Найважливіші події, що відбулися в біосфері з моменту появи скелета, досить чітко зафіксовані в палеонтологічному літописі. В результаті вивчення послідовності появи різних асоціацій скелетних організмів від моменту їх масового поширення до теперішнього часу була створена Геохронологічна шкала (рис. 1) відносного часу, яка широко використовується в
усьому світі.
Рис.1 Геохронологічна шкала
Сьогодні в результаті впровадження електронної мікроскопії та інших методів палеонтологія здатна вивчати те, що раніше здавалося абсолютно неможливо: скам'янілі залишки бактерій та інших організмів мікронного розміру, залишки ДНК і інших фрагментів органічних структур. Всі вони дозволяють будувати висновки про рівень організації тих організмів, які жили, але від яких нібито не залишилося нічого. Крім того, давні організми залишали свої сліди на породі. Вже в дофанерозойських породах були описані так звані строматоліти (рис.2). З'ясувалося, що строматоліт утворюється в результаті життєдіяльності абсолютно ні на що не схожої прокаріотичної спільноти, яка називається ціанобактеріальним матом. Мати існують у багатьох районах світу, головним чином в таких пересолених лагунах, як Сиваш, однак справжні строматоліти, як в ШаркБеї і на Багамскій мілини, вони утворюють досить рідко. Мат, що розташовується на верхній поверхні створюваний ним строматоліт, являє собою щільний багатошаровий "килим" загальною товщиною до 2 см; основу його складають нитчасті або пальмелоїдні ціанобактерії, однак крім них в формуванні спільноти бере участь безліч найрізноманітніших мікроорганізмів.

Рис. 2 Строматоліт
Австралійськими вченими в давніх породах були виявлені мінерали, ізотопний вік яких перевищує 4 млрд років. У карбонатних породах в строматолітах з віком близько 3,5 млрд років американськими і австралійскими вченими виявлено скам'янілі залишки організмів, надзвичайно подібними з ціанобактеріями. Можна з великою часткою ймовірності припускати існування вже в той час "прокаріотичної біосфери", тобто біосфери, насиченою примітивними спільнотами з без'ядерними клітинами. При цьому вкрай важливо, що для життєдіяльності ціанобактерій, як
правило, необхідний кисень. Отже, бактерії, яким не потрібен кисень, існували, ймовірно, раніше і відповідно неціанобактеріальна, але бактеріальна біосфера могла вже існувати.
Проте не можна виключити і позаземне походження життя. У всякому разі матеріали про знахідки в метеоритах органічних сполук або навіть закам’янілих примітивних організмів публікувалися неодноразово. Основна маса дослідників завжди сприймали ці публікації вельми скептично, але і сьогодні з'являються вельми цікаві матеріали, що не дозволяють відкинути позаземне походження життя. Група московських дослідників виявила в вуглистих хондритах (метеоритах) скам’янілі ціанобактерії і, можливо, недосконалі гриби, а американські фахівці знайшли сліди бактерій в уламках Марса. Перехід від переважаючої прокаріотної біосфери до еукаріотної зафіксувати надзвичайно важко. Підходи тут можуть бути різні, але ні один з них не дає однозначної відповіді.
Приблизно 20 років тому московським геологом А. Лейтесом в Удоканських відкладеннях Сибіру були виявлені трубкоподібні утворення, названі ним Udocania problematica, які з самого початку одні дослідники вважали залишками черв'яків або кишковопорожнинних, а інші жорстко встали на позицію їх неорганічного походження. Останні вважали, що ці трубки є псевдоморфізмами по кристалах якихось мінералів. Однак вивчення цього матеріалу за допомогою електронного мікроскопа, показало, що ми маємо справу не тільки з еукаріотами, але і з Metazoa, тобто багатоклітинними тваринами і, крім того, зі скелетом. Навіть якщо вважати, що вік порід з Udocanian не перевищує 1,4 млрд років, це погано узгоджується з попередніми висновками про час появи еукаріот. Треба зауважити, що в останні роки таких даних стає все більше і більше. Польські та російські дослідники виявили в метаморфічних породах фундаменту Східно- Європейської платформи з віком завідомо понад 2 млрд років не тільки нитки ціанобактерій, але і псевдоморфози по конідіям недосконалих грибів. Проте не менш цікаві результати аналізу слідів життєдіяльності організмів. Їх форма в ряді випадків може з переконливістю вказувати на їх надзвичайно складну багатоклітинну будову. Такі сліди були виявлені новосибірськими вченими в відкладеннях Ігаро-Норильському районі.
