Губки (Porifera) – одна из древнейших ныне живущих групп животных, и, возможно, древнейших животных принадлежащих Metazoa. Высокий полиморфизм, вызванный тотипатентностью всех клеток губок, примитивное строение: отсутствие нервной, и, вероятно, мышечной ткани; отсутствие светочувствительных органов и клеток (помимо крестообразных клеток на некоторых стадиях эмбриогенеза); слабая специализация тканей, например, многие функции соединительной ткани у более сложных животных выполняют лишь несколько типов клеток мезоглеи – все это позволяет судить о примитивности данной группы. Но также губки обладают хорошо развитым коллагеновым матриксом, сократительной способностью (оскулярной и тотальной), сложными молекулярными комплексами рецепторов на поверхности клеток (тирозин-киназные, интегриновые), элементами сенсорной системы (метаботропические глутаминовые рецепторы, кристаллины), иммунной системой (иммунолгобулин-подобные молекулы, Rh система, цистеин-богатые клетки-утилиты), что классифицирует губок как истинных Metazoa [6].
Проблема «укоренения» всей группы многоклеточных животных является предметом дискуссий и последовательность ответвления от общей ветки многоклеточных губок (Porifera), гребневиков (Ctenophora), пластинчатых (Placozoa), стрекающих (Cnidaria) и билатерий (Bilateria) достоверно неизвестна, кроме того, что книдарии и билатерии являются ближайшими родственниками. Остальные группы являются самостоятельными и древними группами относительно всех многоклеточных.
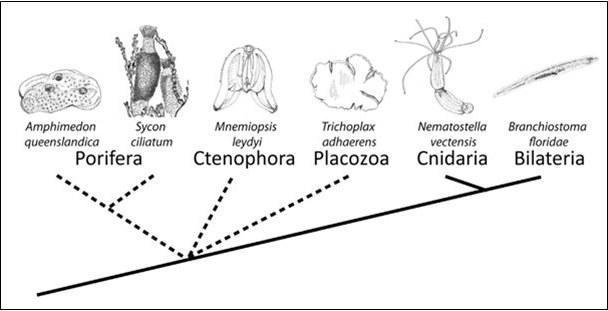
Рис. 1 Порядок ветвления имеющихся групп животных скорее не ясен (штриховкой обозначены группы с невыясненным филогенетическим положением) (Adamska, 2015).
Основная проблема состоит в том, что выяснение филогенетических связей ранних Metazoa и истории образования многоклеточных в целом усложняется по причине древности таких эволюционных событий. Что касается губок, то даже тут даже α-таксономия полностью не известна, также и филогенетические отношения внутри группы, таким образом, даже если мы принимаем гипотезу о том, что именно губки стоят у начала многоклеточности и дают начало другим группам, то на эту роль может претендовать и отдельный класс, например, Calcarea (что показано учеными из центра океанологии в Марселе). Также их данные свидетельствуют о том, что класс Hexactinellida у губок не только морфологически, физиологически но и филогенетически сильно отстоит от демоспонгий и известковых губок[2].
Исследования в области молекулярной филогенетики дают разные представления о том, какое место губки занимают на древе Metazoa, причиной этого является выборочность сравниваемых групп. Также существует большая зависимость топологии дерева от выбранной аутгруппы, которая будет укоренять дерево, а также от статистических моделей субституций сайтов в ДНК (по сути, от этого будет зависеть измерение скорости накопление мутаций и отображаться в силе дивергенции дерева а также на длине ветвей дерева).
Показано, что если мы будем выбирать более отстоящую от многоклеточных аутгруппу, например грибы (как часть монофилетической группы Opisthokonta), то возможно подтверждение альтернативной гипотезы о том, что Placozoa находятся в корне многоклеточности, в то время как выбор более близкородственной группы хоанофлагеллят (Choanoflagellata) дает хорошо поддерживаемую bootstrap методом топологию, подтверждающую гипотезу о происхождении всех многоклеточных животных от предка-губки [9]. Такой выбор может быть более целесообразным, так как хоанофлагеллят относят к близкостоящей относительно многоклеточных группе не только благодаря общим морфологическим признакам (например, часто сравниваются с хоаноцитами губок[4]), но и подтверждается филогенетическими методами[3,7].
