fixative (3 parts methanol and 1 part glacial acetic acid), with the fixative being changed three times every 30 minutes. Karyological preparations were made from the fixed materials by the squashing method. For this, a tissue fragment was transferred to a 70% acetic acid solution, where it was macerated to form a cell suspension. The resulting suspension was applied to glass slides heated to 60°C on a heating stage using a Pasteur pipette in the form of drops 1 cm in diameter. The applied drops were immediately removed with a pipette, resulting in a small number of individual cells remaining on the slides. The preparations were dried and placed in an incubator at 37°C for 3 weeks. Final ploidy determination was performed by counting chromosomes (26 for diploids or 39 for triploids) in at least seven metaphase plates from somatic tissue of each individual. If there were insufficient metaphase plates suitable for chromosome counting, ploidy was determined based on the number of nucleoli in the nuclei of at least 20 cells. For this, we used silver nitrate staining (silver staining) — a method used to detect the nucleolar organizer region (NOR, 18S + 28S rDNA sites) in metaphase chromosomes, as well as nucleoli in interphase nuclei of somatic cells (Birshtein, 1984; Schmid, 1982). As a result of silver staining, nucleoli become clearly visible as dark areas: two in the nuclei of diploid individuals, and, accordingly, three in the nuclei of triploid individuals (Vegerina et al., 2013). The silver nitrate-stained preparations were placed in a 2% Giemsa stain solution, then rinsed in distilled water and dried. All studied individuals turned out to be diploid. On the obtained testis preparations, cells at different stages of mitotic or meiotic division were selected. Metaphase plates suitable for analysis were viewed under magnifications of 160, 640, and 1600 times with oil immersion and photographed using a digital USB microscope camera (ScienceLab View.7). On the obtained photographs, the structures in the plates at the metaphase stage of mitosis, as well as the diakinesis and metaphase I stages of meiosis, where the number and structure of chromosomes or bivalents are most clearly visible, were counted. Statistical processing of the obtained data was performed using the Statistica program. Results and Discussion In the testes of green frogs, we registered a significant number of aneuploid cells, i.e., cells with a chromosome number not a multiple of the haploid set. Aneuploid cells were observed at different stages of gametogenesis, both in hybrid individuals and in individuals of the parental species. The number of chromosomes in such cells varied within very wide limits. While the diploid set in green frogs is 26 chromosomes (Fig. 1), we registered hypohaploid (less than 13), hyperhaploid (more than 13), hypodiploid (less than 26), and hyperdiploid (more than 26) plates (Fig. 2). The presence of hypohaploid, hyperhaploid, and hypodiploid plates could be partially explained by losses during the preparation of karyological specimens by the squashing method, or, in other words, attributed to methodological error. However, it should be taken into account that in the variant of karyoanalysis used by us, not free-floating metaphase plates (which increases the probability of losing some chromosomes or overlapping plates) but whole cells were squashed, which sharply reduces the probability of errors. It would be even more difficult to explain the appearance of hyperdiploid plates as a methodological error. Their presence directly indicates disturbances in the gametogenesis mechanism characteristic of P. esculentus . Finally, it should be noted that the spatial distribution of chromosomes indicates that the increase in ploidy occurred as a result of segregation errors, not overlapping of different plates. Fig. 1. Spermatogonia of a diploid male P. esculentus . 2n metaphase of mitosis, 26 chromosomes: A — metaphase plate; B — karyogram. Fig. 2. Aneuploid spermatogonia of diploid P. esculentus males. Hypodiploid chromosome set in metaphase of mitosis, 22 chromosomes: A — metaphase plate; B — karyogram. Hyperdiploid chromosome set in metaphase of mitosis: C — metaphase plate; D — karyogram. In meiosis, we also observed aneuploid and duplicated sets of bivalents, and, in addition, disturbances in the formation of bivalents themselves — univalents, the number of which, in turn, could also be a non-multiple of the haploid set (Fig. 3). It should be noted that other authors studying female P. esculentus have also noted aneuploid cells among oogonia (Tunner, Heppich-Tunner, 1991), as well as oocytes containing univalents, a doubled number of bivalents, and even bivalents and univalents simultaneously (Dedukh et al., 2015). In the testes of three diploid males of P. esculentus , in addition to aneuploid plates, polyploid plates — 3n and 4n — were found (Fig. 4). It should be emphasized that in these individuals, only diploid chromosome sets were observed in somatic tissues, and the erythrocyte size also varied within the limits corresponding to the size of diploid cells (Bondareva et al., 2012). Fig. 3. Spermatocytes I of diploid P. esculentus males: A — prophase I of meiosis, 13 bivalents (normal); B — prophase I of meiosis, 26 bivalents; C — 13 univalents. Fig. 4. Spermatogonia of a diploid male P. esculentus . Metaphase of mitosis, 51 chromosomes (normal — 4n = 52), one chromosome in the 5th pair is missing: A — metaphase plate; B — karyogram. In general, the pattern of anomalies registered for P. esculentus corresponded to the results of previous studies conducted by us using a different method (Mikhailova et al., 2011).
