
IV. Amphibia sensu lato-02. Origin of the larval-like body form and some features of Anura
The largest group of amphibians — the order Caudata — is characterized by a distinctive syndrome (a stable combination) of morphological traits. What determined this syndrome — the salamander-like body form?
Amphibian body form
As can be seen here, the amphibian group that dominates in species number is the tailless one. All adult tailless amphibians are easily distinguished from representatives of any other animal groups by their characteristic amphibian body form. Despite various life forms, all tailless amphibians retain the main traits of the amphibian body form: shortened trunk, absence of a tail, elongated hind limbs, and a broad flat head.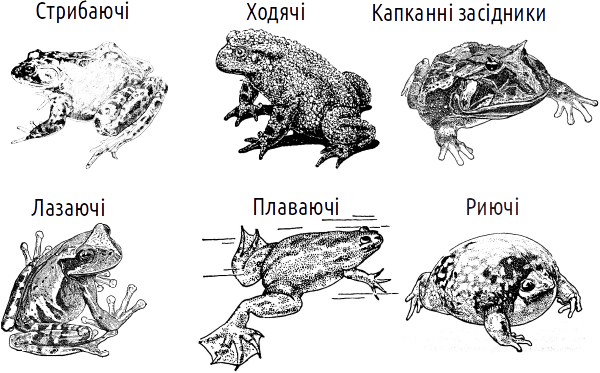
Owners of the amphibian body can move in various ways. For example, this video shows two different burrowing methods demonstrated by the pig frog (family Hemisotidae) and the Myobatrachidae.
Tailless amphibians can climb trees exceptionally well, as this brightly‑eyed tree frog (family Phyllomedusidae) shown in the second half of the following video (after another tree frog).
But which mode of locomotion was primary for amphibian animals?
One of the most significant transitional forms in biology is the Triassic Triadobatrachus – a member of the order Primitive tailless amphibians. Look at the reconstruction of its skeleton – all the features of the tailless skeleton are indicated, yet these changes are not fully completed! The body shortens, ribs are reduced, the pelvis elongates, a new, still small segment appears in the hind limbs, the skull lightens, the tail is very short… All this is an adaptation for jumping.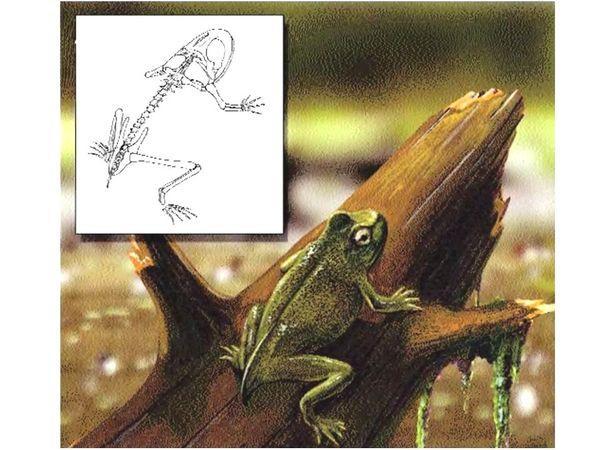
Another reconstruction of Triadobatrachus. It is hard for us to imagine it because we compare it with our ideas of modern tailless amphibians, and it seems somehow "unfinished". Of course, each species exists outside any calculations for a distant evolutionary perspective. Triadobatrachus did not stop halfway to modern amphibians; it simply lived in the conditions characteristic of its time. Yet after its existence, the same selective vector that shaped this species continued to act on its relatives and produced modern tailless amphibians.
The key reason that shaped the amphibian body form is adaptation to jumping. For amphibians (a group of animals whose majority have an aquatic development and a definitive terrestrial adult stage) it is natural to stay near the water’s edge. Among the most important tasks that all animals solve are not only obtaining food but also escaping predators. A common escape method is flight, which can be achieved by various locomotor strategies. Near the water’s edge there is the possibility to quickly enter the water, out of reach of terrestrial predators. The fastest way to do this is with a single jump.
Such a jump must be powered by strong limbs that push the body off a substrate. The effort of the limbs must be transmitted to a rigid axial skeleton that moves the whole body. Recall how the adaptation to jumping altered the body of the early jumper, the *Kakops* (one of the first water‑jumpers). The evolutionary line that gave rise to modern tailless amphibians adapted to jumping for much longer than *Kakops* and became far more transformed by this adaptation. *Kakops* can be viewed as a non‑adaptive water jumper, whereas tailless amphibians are adaptive jumpers.
The longer the push off the substrate, the greater the acceleration of the jumping body. Look at the photograph: the frog’s body is already in the air, and the legs continue to push it. Elongation of the hind limbs leads to the formation of a secondary tibia from the pre‑tarsus – another limb segment. The large and small tibial bones fuse, and the secondary tibia consists of two bones that allow rotation at the joint formed within the hand.
The longest frog toe will push its body off the substrate literally to the last moment.
This is how it happens.
