Каптриноморфы (Сaptrinomorpha) – группа примитивных ангуститабуляров с апопарейальной конструкцией крыши черепа, унаследованной от примитивных предков, что выражается в отсутствии широкого подстилания надвисочной кости, чешуйчатой.
____ Время существования и места известных находок ____
Считается, что данная группа рептилий появляется в среднем карбоне (Канада), чуть раньше пеликозавров и ареосцелидий. Первые находки (Канада) были не многочисленны (периоду среднего карбона). Более частые находки относятся к периоду верхнего карбона (Северная Америка). "Рассвет" эта группа начала достигать предположительно в нижней (Западная и Восточная Европа) и верхней перми (Китай, Восточная Европа, Северная Африка, Зимбабве, Южная Африка).
____ Систематика ____
Согласно Олсону капториниды происходят от эврептилий (Olson, 1947). В основном капториноморф выделяли, как предков зверообразных рептилий (Watson, 1957), однако после развития концепции парарептилий (Olson, 1947) их все более вероятней начали относить к предкам всех эврирептилий (Carroll, 1991). М.Ф. Ивахненко (2008) представляет нам трактование формальности о том, что в меру особенностей конструкции черепа, в состав капториноморф должны входить все примитивные апопарейальные ангуститабуляры со стегальным черепом, т.е антракозаморфы и производные от них (Diadectomoppha), за исключением Bolosauridae. В противовес, в таком случае, Tseajaidae сближают с Limnoscelidae. Хюне в 1956 году, предпринял попытки объединения каптриноморф с миллерозаврами в один подкласс Captorhinidia примитивных эврептилий. Некоторые источники говорят, что данная группа является единственной, у которой просматривается некое сходство в строении черепа с нижнекарбоновыми Westlothiana ( Татаринов, 2006). Эти же источники указывают на то, что некоторые автора выделяют примитивных капториноморф в семейство проторотирид (Protorothyridae = Romeriidae).
Для систематического выделения, считают приемлемым воспользоваться краниальным признаком (как и в случае с парарептилиями). Этим признаком является степень редукции костей затеменного ряда (заднетеменных и таблитчатых). Предположительно это связано со степенью развития шейной мускулатуры, что соответствует относительно наземному образу жизни. Если опираться на таких автаров, как Татаринов, можно говорить о том, что, всего описано более 22 родов каптриноморф. Подкласс Captrhrinomorpha включает в себя 2 отряда :
-
Отряд Сaptorhinida (1 семейство, 2 подсемейства, 15 родов)
-
Отряд Bolosaurida (2 семейства, 7 родов)
Отряд Сaptorhinida
-
Семейство Captorhinidae (Case, 1911)
- Подсемейство Сaptorhininae (Case, 1911)
- Род Captorhinus (Case, 1895)
- Род Riabininus ( Иванченко, 1990)
- Род Romeria (Price, 1937)
- Род Labidosaurus (Cope, 1896)
- Род Protocaptorhinus (Clark et Carroll, 1973)
- Род Rhiodenticulatus (Berman et Reisz, 1986)
- Род Acrodonta (Dutuit, 1976)
- Род Saurorictus (Modesto et Smith, 2001)
- Подсемейство Moradisaurinae (de Ricyles et Taquet, 1982)
- Род Moradisaurus (Taquet, 1969)
- Род Gecatogomphius (Vjuschkov et Tchudinov, 1957)
- Род Kahneria (Olson, 1962)
- Род Rhotianiscus (Olson, 1935)
- Род Lebidosaurikos (Stovall, 1950)
- Род Captorhinikos (Olson, 1954)
- Род Moradisaurus (Taquet, 1969)
Отряд Bosaurida
-
Семейство Bolosauridae (Cope, 1878)
- Род Bolosaurus (Case, 1878)
- Род Stepnospohdylus (Stappenbeck, 1905)
- Род Timanosaurus (Gubin, 1993)
- Род Gnirhimosuchus (Efremov, 1951)
-
Семейство Belebyidae (Ivachenko, 2001)
- Род Beley (Ivachnenko, 1973)
- Род Davletkulia (Ivachnenko, 1990)
- Род Permotriturus (Tatarinov, 1968)
____ Морфология ____
Имея перед глазами систематический состав можно предоставить некоторые аспекты морфологических особенностей представителей данного подкласса.