Furthermore, these results coincide with data from other authors on frogs from the Kharkiv region (Suryadnaya, 2005; Manilo et al., 2007, Dedukh et al., 2015), and also echo the data obtained from studying populations of green frogs from Western Europe (Günther, 1975; Tunner, Heppich-Tunner, 1991). In the testes of P. ridibundus , abnormal cells with an incorrect number of structures were found less frequently than in hybrid individuals.
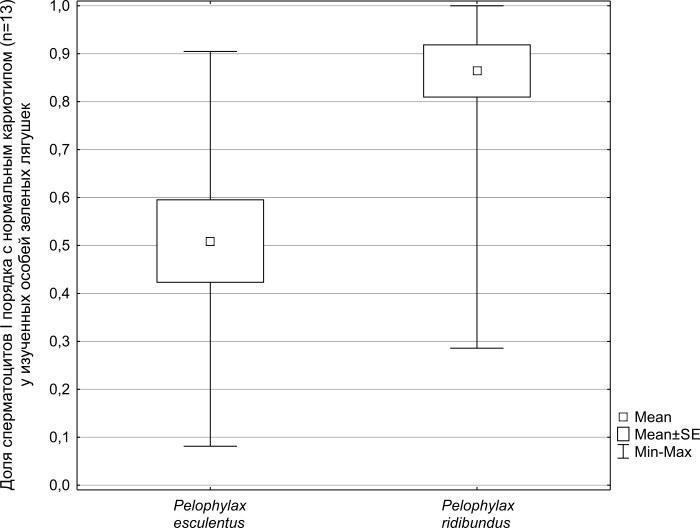
All registered abnormalities in P. ridibundus were analogous to those described above for P. esculentus . In one of the marsh frog males, we registered a polyploid cell with a chromosome set exceeding the triploid number (n = 47). This was likely a tetraploid plate that had undergone losses. Such a deviation from the norm in P. ridibundus has been registered and described previously by other authors (Günther, 1975; Manilo, Radchenko, 2010). Mann-Whitney U-test analysis of the differences between the hybrid and parental species samples in the number of normal cells (with thirteen bivalents) showed that the difference between these samples is significant (p = 0.0015). It is clearly visible (Fig. 5) that the average proportion of normal spermatocytes out of their total number for the studied P. ridibundus individuals (0.86) is more than one and a half times higher than that for P. esculentus (0.51), and the range of changes in this proportion among edible frog individuals is significantly wider. Overall, the proportion of normal spermatocytes I among all studied plates of P. ridibundus was 0.75, and for P. esculentus — 0.40. The lower overall proportion of normal cells in P. ridibundus among the cells we studied (0.75) differs from the average proportion of such cells we established for this species (0.86). This difference is a consequence of our more thorough examination of parental species individuals with an increased frequency of karyotype anomalies: we studied a larger number of cells from each such individual.