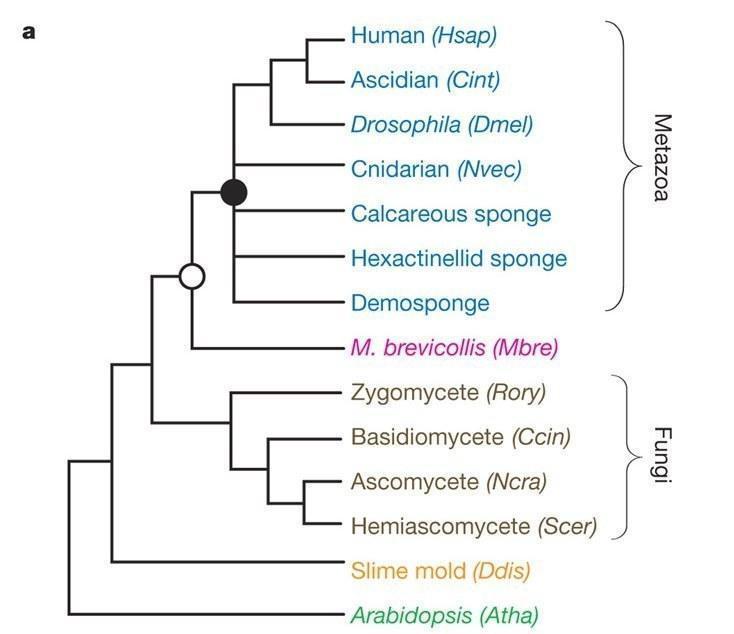
Рис. 2: Полногеномное исследование показывает близость хоанофлагеллят M. Brevicollis к многоклеточным относительно грибов. (King, 2008)
В целом же ряд молекулярных филогенетических исследований показывают, что именно губки стоят в основе Metazoa, при этом использовались различные методы поиска оптимальных топологий (например, максимальной парсимонии, neighbor-joining) а также с различные модели субституций нуклеотидов[2,9].
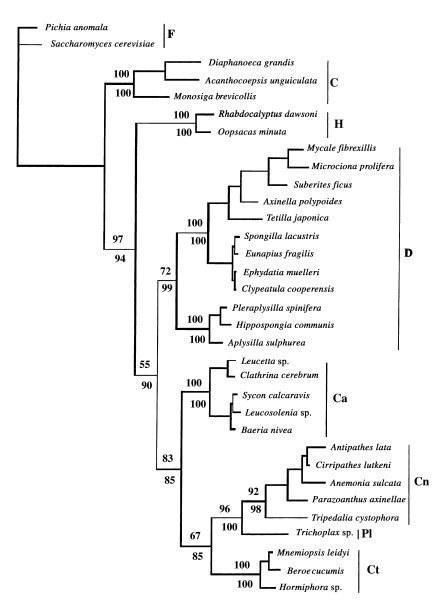
Рис. 3: Филогенетическое дерево, указывающее на базальное положение губок (H,D,Ca) относительно других многоклеточных: книдарий, пластинчатых, грибневиков (Cn,PI,Ct). Также видно, что известковые губки (Ca) объединяются с ними в одну кладу с высокой степенью поддержки (в статье даже предлагается выделить их в отдельный тип, дающий начало другим многоклеточным). Также подтверждается предположение близости хоанофлагеллят (С) к губкам и другим многоклеточным. (Borchiellini, 2001)
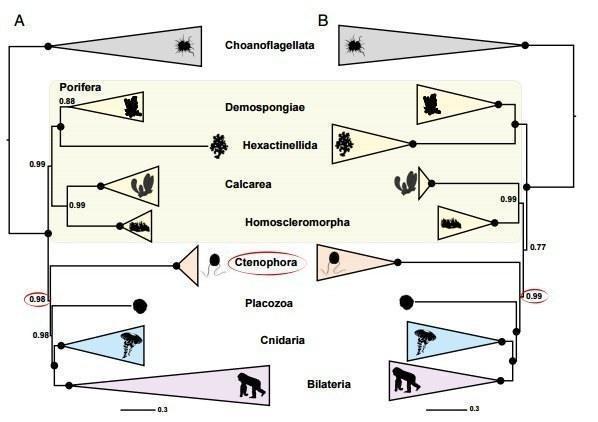
Рис.4: Филогенетическое дерево построенное с выбором хоанофлагеллят аутгруппой, используются сложные модели субституций нуклеотидов(А - CAT, В - CAT-GTR), с высоким уровнем поддрержки именно губки (Demospongiae, Hexactinellida, Calcarea, Homoscleromorpha) занимают базальное положение. Стоит отметить, что тут также изветковые губки вместе с другой групой гомосклероморф выделяются в отдельную "продвинутую" кладу. ( Pisani, 2015)
Касательно сравнительной физиологии: еще в конце 90-х учеными из университета в Ерфурте приводилось изучение некоторых рецепторных молекул, коллагена, белков иммунной системы губок и их ДНК последовательностей, используя модели субституций аминокислот также предполагает губок как самых древних представителей многоклеточных. При этом на основании анализа дается датирование расхождения губок и хоанофлагеллят, и эти оценки сходятся с другими методами определения времени появления губок (Ранний Кембрий). Что интересно, предполагается, что возможность многоклеточности , дифференциации клеток и