Как правило, представителей данного таксона разделяют на мелких всеядных/насекомоядных с остроконическими зубами (отряд Captorhinida) и более крупных растительноядных с развитым вторичным небом и обладающих субапсидным окном (отряд Bolosaurida). Их представители имели стегальную крышу (без височных окон) или только с одним субапсидным окном. Таблитчатая и заднетеменная (кости затеменного ряда) значительно редуцированны; не выходят на дорсальную поверхность крыши черепа и могут сохраняться только в виде затылочных пластин.Иными словами таблитчатые и парные заднетеменные кости отнесены на затылок, затылочное задневисочное окно отсутствует. Постквадратная ушная вырезка отсутствует. Имеющаяся чешуйчатая кость узкая и развита только вдоль задне-бокового угла теменной кости, а чешуйчатая кость относительно широкая и обосабливает надвисочную кость от заглазничной. Парокципитальный отросток заднеушной кости зачаточный и не достигает ни чешуйчатой, ни таблитчатой кости. Небо с подвижным базиптеригодным сочленением, небольшой межптеригоидной впадиной и хорошо развитыми поперечными отростками птеригоидов с выраженными поперечными флангами.
Представители отряда Captorhinida характеризуются относительно массивным, суженным черепом в котором отсутствуют височные окна и второе небо (палатинальные фланги) в челюстях. Зубы могут быть остроконические, притупленно-цилиндрические, многоядерные. Представители Protorothyridae, имеющее спорное систематическое положение, известны как мелкие ящеры с черепом 4-5 см, тело с хвостом и относительно короткими конечностями достигает 35-50 см. Питались, предположительно, приемущественно насекомыми или мелкой животной пищей. Череп, исходя из общих черт, похож на соленодонзавровый, однако, без ушной вырезки. На парасфеноиде и птеригоидах с отростками, имеются ряды небных зубов, иногда заходящие и на небные кости, однако без шагреневых полей мелких зубов на небе. Слуховая косточка "теропсидного" типа, на концах лежит по направлению к quadratum , над областью медиального мыщелка челюстного сочленения. В позвоночнике до 32 предкресцовых позвонков, атлант и эпистрофей проторептилийного типа с самостоятельным интерцентром последнего. Невральные дуги позвонков не упрощены (в отличаи от сеймуриаморф, диадектоморф). Межключитницца Т-образная. Тазовый пояс пластинчатый, сочленовный отросток направлен назад, верхний отросток антракозаврового типа отсутствует (Holmes, 1977). Стопа, как правило, имеет не полностью сросшийся рептилийный астрагал. Брюшные ребра - поперечные ряды удлененных чешуек.
В отличаи от проторотид, типичные персмких капториниды несколько крупнее. На их возможную моллюскоядность указывает многорядные притупленные челюстные зубы. Парокципитальный отросток у капторинид удлиняется и соеденяется с чешуйчатой костью. Таким образом можно говорить, что они достигают сеймуриаморфного уровня. Типичные капториниды утрачивают табличные кости и эктоптеригоиды, также, как и зубы на парасфеноиде. Невральные дуги на позвонках расширены (в отличаи от проторотид). Имеются два кресцовых позвонка. Особенностью является обособленная лопатка плечевого пояса с двумя коракоидами. Некоторые авторы выводят в тенденцию образование у капторинид комплексного астрагала за счёт слияния tibiale, intermedium и проксимального centrale (однако у Labidosaurus, intermedium не сростается с tibiale).
В семействе Captorhinidae череп удленен в предглазничной части. Представители данного семейства могут быть представленны следующими формами:
- мелкие, с однорядными челюстями (в отдельных случаях до 3х рядов (Татаринов, 2008)) - подсемейство Captorhininae
- крупные, с 5-10 рядами зубов в челюстях - подсемейство Moradisaurinae
-
Рис.1 Зубные кости представителей семейства Captorhinidaе