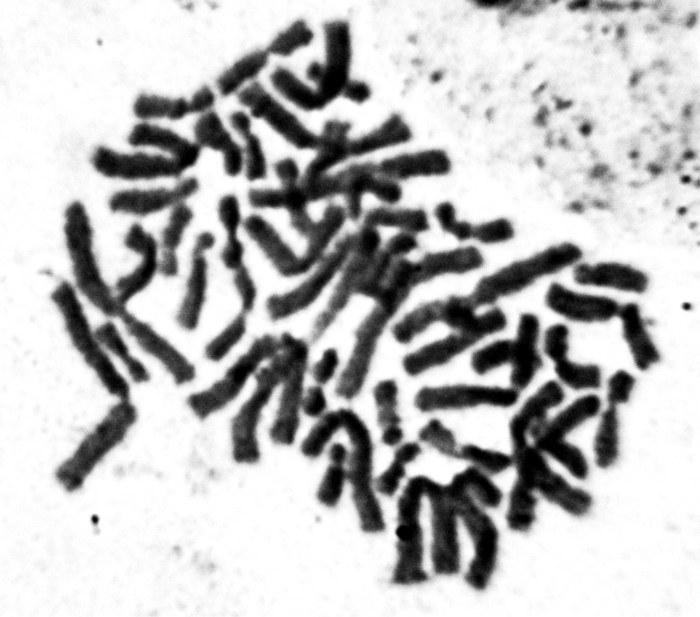
Fig. 5. Mean values and confidence intervals of spermatocytes I with a normal amount of bivalents (n = 13) share.
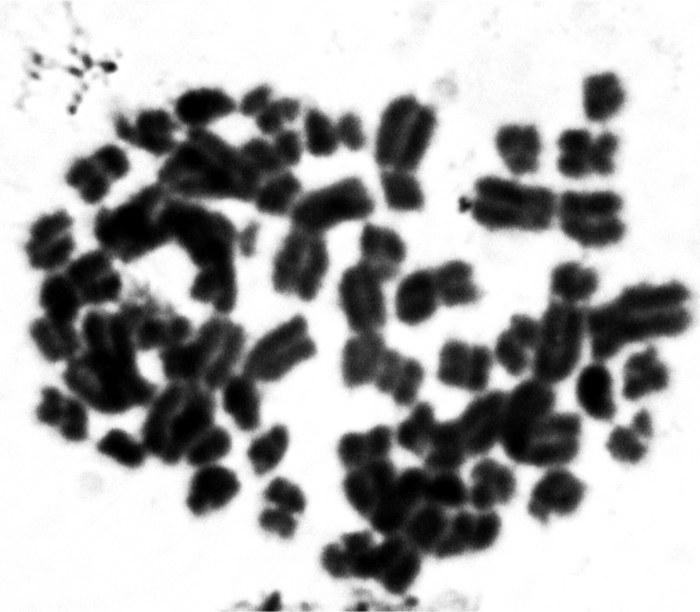
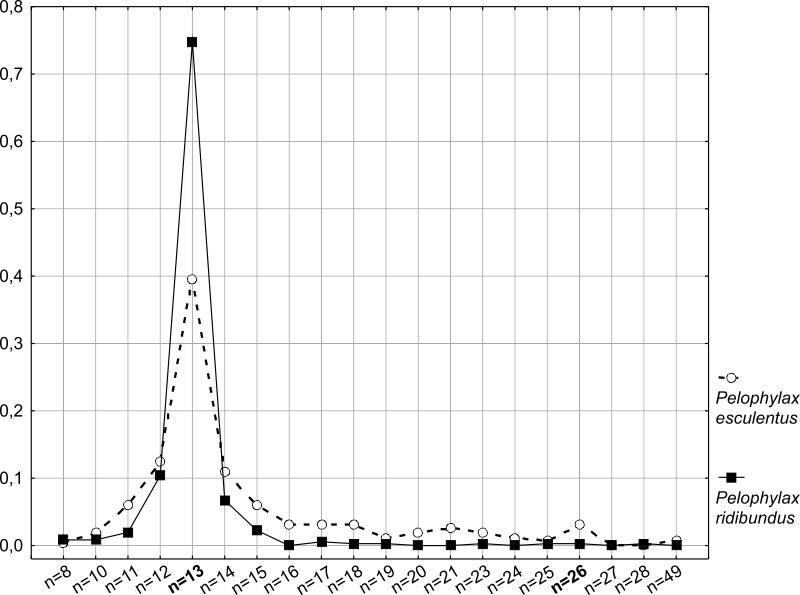
Figure 6 shows the frequency distribution of spermatocytes I with a specific number of bivalents. It is clearly seen that the frequency of cells with an incorrect number of bivalents is significantly higher for P. esculentus . Moreover, the range of variation in the number of structures in spermatocytes is somewhat wider in edible frogs than in marsh frogs. Probably, a significant proportion of such abnormal cells are eliminated at subsequent stages of spermatogenesis. It is possible, however, that some of these cells may pass through all stages of gametogenesis, giving rise to aneuploid spermatozoa. The presence of a small peak for n = 26 in P. esculentus is also interesting. This means that in the testes of diploid hybrids, a certain number of tetraploid cells are formed, which could then lead to the formation of diploid sperm. Despite more than a decade of studying the Seversky-Donets center of diversity of green frogs, the question of the crosses that lead to the formation of triploid P. esculentus in this region remains unclear. The formation of diploid sperm by diploid hybrid males is one potential explanation for the phenomenon of triploid appearance. Fig. 6. The arrangement of different bivalent sets frequencies of occurrence in the totality of P. ridibundus and P. esculentus cells studied. Thus, it can be concluded that hemiclonality in interspecific hybrids of green frogs is a less stable inheritance mechanism than the formation of recombinant gametes characteristic