SexOnR–08_Sex. "Sex – the queen of problems in evolutionary biology" (Problem description II)
«Sex is the main challenge of modern evolutionary theory... the queen of problems of evolutionary biology», said American biologist Graham Bell. Why did he think so? Why does it make sense to agree with him?
"Sex – the queen of problems in evolutionary biology"
"Sex — the main challenge of modern evolutionary theory... the queen of problems in evolutionary biology," said American biologist Greg Bell. Why did he think so? Why is there any reason to agree with him?
1. The Maynard Smith paradox (the fact that sexual reproduction in populations is somehow not displaced by clonal reproduction) can be explained:
— either by the presence of an internal safeguard against the loss of sexual reproduction;
— or by a rapid “environmental penalty” for switching to clonal reproduction.
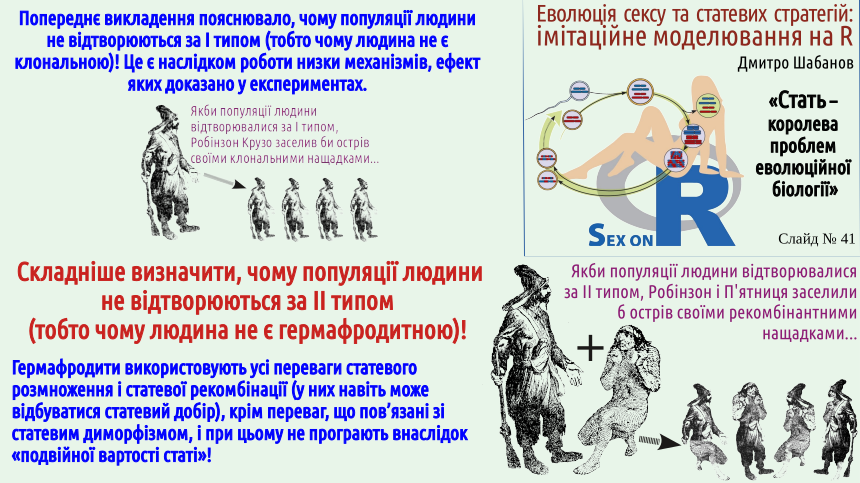
At the end of the previous lecture the Maynard Smith paradox was explained. On the one hand, we know many cases of the appearance of clonal (e.g., parthenogenetic) females. On the other hand, we observe the stable existence of bisexual populations that, apparently, should be very fragile and unstable to the emergence of clonal individuals. Why does this happen?
It should be noted immediately that parthenogenesis and other forms of clonal reproduction are not known in all animal groups. For example, parthenogenetic mammals are unknown; presumably, the regulatory mechanisms of mammalian development are more complex than those of other groups. It seems that only one case of avian parthenogenesis is known (in ostriches). And, suppose, in reptiles, amphibians and fishes many cases of clonal reproduction are known (and we considered it using the example of reptiles: the Darevskian lizards). Among the numerous invertebrate groups many cases of clonal reproduction are known; later in this lecture we mention a fairly typical case of Daphnia, which alternate parthenogenetic and sexual reproduction (and thus in the proposed classification of reproductive types should be placed on some bridge between types I and III — how to show this on a diagram is not very clear).