Відомі на весь світ вендські місця знаходження безскелетних Metazoa Європи, Австралії та Ньюфаундленду буквально кишать відбитками незвичайних тварин. Через першого відкритого і описаному місцезнаходженню в Едіакарії (Південна Австралія) цю асоціацію бесскелетних організмів стали називати едіакарською фауною. Пізніше, коли були відкриті ще багатші місцезнаходження в вендських відкладеннях Російської (Східно-Європейської) платформи, цю фауну нерідко називають вендо-едіакарською. Найбагатші місцезнаходження цих залишків виявлені також в Південній Африці, Сибіру, Поділлі і на узбережжі Білого моря. Кількість цих знахідок все зростає (рис. 3). За різними оцінками, 65-70% вендської біоти становлять кишковопорожнинні: близько 25% - черв'яки і близько 5% - членистоногі.

Рис.3 Біота вендського часу
Основною відмінною рисою вендо- едіакарської фауни є те, що вона представлена тільки відбитками бесскелетних організмів. І хоча деякі з цих тварин мають розмір до 1 м в довжину, вони, ймовірно, складалися з желеподібної речовини точно такої, як і сьогоднішні медузи. Дивує, звичайно, як же тоді вони могли зберегтися в похованнях в таких масових кількостях?
Серед вендо-едіакарскіх тварин були як бентосні (жили на дні, прикріплені або вільно лежачи), так і нектонні та планктонні форми, тобто плаваючі пасивно або активно в товщі води (рис. 4).

Рис.4 Реконструкція біоти раннього кембрію (по А. Журавльову і Л. Толпигіну)
З порід венда за допомогою хімічного препарування добувають величезну кількість мікроскопічних організмів і фрагментів організмів, що мали, ймовірно, пектинову оболонку. Завдяки пектиновій оболонці вони добре зберігаються у викопному стані і майже не руйнуються при хімічному впливі на породу. Ці утворення в більшості випадків є фітопланктоном. Фітопланктон венди виключно чисельний, хоча в основному це гладкі сферичні форми розміром 10-20 мк, але мають різноманітні особливості структури поверхні. На великих територіях вендського мілководдя виростали досить великі (до декількох сантиметрів в довжину) водорості - вендотеніди, дуже нагадують сучасні ламінарії. Збереглися у вигляді вуглефіцированних залишків (фітолейм). За формою і будовою талому поділяються на 5 порядків: округлі; корковидні; кущисті та шнуровидні; стрічкоподібні. Зв'язок вендотенідів з сучасними групами водоростей неясна.
На останньому етапі розвитку вендської біоти, спостерігається вельми своєрідна картина. Тут вже присутні кілька груп організмів, які будують мінеральну або органічну трубку. А в самих верхах з'являються примітивні молюски з колпачковидною раковиною. З немакіт-далдинського часу починається прогресивне збільшення форм зі скелетом (рис.5).