последующий кембрийский взрыв в целом стал возможен после инициации эволюционно значимого механизма альтернативного сплайсинга (перетасовки экзонов в процессе транскрипции) 1000 млн лет назад. Также сравнение последовательностей, кодирующий белок-киназу С в этой работе указывает на Calcarea как близжайшего родственника Eumetazoa, и отделяет Hexactinellida от других губок[6]. Тут тоже можно указать на «конкуренцию» гипотез о первичности гребневиков и губок и тут также выигрывают губки, так как имеют еще более примитивную сократительную систему (мы знаем только о возможном участии в этом эндопинакоцитов[8]), нежели гребневики. Сравнение синапсов и нейромедиаторов гребневиков с таковыми у билатерий, правда, тоже выявляет сильные различия, но тут может играть роль конвергентная эволюция нервной системы [10].
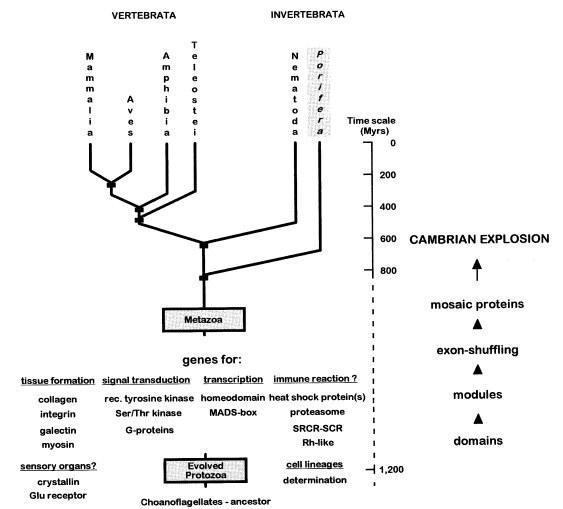
Рис. 5: Сравнительнй анализ белков (коллаген, интегрин, галектин, миозин), сигнальных молекул (тирозин-киназы, серин-тирозин киназы, G-белка), тринскрипционных факторов (гомеодомен, MADS--бокс), иммунорецепторов (Белки теплового шока, протеасомы, CRCR-SCR, Ph-подобные белки), клеточных линий и клеток-предендентов на сенсорные функции указывает также на базальное положение губок с временным датированием расхождения с хоанофлагеллятами (около 600 млн лет). В анализе не даются данные о гребневиках или пластинчатых. Также относительно временной шкалы набросаны этапы возникновения механизма, позволяющего значительно увеличить вариативность сложных белков (образование доменов, модулей, альтернативного сплайсинга). (Muller, 1998)
Но что если предположить существования предка-не губки, что имел еще более простое строение, при этом имел относительно дифференцированное многоклеточное тело? Это предположение действительно имеет основания существовать, но тут возникает ряд проблем, которые вызваны необходимостью палеонтологических данных, или невозможностью на данный момент такого предка смоделировать. Таким образом, этот вопрос можно оставить только открытым. А что если такой предок был сложнее?