Рис.2 Представители семейства Captorhinidae (достигали от 40 до 80 cм в длинну)
_____ Яркие представители семейства Сaptorhinidae _____
- Подсемейство Сaptorhininae (Case, 1911)
Captorhinus aguti Cope,1882
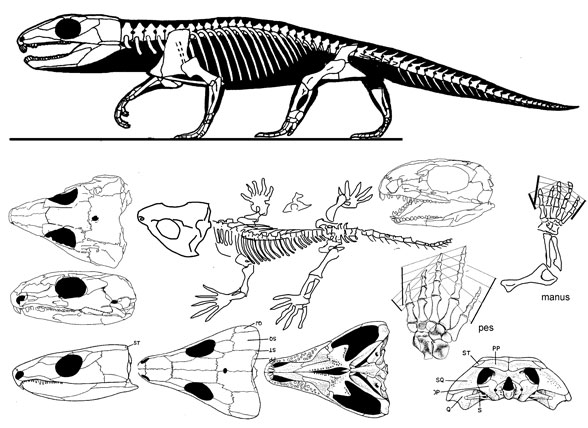

Naosaurus uralensis Riabinin, 1915

Имеют массивную, слабо выдвеннутую вперед и вниз, предчелюстную кость, которая несет не более трех зубов. На зубной кости - не более 15 зубов в одном ряду. Два передних укрупнены, со слабо уплощенными продольно коронками, остальные постепенно каудально уменьшаются в размерах, не меняясь в форме.
Riabininus uralensis Riabinin, 1915

Длина черепа не более 5 см. Челюстные зубы массивные, заостренные.
Ранняя пермь Восточной Европы. Типовое местонахождения; одновозрастное местонахождения Инта, Интский р-н, Россия.
- подсемейство Moradisaurinae (de Ricyles et Taquet, 1982)
Gecatogomphius kavejevi Vjushkov et Tchudinov, 1957

Средняя пермь Восточной Европы. В верхней челюсти 6 параллельных, не конвергирующих зубных рядов, в нижней - 5 передних конусовидных зубов, увеличивающихся в размерах, со слегка упрощенными продольными коронками, со слабо выраженными боковыми кантами и 5 не конвергирующих рядов притупленно-цилиндрических зубов, не менее 11 зубов в ряду.
Moradisaurus Taquet, 1969

Ранняя пермь Северной Америки. Крупные, с 5-10 рядами зубов в челюстях.
Представители отряда Bolosaurida имели короткий и высокий череп с нижним виисочным окном на стыке швов квадратно-скуловой, чешуйчатой и скуловой костей. Неполное костное небо может образовываться за счет широких фалангов небных костей. Зубы с выраженными птками. У представителей семейства Bolosauridae (Cope, 1878) верхнечелюстная кость довольно короткая ( не более трети длины черепа). Коронки зубов высокие, булавовидные. Не усложненные пятки выраженны хорошо, расположены косо субвертикально или почти субгоризонтально, у основания округлые или слабо овальные. От одного до трех последних челюстных зуба остроконические, так как размер к основанию уменьшается. Некоторые авторы (Иваненко, 2008) считают, что вполне могло присутствовать слабо выраженное вторичное небо. В отличаи от представителей предидущего семейства у таковых в семействе Belebyidae (Ivachenko, 2001) верхнечелюстная кость длинная (более половины длины черепа). Коронки зубов низкие, уплощенные, с хорошо выраженными широкими горизонтальными пятками, что поперчно сжаты у основания. В противовес представителям предидущего семейства, в данном случае размер кзади увеличивается, так, что последние зубы массивные, с хорошо выраженными горизонтальными пятками. Вторичное небо выраженно хорошо, так как небная кость почти наполовину прикрывает заднюю часть костной хоаны.