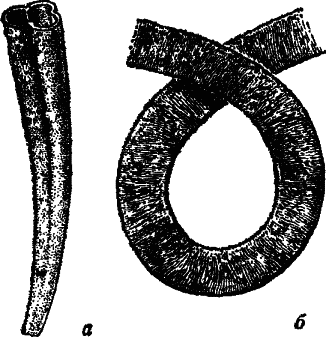
Рис. 5 Немакіт-далдинскі трубчасті копалини а - Anabarites, × 25, б - Sabellidites, × 16
ПОЧАТОК "СПІЛЬНОЇ СКЕЛЕТИЗАЦІЇ"
Рухаючись вгору по розрізу від докембрійських порід до кембрійських, виявляють, що порода насичена численними і різноманітними залишками організмів, вигляд котрих більш-менш звичний для нас. Тут починают зустрічати звичайні для всього фанерозою залишки губок, молюсків, брахіопод та інших організмів. Тільки незвичні їх розміри (перші міліметри). Всі відомі (виявлені) в кембрії, а точніше, в нижньому кембрії залишки організмів складають дві великі групи: 1) залишки організмів, систематичне положення яких ясно, оскільки є їхні молодші і навіть сучасні аналоги, і 2) залишки організмів, систематичне положення яких абсолютно неясно або вельми проблематично.
Серед одноклітинних тварин (Protozoa) відомі достовірні форамініфери. Перші вочевидь форамініфери мають дискоїдальну агглюттиновану раковину. Вони описані в атдабанськіх відкладеннях Прибалтики і називаються люкатієлами (Lukatiella). До форамініфер досить часто відносяться і трубки Platysolenites (рис. 6), перші представники якого знайдені в самих верхах докембрію.
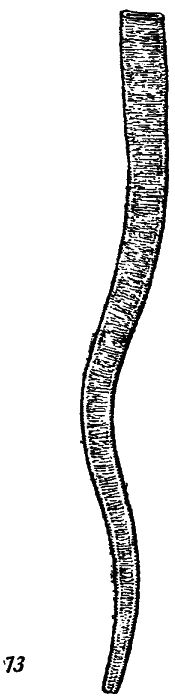
Рис.6 Platysolenites
Губки з крем'яними і карбонатними спікулами відомі з самого початку кембрію. Виключно рідко зустрічаються більш-менш цілі екземпляри. Майже завжди, коли знаходять залишки губок, це тільки скупчення розрізнених спікул. До викопних губок деякі дослідники відносять і археоціат (рис.7). Хоча вже давно вважається, що археоціати є самостійним типом тварин. Археоціати представляють собою в більшості випадків двостінні кубки різної форми. Зазвичай вони були прикріплені на дно, хоча могли бути і вільно лежачи. Діаметр кубків - від декількох міліметрів до декількох сантиметрів.

Рис. 7 Різноманітні форми кубків археоціат
Археоціати - перші з скелетних організмів - стали активними будівельниками рифогенних споруджень. Пізніше, їм на зміну прийшли корали. Інтерес до археоціат багатьох дослідників спочатку визначався тим, що це була друга після трилобітів група, що використовувалася для розчленування та кореляції кембрійських відкладень.
Кишковопорожнинні на початку кембрія представлені бідно. Однак все ж кілька груп було виявлено. Дуже широко представлені молюски. Але не зрозуміло, до яких класів молюсків повинні бути віднесені зі знайдених тут раковинок. І часто це дуже складне завдання, тому що саме тут ми стикаємося з предковими формами всіх молюсків, що несуть архаїчні признаки. Але в усякому разі ясно, що вже в ранньому кембрії зустрічаються достовірні моноплакофори. В останні десятиліття дослідження древніх представників молюсків призвело до думки про те, що в ті давні часи існував самостійний клас молюсків, названий Rostroconhia. Досить рано, майже з самого початку кембрію, зафіксовані і перші примітивні двустворчаті молюски. Вони зустрічаються не дуже часто, але їх знахідки відомі зараз практично по всьому світу. Вони були названі Fordilla. Широко в кембрії представлені так звані хіоліти. Вони мають зазвичай раковину у вигляді прямого або вигнутого ріжка з перетином різної форми. Раковина закривалася кришкою, форма якої відповідала формі перетину раковини.