of representatives of the parental species. The relative instability of hybrid gametogenesis should lead to a decrease in their reproductive efficiency. However, it should be taken into account that, even considering the described anomalies, hemiclonal inheritance, as a naturally occurring method of overcoming hybrid sterility, ensures the mass reproduction of P. esculentus when crossed with parental species. In the opinion of the authors, the methodological approach tested in this work needs to be applied to a wider range of material. It is desirable to assess the proportion of anomalies during spermatogenesis in another parental species, P. lessonae . It is of interest to determine how the level of anomalies in spermatogenesis of F1 hybrids, resulting from crosses between parental species, compares with that of hybrids that have transmitted the clonal genome over a significant number of generations.
It can be assumed that the level of gametogenesis anomalies in parental species inhabiting HPS along with hybrids will differ from that of representatives of the same species from typical monotypic populations that do not contact hybrids. Bondareva A.A., Bibik Yu.S., Samilo S.M., Shabanov D.A. Cytogenetic features of green frog erythrocytes from the Seversky-Donets center of diversity of the Pelophylax esculentus complex // Visnyk Kharkivskoho natsionalnoho universytetu imeni V.N. Karazina. Ser. Biolohiia. — 2012. — Issue 15, No. 1008. — P. 116–123. — /Bondareva_2012_Cytogenetic Borkin L.Ya., Zinenko A.I., Korshunov A.V. et al. Mass polyploidy in the hybridogenetic complex Rana esculenta (Ranidae, Anura, Amphibia) in Eastern Ukraine // Proc. I Conf. Ukrainian Herpetological Society. — K.: Zool. Mus.