The slide states that in humans parthenogenetic females could appear. Most likely this is impossible because humans are mammals. By the way, at one time the possibility of human parthenogenesis was seriously considered as a scientific explanation of the immaculate conception of the Virgin Mary. This is a good example of how such attempts to “sit on two chairs” become untenable. From a scientific point of view this explanation is unacceptable. Moreover, in species with an XY sex‑determination system (which is exactly the case for humans), the offspring of parthenogenetic females would have to consist exclusively of females. Where could a Y chromosome arise in a female egg? This idea is unacceptable also from the standpoint of faith. For believers in the evangelical narrative, immaculate conception is a miracle, a violation of natural laws, not an interesting embryological anomaly. Note: in this paragraph we do not accept or refute the beliefs of conscientious believers; we merely emphasize that our discussion and faith lie on different planes, and attempts to combine them are counterproductive.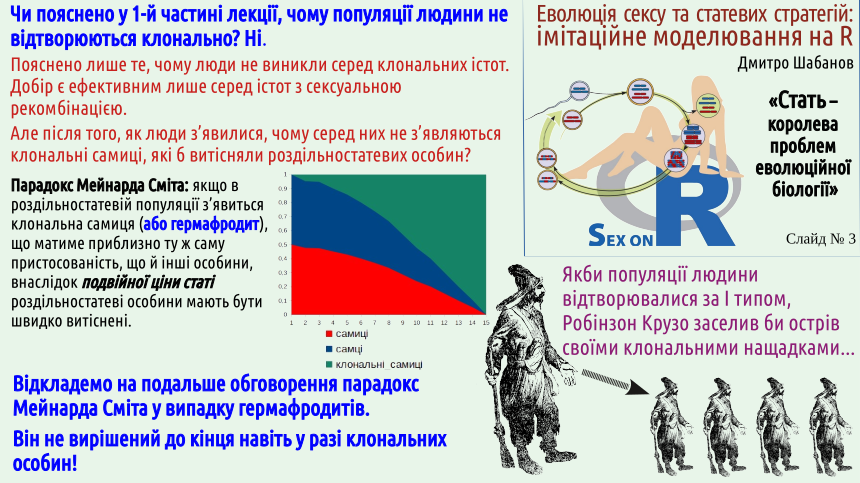
The next slide also reminds of what was discussed in the previous lecture.
In the author's view, the fact that in many species' populations the transition to parthenogenetic reproduction does not occur, as the Maynard Smith paradox would predict, can be explained in two different ways.
— First: in the developmental regulation system of many species there exists some mechanism (let us call it a safeguard against loss of sexuality). In this case a complex question arises: how could such a safeguard have originated? What selection could have shaped it? Recall two theses from the previous lecture: the first, that usefulness is created by selection, and the fifth, that there is no foresight in evolution.
— Second: populations that could switch to clonal reproduction are prevented from doing so by some external mechanism (let us call it a rapid environmental penalty for clonality). The long‑term penalty for clonality is known to us: Müller’s ratchet, but the accumulation of deleterious consequences of clonality requires a much longer time than the rise of clonal reproduction. Again we recall the thesis about the absence of foresight in evolution.
This choice can be phrased differently. Do the populations themselves not switch to clonal reproduction, or does the environment prevent it? The answer is not obvious. Spontaneous transitions to clonal reproduction are known in many species (not among mammals, but, say, among reptiles – fairly often). There are populations that easily alternate sexual and clonal reproduction. Williams’s balance argument expects a certain environmental influence.
It may also be that it makes no sense to ask the question in this way: internal OR external mechanisms maintain sexual reproduction. Perhaps both internal and external mechanisms operate.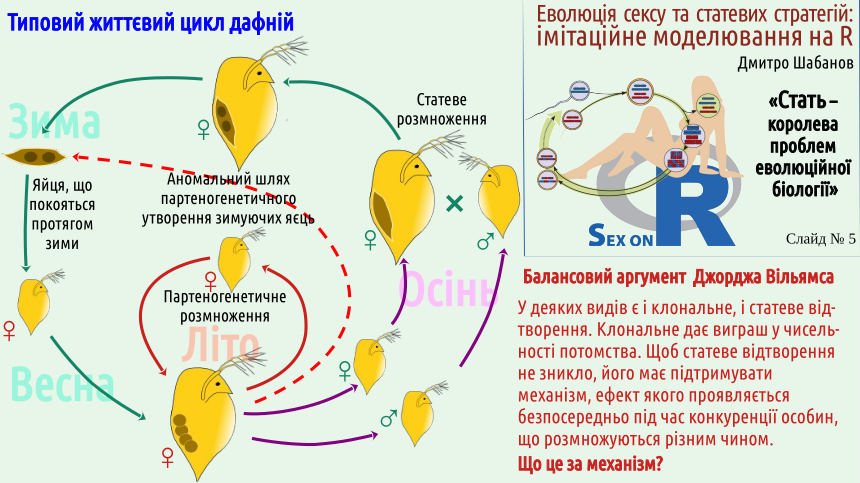
Brilliant researchers have proposed a series of hypotheses that explain the maintenance of sexual reproduction. We will list a certain set and then examine them in more detail. By the way, even here different mechanisms may act simultaneously.