Кембрій - це час бурхливого розвитку членистоногих, і особливо однієї з груп - трилобітів. Близько 50 сімейств цих тварин було поширено тільки в ранньому кембрії. Також з атдабанського часу відомі і найдавніші представники іншого класу членистоногих – Crustacea, її представники відомі з Сибіру, Китаю, Східно-Європейської платформи і Північної Америки.
З другої половини раннього кембрію починають освоювати великі простори брахіоподи, а до початку середнього кембрію стають другою за чисельністю групою серед копалин зі скелетом. І нарешті, ще одна група - це голкошкірі. Сучасні їхні нащадки - морські зірки, морські їжаки і т.д. Перші представники цієї групи з'являються кінця атдабанського століття і швидко розселяються по всьому світу. Скелети голкошкірих складаються з окремих табличок, які, в свою чергу, представлені монокристалами кальциту. Після смерті організма його скелет, як правило, розсипається, і в породі можна було знайти тільки розрізнені таблички. Але все ж вченим пощастило. У Північній Америці в ранньому кембрії були знайдені цілі скелети, і після цього стало легко виявляти присутність голкошкірих в породі по розрізненим мікрокристалічним табличкам. Всі перераховані вище залишки тварин належать до безхребетних. Однак є підозра, що найпримітивніші хребетні з'явилися вже в кембрії і, більш того, в ранньому кембрії. Таке уявлення базується на дослідженні так званих Hadimopanelа, які внутрішньою будовою більш всього нагадують луску найдавніших рибоподібних (рис. 8а). Так само як і раніше в венді, в морях кембрію, ймовірно, було дуже багато фітопланктону. Свідоцтво того - численні залишки сферичних гладких або шипуватих форм розміром зазвичай 10-20 мк, рідко до 100 мк, які виявляються у великій кількості в кембрійських породах. У карбонатних породах, як правило, зустрічаються вапновивідні водорості. Сьогодні дуже багато свідчень досить широкого розвитку ціанобактерій в кембрії. Особливо ефектно це було показано на прикладі вивчення фосфоритів і високовуглецевих сланців (рис. 9).
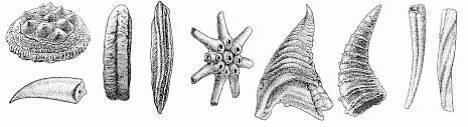
Рис. 8 Деякі копалини раннього кембрію: а - Hadimopanella, б - Sachites, в - Сіфогонухітіди, г- Ханцеллорія, д - Томмотііди, е – Anabarites
Дуже цікава і незвичайна група проблематичних залишків отримала назву томмотііди (рис.4,д). Вона була вперше описана з нижньокембрійських відкладень Сибірської платформи.
Рис.9 Ціанобактерії в фосфоритах нижнього кембрію
Останньою групою серед проблемних копалин, про яку необхідно сказати кілька слів, є так звані анабарітіди. Це трубчасті форми зазвичай довжиною в кілька міліметрів. Найбільш характерна особливість цих копалин - їх трипроменева симетрія (рис. 8 е). Трубки у них карбонатні. Вказується на ймовірність зв'язку анабарітідів з вендськими трипроменевими м'якотілими організмами. Переважна більшість перерахованих організмів жило в області мілководдя, в межах епіконтінентальних морів, тобто морів, розташованих на континентах.
Список літератури:
1. Розанов А.Ю. Что произошло 600 млн лет назад? М.: Наука, 1986.
2. Соколов Б.С ., Федонкин М.А . И еще 100 миллионов лет // Наука в СССР. 1983. № 5.
3. Томмотский ярус и проблема нижней границы кемб-рия / Под ред. М.Е. Раабен. М.: Наука, 1969.
4. Федонкин М.А . Бесскелетная фауна венда и ее место в эволюции Metazoa // Тр. ПИН. 1987. Т. 226.
5.Федонкин М.А. Бесскелетная фауна венда и ее место в эволюции Метазоа. М., 1987.
6. http://www.nestego.ru/2013/09/blog-post_9.html