Недавние исследования, опубликованные командой австралийских биологов, проделанные на базе Международного центра морской молекулярной биологии при Бергенском университете в области сравнения транскрипционных факторов эмбриогенеза трех классов губок, предполагают, что относительно примитивное строение тела губок может быть связано с утратой генов в следствии эволюции от более сложного предка, который дал начало губкам и кишечнополостным. Такой гипотетический общий предок должен иметь более богатый набор генов в сравниваемых генных семейств (в данном случае T-боксные гены, Sox-гены Гены класса ANTP, Six-гены, Pax-гены, GATA и Smad-гены), которые сохранились только частично свои для каждого класса губок. Филогенетический анализ также показал, что все гены этих семейств демонстрируют сходство с ближайшей группой – стрекающими. Также авторы указывают на то, что такая утрата генов происходила в ходе эволюции неоднократно у разных таксономических групп (например, те же кишечнополостные (Hydra), или даже позвоночные (Tunicata))[1]. Большим плюсом в поддержку гипотезы является то, что рассматриваемые гены играют иногда ключевую роль в эмбриогенезе губок (например, в ходе екскурвации кальциевых губок активно экспрессируется ген Brachyury, который, что интересно, принимает участие и в образовании осевого скелета у хордовых), таким образом, такие события как утрата генов этих семейств или дупликация будет действительно влиять на ход эволюции и являться информативными для выдвинутой гипотезы. В то же время это может говорить лишь о том, что многие гены просто менялись или модифицировались в геноме с ходом эволюции, или даже просто менялась функция гена, и это не является следствием утраты генов у более сложного предка. В любом случае, без палеонтологических данных это остается лишь гипотезой.
А что с палеонтологическими данными? Совсем свежее исследование 2016 года от ученых из Массатчусетского технологического университета не только подтверждает гипотезу о губках, как о самых древних многоклеточных, но и увеличивает их «древность» к докембрийскому периоду (около 640 млн лет). Нехватка традиционных ископаемых губок может быть восполнена обилием «молекулярных» ископаемых (минерализированных отложений). В данном исследовании это были отложения стерола 24-изопропилхолестана, для которого был установлен ген, отвечающий за выработку липида – SMT-ген. В дальнейшем использовались методы сравнительной геномики , с помощью которых были показали два возможных хозяина липида: водорослей и губки, при этом с помощью метода молекулярных часов (датирование эволюционных событий) было показано, что у губок он появился ранее а именно 640 лет назад[5].