2. Biological systems are hierarchical; theoretically selection can act at different levels of the biosystem hierarchy.
When characterizing the listed hypotheses that attempt to explain the mechanism maintaining sexual reproduction, we will identify which level of selection creates and sustains these mechanisms. For this we need to deal with the problem of selection levels.
The slide shows one version of the hierarchy of organization levels of biosystems (details – here). It should be emphasized that there is no universal set of levels; in different cases the number of levels at which new qualities arise through the integration of subsystems and their interactions may differ. The basis for treating a biosystem as a distinct level of organization is precisely the emergence of properties that are absent in its separate parts. Since a population can be immortal whereas an organism cannot, we have every reason to consider the population level as a separate level.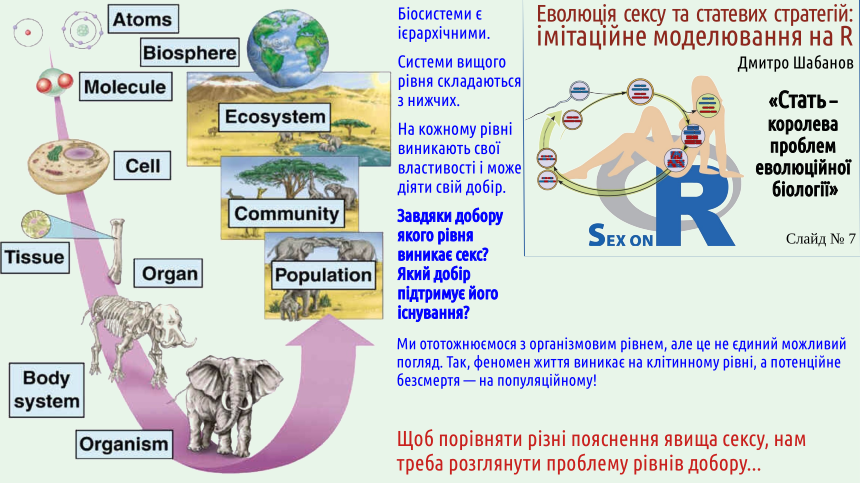
3. Various authors have considered selection at least at the gene, cellular, organismal, group (population), and species levels.
At which level does natural selection act? Classical British evolutionism was confident in the answer: the organismal level, because organisms die and survive, leave offspring or are excluded from reproduction. Darwin (who was known for penetrating conjectures) entertained, when discussing human evolution, the possibility of group selection.
Weismann realized that the history of organisms can also be examined at the cellular level. Most cells of an organism are mortal; potential immortality, the passage from generation to generation, is characteristic of cells of the germ line. We have already discussed the difference between soma and germ (germ line). Selection can be considered as something that also operates at the cellular level.