National Academy of Sciences of Ukraine, 2005. — P. 23–26. — /Borkin_et_al_2005_Polyploidy Vegerina A.O., Meleshko E.V., Pyrina I.S. et al. Determination of the ratio of diploids and triploids among metamorphs of green frogs in the Seversky-Donets center of diversity of the Pelophylax esculentus complex // Visnyk Kharkivskoho natsionalnoho universytetu imeni V.N. Karazina. Ser. Biolohiia. – 2013. — Issue 18, No. 1079. — P. 107–113. — /Vegerina_et_al_2013_Metamorphs . Manilo V.V., Radchenko V.I., Korshunov A.V. Study of the karyotype of the edible frog ( Rana kl esculenta ) from the Kharkiv region of Ukraine // Nauk. Visnyk Uzhhorod. Univ. Ser. Biolohiia. — 2007. — Issue 21. — P. 68–73. Manilo V.V., Radchenko V.I. Karyological study of Pelophylax ridibundus (Anura, Amphibia) from the eastern part of Ukraine // Zbirnyk prats Zoolohichnoho muzeiu. — 2010. — No. 41 — P. 111–121. Mikhailova O.V., Kechedzhi A.E., Shabanov D.A. Study of spermatogenesis in diploid Pelophylax esculentus (Amphibia, Anura) using karyoanalysis in squashed preparations // Pratsi Ukrainskoho herpetolohichnoho tovarystva. — 2011. — No. 3. — P. 120–127. — /Mikhailova_et_al_2011_Spermatogenesis_diploids Mikhailova O.V, Usova O.Ye., Shabanov D.A. How to assess the population burden associated with hemiclonal hybridization in population systems of the Pelophylax esculentus complex? // Biolohiia ta Valeolohiia. — Issue 13 — Kharkiv : KhDPU, 2011. — P. 44–50. Morozov-Leonov S.Yu., Mezhzherin S.V., Nekrasova O.D. et al. Inheritance of parental genomes by the hybrid form Rana “ esculenta ” (Amphibia, Ranidae) / // Genetika. — 2009. — Vol. 45, No. 4. — P. 488–495. Suryadnaya N. N. Materials on the karyology of green frogs ( Rana ridibunda , Rana lessonae , Rana esculenta ) from the territory of Ukraine // Vestn. Zool. — 2003. — 37, No. 1. — P. 33–40. Shabanov D.A., Zinenko A.I., Korshunov A.V. et al. Study of population systems of green frogs ( Rana esculenta complex) in the Kharkiv region: history, current state, and prospects // Visnyk Kharkivskoho natsionalnoho universytetu im. V.N. Karazina. Seriia: Biolohiia. — 2006. — Issue 3, No. 729. — P. 208–220. Shabanov D.A., Litvynchuk S.N. Green frogs: life without rules or a special way of evolution? // Priroda. — 2010. — No. 3 (1135). — P. 29–36. — /Frogs Dedukh D., Mazepa G., Shabanov D. et al. Optional endoreplication and selective elimination of parental genomes during oogenesis in diploid and triploid hybrid european water frogs // Plos One. — 2015. — 10, N 4. — P. 1–19. Berger L. Is Rana esculenta lessonae Camerano a distinct species? // Ann. Zool. PAN. — 1964. — 22, N 13. — P. 245–261. Berger L. European green frogs and their protection. — Poznan : Fundacja Biblioteka Ekologiczna, 2008. — 72 p. Birstein V.J. Localization of NORs in karyotypes of four Rana species // Genetica. — 1984. — N 64. — P. 149–154. Günther R. Untersuchungen der meiose bei mänchen von Rana ridibunda Pall., Rana lessonae Cam. und der bastardform „ Rana esculenta “ L. (Anura) // Biologisches Zentralblatt. — 1975. —.94, N 3. — S. 277–294. Plötner J. Die westpaläarktichen Wasserfrösche. — Bielefeld : Laurenti-Verlag, 2005. — 161 S. Schmid M. Analysis of the Structure and Variability of NORs in Anura // Chromosoma. — 1982. — N 87. — P. 327–344. Tunner H. G. Die Klonale Struktur einer Wasserfroschpopulation // Z. zool. Syst. und Evolut.-forsch. — 1974. — 12, N 4. — S. 309–314. Tunner H.G., Heppich-Tunner S. Genome Exclusion and Two Strategies of Chromosome Duplication in Oogenesis of a Hybrid Frog // Naturwissenschaften. — 1991. — 78. — P. 32–34. Vinogradov A.E., Borkin L.J., Günther R., Rosanov J.M. Genome elimination in diploid and triploid Rana esculenta males: cytological evidence from DNA flow cytometry // Genome. — 1991. — 33. — P. 619–627.