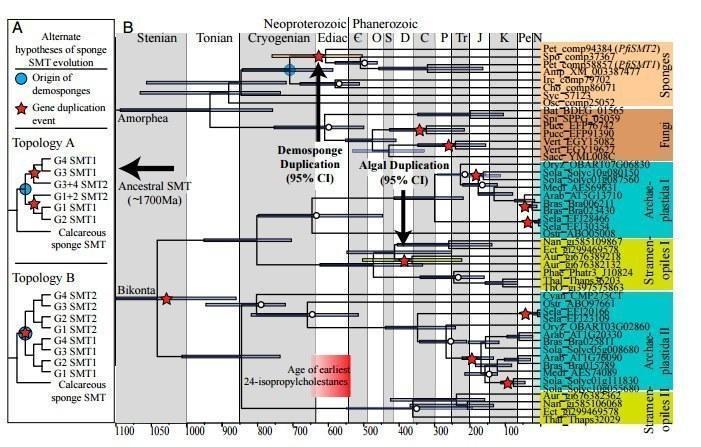
Рис. 6: А - альтернативные гипотезы эволюции SMT-гена, где звездочкой обозначен момент дупликации гена (образование SMT-гена, который ответственнен на синтез 24-изопропилхолестана) а синим кружком - момент происхождения кремниевых губок. В- датирование происхождение SMT-гена для губок (Demosponge Duplication) и водорослей (Algae Duplication). На схеме видно, что разница во времени для этих событий составляет до 300 млн лет. (Golda,2016)
Таким образом, скорее всего губки являются-таки самыми древними многоклеточными животными на Земле, но это порождает множество дополнительных вопросов. Например, являются ли они первыми представителями многоклеточных, или являются просто более успешной группой, дожившей до наших дней? Если являются, то какая была эта губка, так как мы имеем невероятное разнообразие губок, вплоть до синцитиальных форм (Стеклянные губки), и возможно даже есть основания выделять их в отдельные типы. Также принципиальным является вопрос об изучении их физиологии в рамках эволюции, так как, например, их сократительные способности могут сыграть большую роль в понимании того, как развивалась нервная система на начальных этапах. Открытым остается и вопрос, были ли именно губки предками книдарий и билатерий, или просто ближайшей сестринской группой дожившей до наших дней. Я думаю, что для этого еще долго необходимо собирать молекулярные, палеонтологические и физиологические данные по всем группам ранних многоклеточных, чтобы иметь возможность смоделировать гипотетического предка, что впервые позволил себе иметь множество специализированных клеток.
Источники:
- Sofia A. V. Fortunato, Marcin Adamski, Maja Adamska Comparative analyses of developmental transcription factor repertoires in sponges reveal unexpected complexity of the earliest animals // Marine Genomics. 2015 (in press)
- C. Borchiellini, M. Manuel, E. Alivon, N. Boury-Esnault, J. Vacelet and Y. Le Parco Sponge paraphyly and the origin of Metazoa// Journal of Evolutionary Biology. 2001. V. 14, Issue 1, P. 171–179.
- M. Carr, B. S. C. Leadbeater, R. Hassan, M. Nelson, and S. L. Baldauf Molecular phylogeny of choanoflagellates, the sister group to Metazoa// PNAS. 2008. V. 105(43). P. 16641–16646.
- James-Clark H. Conclusive proofs on the animality of the ciliate sponges, and their affinities with the Infusoria Flagellata. Am J Sci Ser. 2. 1866. V. 42 P. 320–325.
- David A. Golda, Jonathan Grabenstattera, Alex de Mendozab, Ana Riesgoc, Iñaki Ruiz-Trillob,d, and Roger E. Summons Sterol and genomic analyses validate the sponge biomarker hypothesis// PNAS V. 113. No. 10. P. 2684–2689.
- Muller WE Origin of Metazoa: Sponges as Living Fossils // Naturwissenschaften.1998. V. 85(1). P.11-25.
- Nicole King and oth. The genome of the choanoflagellate Monosiga brevicollis and the origin of metazoans// Nature. 2008. V. 451(7180). P. 783–788.
- Michael Nickel, Corina Scheer1, Jörg U. Hammel, Julia Herzen and Felix Beckmann The contractile sponge epithelium sensu lato – body contraction of the demosponge Tethya wilhelma is mediated by the pinacoderm// The Journal of Experimental Biology. 2011. V. 214. P. 1692-1698.
- Davide Pisani, Walker Pett, Martin Dohrmann, Roberto Feuda, Omar Rota-Stabelli, Hervé Philippe, Nicolas Lartillot and Gert Wörheide Genomic data do not support comb jellies as the sister group to all other animals// Proceedings of the National Academy of Sciences. 2015. V. 112. No 50. P. 15402–15407.
- Ястребов С. А. Сравнительная геномика вынуждает пересмотреть место гребневиков на эволюционном древе животных// Элементы, 2015.