The vertebral column of tailless amphibians is sharply shortened (typically to 9 vertebrae). The tail disappears (nothing to wobble during a jump!), the caudal vertebrae become a rod located inside the body contour. The body will remain stable in flight if the force is transmitted to the axial skeleton forward of the center of mass. If the thrust is applied behind the center of mass and not exactly in line with its direction (upward, for example), the body may simply flip in flight! Hence the pelvic girdle is strongly elongated. The sacral vertebra that receives the thrust from the hind limbs is positioned anterior to the body’s center of mass. Lightening of the head and the anterior body also contributes to this effect.
In a jump it is important not only to jump well but also to land successfully (the jump is not always directed into water). Probably, rib reduction is related to preventing internal organ damage upon landing. The forelimbs act as shock absorbers. Note that they are not connected to the axial skeleton by any bones: the pectoral girdle simply surrounds the vertebral column.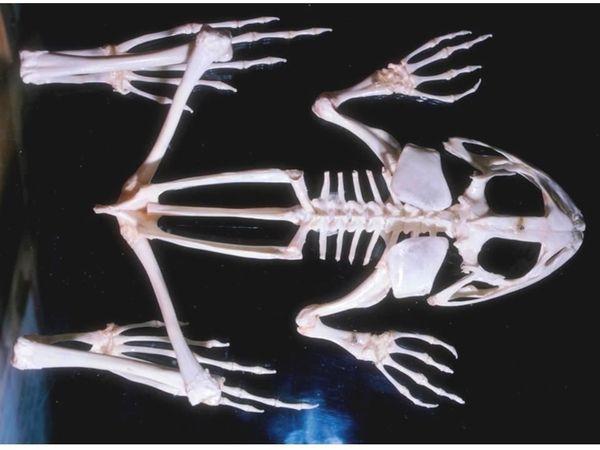
In the skeleton shown above the forelimbs are positioned incorrectly: the hands point forward. In most tailless amphibians, except the most primitive, the hands are pronated (turned inward). Below is shown how they are positioned in a living frog. With the limbs straight, the frog’s elbows would strike the abdomen on landing; with pronated hands they allow a soft landing by splaying the elbows outward and dissipating jump energy.
Do you see, by the way, where this frog’s pelvis ends? Another error in the assembled photographed skeleton is that the angle between the pelvic girdle and the sacral vertebrae processes is not shown.
A predator, whose prey could escape with a single jump, would approach it. Thus, a water jumper must have well‑developed vision and hearing.
If hearing is well developed, complex acoustic signaling becomes possible. By the way, even a splash from a single frog that jumps into water alerts all others. However, the greatest complexity of acoustic signaling is reached in mating vocalizations.
Amphibian development is not particularly stable. Studies show that a significant portion of individuals in many populations have various morphological anomalies. Often these anomalies are used to assess the "health" (quality) of the environment. The causes of anomalies are not always linked to external adverse factors. Interestingly, in some cases characteristic anomalies result from parasite influence; in other cases the same anomalies may arise "by themselves" due to developmental instability, the cause of which remains unknown.
Overall, tailless amphibians (as amphibians in general) are one of the most vulnerable groups. This has several reasons. One is that most amphibians require two habitats: terrestrial and aquatic, as well as safe corridors between them. In the modern fragmented landscape, the probability that at least one of the two required habitats for tailless amphibians will be destroyed is higher than the probability of destroying the single habitat of many other settled animal species.
Globally, the sharp decline in amphibian numbers is linked to the spread of infections. A particular danger is chytridiomycosis caused by *Batrachochytrium dendrobatidis*. Some groups of tailless amphibians, such as members of the genus *Atelopus* (harlequin toads, family Bufonidae), have suffered catastrophic impacts from this chytrid fungus.
As far as the author knows, data on *Batrachochytrium dendrobatidis* findings in Ukraine are fragmentary. The author of this online textbook found in 2006 a male of the sharp‑marbled frog suffering from an unknown disease at the Iskovy Reservoir near the Biological Station of V.N. Karazin KhNU. Shortly after capture the frog died. Its morphology was examined by a qualified mycologist (O.Yu. Akulov), who determined that the skin lesions fully corresponded to the impact of *Batrachochytrium dendrobatidis*; unfortunately, molecular analysis of this sample was not performed.