_____ Яркие представители семейства Bolosauridae _____
Семейство Bolosauridae (Cope, 1878)
Bolosaurus striatus Cope 1878
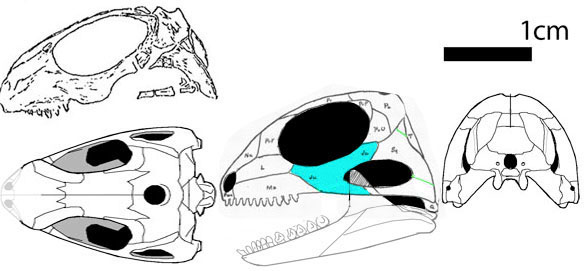 Ранняя пермь Северной Америки
Ранняя пермь Северной Америки
Stephanospondylus traati Tatarinov,1974
 Длина черепа не менее 7-8 см. В основании зубов заметны следы складчатости. Зубной ряд содержал 13-14 зубов, высота коронки которых чуть меньше высоты шейки (соотношение диаметра коронки среднего зуба максиллы к высоте - 1:3)
Длина черепа не менее 7-8 см. В основании зубов заметны следы складчатости. Зубной ряд содержал 13-14 зубов, высота коронки которых чуть меньше высоты шейки (соотношение диаметра коронки среднего зуба максиллы к высоте - 1:3)
Timanosaurus ivachnenkoi Gubin, 1993
Средняя пермь восточной Европы. Описан по отдельным позвонкам, схожим с позвонками болозавров в меньшей степени диадектид и парарептилий. Если смотреть на формальности, то от позвонков диадектид они отличаются отсутствием дополнительных сочлинений, от парарептилий - более высокими невральными дугами.
Семейство Belebyidae (Ivachenko, 2001)
Belebey vegrandis Ivachnenko, 1973
 Длина черепа более 6 см. убная кость массивная, коронки буккальных зубов слабо выдаются латерально. Два передних буккальных зуба небольшие, по форме идентичны последующим зубам. В верхнечелюстной кости передний зуб резцеобразный , по форме повторяющий резцы предчелюстной кости, после него следует небольшая диастема. Находки были обнаружены на территории России (Оренбургская обл., Александровский р-н.)(скелет без черепа) и Северного Китая (провинция Гансу) (челюстная кость).
Длина черепа более 6 см. убная кость массивная, коронки буккальных зубов слабо выдаются латерально. Два передних буккальных зуба небольшие, по форме идентичны последующим зубам. В верхнечелюстной кости передний зуб резцеобразный , по форме повторяющий резцы предчелюстной кости, после него следует небольшая диастема. Находки были обнаружены на территории России (Оренбургская обл., Александровский р-н.)(скелет без черепа) и Северного Китая (провинция Гансу) (челюстная кость).
Belebey maximi Tverdochlebova, 1987

Мелкая форма с длиной черепа не более 4 см. Зубная кость очень тонкая, коронки буккальных зубов сильно расширены и значительно выдаются латерально. Местоположение находок - территория России.
Davletkulia gigantea Ivachnenko, 1990
 Длина черепа не более 15-16 см. Коронки буккальных зубов овальные, сильно расширенные поперечно, поверхность коронки уплощенная, практически без выраженного режущего гребня (зубной аппарат перетирающего типа). Находки относятся к средней перми Восточной Европы.
Длина черепа не более 15-16 см. Коронки буккальных зубов овальные, сильно расширенные поперечно, поверхность коронки уплощенная, практически без выраженного режущего гребня (зубной аппарат перетирающего типа). Находки относятся к средней перми Восточной Европы.
Permotriturus herrei Tatarinov, 1968
 Находки относят к средней перми Восточной Европы. Длина черепа примерно 7 см. Имеют очень массивную зубную кость. Считают, что коронки буккальных зубов очень резко уплощены поперечно (судя по сохранившимся основаниям). Так как верхний край зубной кости массивен и сильно расширен, предпологают наличие давяще-перетирающих зубов (как у Davletkulia) однако с сильно сжатыми основаниями.
Находки относят к средней перми Восточной Европы. Длина черепа примерно 7 см. Имеют очень массивную зубную кость. Считают, что коронки буккальных зубов очень резко уплощены поперечно (судя по сохранившимся основаниям). Так как верхний край зубной кости массивен и сильно расширен, предпологают наличие давяще-перетирающих зубов (как у Davletkulia) однако с сильно сжатыми основаниями.
____ Список используемой литературы ____
-
М.Ф. Ивахненко, Е.Н. Курочкин (ответственные редакторы). Ископаемые позвоночные России и сопредельных стран. Ископаемые рептилии и птицы. Часть 1. Справочник для палеонтологов, биологов и геологов. - М.: ГЕОС, 2008. - 348 с.
-
Л.П. Татаринов. Очерки по эволюции рептилий. Архозавры и зверообразные. - М.: ГЕОС, 2009. - 377 с.
-
https://phys.org/news/2011-05-captorhinid-reptile-china.html