By the way, suppose that during fertilization in many animal species it is impossible not to recognize selection at the cellular level: among many sperm, only one fertilizes the egg. If this is a random sperm, there is no selection, only a reduction in number (since selection is success or failure depending on the system that is the object of selection). If the success of the sperm depends on its properties (and most likely it does), we observe an example of selection at the cellular level.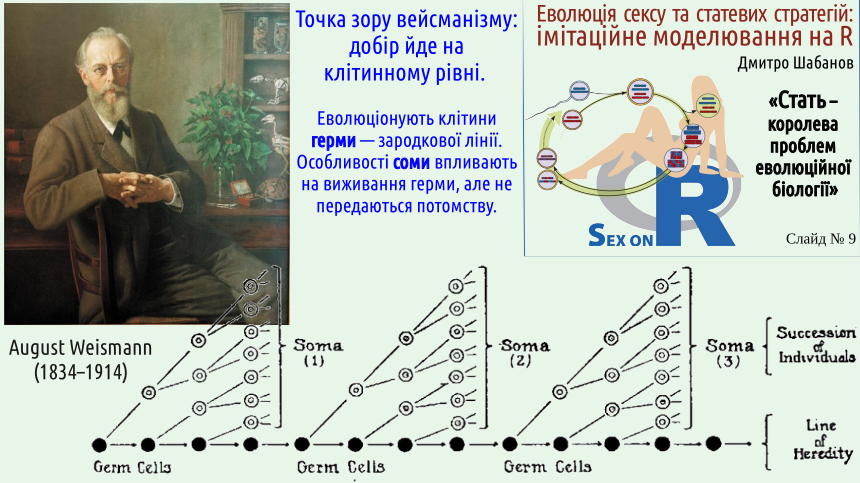
In the 20th century a revolution occurred. It was started by Haldane and made a fact of public consciousness by Dawkins. In the next generation it is not organisms but their genes that pass on. In the gene pool of a population these genes pass from individual to individual, form different combinations, and change over time. Thus selection can be considered at the gene level.
Dawkins’s “The Selfish Gene” became a book whose popularity rivals Darwin’s “On the Origin of Species”. Many are convinced that in this book Dawkins proved that selection operates exclusively at the gene level. In fact even Dawkins himself did not state this categorically. In “The Extended Phenotype” – another of his books, largely devoted to the topic of selection levels – the viewpoint is much more moderate and reasoned than in “The Selfish Gene” (and therefore it received far less popular attention). The “Extended Phenotype” perspective is: selection could also act at other levels, for example at the group level, but group selection would be ineffective compared with gene selection, and therefore the focus should be on understanding selection at the gene level.
The idea of group selection became intensively discussed as a result of Vero Wynne‑Edwards’s ideas. He considered such selection an important factor directing evolution.
Wynne‑Edwards’s arguments were critiqued by George Williams (the same one who proposed the balance argument). Let us consider (perhaps we should consider not only at the discussion level but also at the level of simulation modeling) a typical situation.
When a certain population experiences famine, many organisms curb their reproduction. Suppose that in years with few mice, specialized predators that feed on mice may curb their reproduction. Vipers may skip breeding or produce fewer offspring; owls may lay one clutch instead of two, and the number of eggs (and chicks) in each clutch will be smaller than in a “mouse year”. Why?
Wynne‑Edwards’s version: individuals sense that a hard year lies ahead for the population and restrain reproduction to reduce competition and mortality.
Williams’s version: those individuals that do not spend resources on reproduction (which, due to high mortality, is likely to be ineffective) will have a higher chance of surviving and waiting for better times. Even under conditions of high mortality, individuals that produce more offspring will have higher chances of their offspring surviving. If an individual contributes more to the future of the population, it will be favored by selection, even when excessive reproduction harms the population level.
The discussion of selection levels became even more complicated after brilliant American paleontologists suggested that the unit of selection could also be whole species.
Is the list of levels we have considered complete? In fact, no. For example, one could mention the concept of ecosystem evolution, which focuses on the stability or instability of ecosystems (or assemblages) as a whole.
Nevertheless, in the 20th century the most popular ideas held that evolution should be explained by selection at the gene level. It seemed that one of the most authoritative biologists, Edward Wilson, was one of the architects of this view. Moreover, Wilson created sociobiology – the science of the evolution of social behavior as a result of gene selection.
4. The concept of multilevel selection appears convincing, according to which selection can act on several levels simultaneously.
David Wilson (a namesake of Edward Wilson) and Eliot Sobel at the end of the 20th century put forward a very simple idea: selection works simultaneously on several nested levels, like Russian matryoshkas.
A scandal related to the problem of selection levels was triggered by a change of viewpoint of the patriarch of American evolutionary biology Edward Wilson: he accepted the multilevel selection concept. Together with young mathematicians he engaged in simulation modeling and recognized the important role of group selection. A sharp debate began between Wilson and Dawkins, in which, ultimately, Dawkins began to deny the very possibility of group selection, and Wilson denied the possibility of gene‑level selection. It seems both positions are skewed.