Anurans can climb trees superbly, as shown by this colorful tree frog (family Phyllomedusidae) in the second half of the following video (after some other tree frog). But which mode of locomotion was primary for frog-like animals? One of the most significant transitional forms in biology is the Triassic Triadobatrachus, a representative of the order Proanura. Look at the reconstruction of its skeleton – it shows all the features of the anuran skeleton, but these changes are not yet complete! The body is shortening, ribs are reducing, the pelvis is elongating, a new, still small, segment appears in the hind limbs, the skull is lightening, the tail is greatly shortened... These are all adaptations for jumping locomotion. Another reconstruction of Triadobatrachus. It is difficult for us to imagine it because we compare it with our ideas about modern anurans, and it seems somewhat "unfinished" to us. Of course, each species exists outside any calculations for a distant evolutionary perspective. Triadobatrachus did not stop halfway to modern amphibians; it simply lived in the conditions characteristic of its time. But even after its existence, the same selection vector that shaped this species continued to act on its relatives and created modern anurans. The key reason that shaped the frog-like body shape is adaptation to jumping. For amphibians (a group of animals for most of which aquatic development and emergence onto land in a definitive state are characteristic), it is natural to be near the water's edge. Among the most important tasks that all animals solve is not only obtaining food but also escaping predators. A common method of escape is flight. It can be achieved through various means of locomotion. Near the water's edge, there is an opportunity to quickly enter the water from the sphere of accessibility of terrestrial predators. This can be done most quickly with a single jump. Such a jump must be supported by strong limbs that push the body off the support. The force of the limbs must be transmitted to a rigid axial skeleton, which sets the entire body in motion. Recall how adaptations for jumping changed the body of Kakops, one of the first to jump into the water. The evolutionary line that gave rise to modern anurans adapted to jumping much longer than Kakops, and was much more profoundly transformed by this adaptation. Kakops can be considered an inadaptive jumper into water, while anurans are evadaptive jumpers. The longer the push-off from the support continues, the faster the jumping body can accelerate. Look at the photograph: the frog's body is already in the air, and its legs continue to push it. The elongation of the hind limbs contributes to the formation of a secondary tibia from the tarsus – another segment of the limb. The tibia and fibula fuse, and the secondary tibia consists of two bones, which provide the possibility of rotation in the joint formed within the wrist. The longest toe of the frog will push its body off the support literally until the last moment. This is how it happens. The spine of anurans is sharply shortened (typically up to 9 vertebrae). The tail has disappeared (nothing to dangle during a jump!), the caudal vertebrae have turned into a urostyle, located within the body contour. The body will be stable in flight if the force is transmitted to the axial skeleton forward of the center of gravity. If the push is applied behind the center of gravity and slightly off to the side of the direction to it (upward, for example), the body may simply flip over in flight! Therefore, the pelvic girdle is greatly elongated. The sacral vertebra, to which the propulsive force of the hind legs is transmitted, is located in front of the body's center of gravity. The lightening of the head and, in general, the anterior part of the body contributes to this effect. In a jump, it is important not only to jump well but also to land successfully (the jump is not always directed into the water). Apparently, the reduction of ribs is related to the fact that they would damage internal organs upon landing. The forelimbs act as shock absorbers. Note that they are not connected to the axial skeleton by any bones: the shoulder girdle simply encircles the spine. In the skeleton shown above, the forelimbs are positioned incorrectly: the wrists are directed forward. In fact, in most anurans, except for the most primitive ones, the wrists are pronated (turned inward). Below is shown how they are positioned in a live frog. With the limbs in a straight position, the frog's elbows would press against the abdomen upon landing; with pronation, they allow for a soft push-off by spreading the elbows to the sides and absorbing the jump's energy. Notice, by the way, where the pelvis ends in this frog? Another error in assembling the skeleton photographed above is that the angle between the pelvic girdle and the sacral vertebra processes is not shown. A predator whose prey can escape it with a single jump will stalk it. Therefore, a water-jumper must have well-developed sight and hearing. If hearing is well-developed, there is a possibility for fairly complex acoustic signaling. By the way, even the splash of one frog jumping into the water is a reason for all others to be alert. But, of course, acoustic signaling reaches its greatest complexity in mating vocalizations.
Amphibian development is not particularly stable. Studies show that a significant portion of individuals in many populations have various morphological anomalies. Often these anomalies are used to assess the "health" (quality) of the environment. The causes of anomalies are not always linked to external adverse factors. Interestingly, in some cases characteristic anomalies result from parasite influence; in other cases the same anomalies may arise "by themselves" due to developmental instability, the cause of which remains unknown.
Overall, tailless amphibians (as amphibians in general) are one of the most vulnerable groups. This has several reasons. One is that most amphibians require two habitats: terrestrial and aquatic, as well as safe corridors between them. In the modern fragmented landscape, the probability that at least one of the two required habitats for tailless amphibians will be destroyed is higher than the probability of destroying the single habitat of many other settled animal species.
Globally, the sharp decline in amphibian numbers is linked to the spread of infections. A particular danger is chytridiomycosis caused by *Batrachochytrium dendrobatidis*. Some groups of tailless amphibians, such as members of the genus *Atelopus* (harlequin toads, family Bufonidae), have suffered catastrophic impacts from this chytrid fungus.
As far as the author knows, data on *Batrachochytrium dendrobatidis* findings in Ukraine are fragmentary. The author of this online textbook found in 2006 a male of the sharp‑marbled frog suffering from an unknown disease at the Iskovy Reservoir near the Biological Station of V.N. Karazin KhNU. Shortly after capture the frog died. Its morphology was examined by a qualified mycologist (O.Yu. Akulov), who determined that the skin lesions fully corresponded to the impact of *Batrachochytrium dendrobatidis*; unfortunately, molecular analysis of this sample was not performed.