There are many documented examples of “selfish genes”, for instance those that intervene in meiosis and ensure their preferential transmission to the next generations.
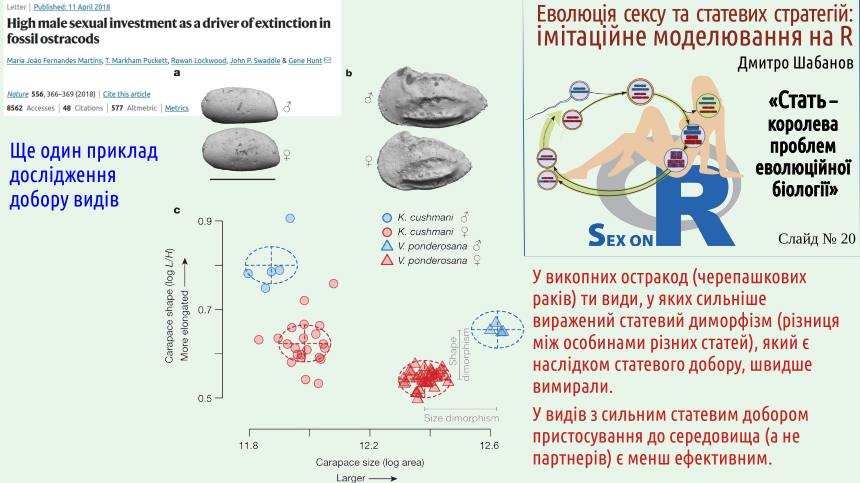
There are also well‑documented examples of selection at higher levels, including at the species level.
In the discussed example we see how selection at the species level influences the evolution of sexual reproduction in plants. The advantage of such studies is that modern molecular‑genetic methods allow precise reconstruction of how the evolutionary tree branched during the formation of contemporary species.
In the author's view there are ample grounds to adopt the multilevel selection concept. Thus we assume that the traits we are interested in may be influenced by selection operating at different levels. This makes it possible to analyze at which specific level selection responsible for particular evolutionary changes is acting.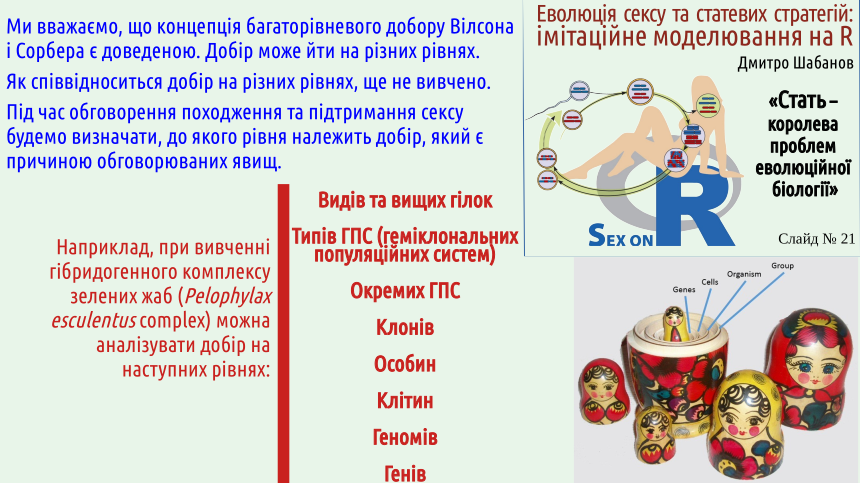
5. A set of hypotheses explaining the advantages of sexual reproduction is proposed. Some of these hypotheses have received experimental confirmation. Special attention is due to “Kondrashov’s axe”, Menning’s sexual selection hypothesis, and Van Valen’s “Red Queen” hypothesis.
Now we can move to discussing the hypotheses that explain the stability of separate sexes (their list is on slide 6). The first is the “branch selection” hypothesis of Hanne.
In fact, this is an attempt to explain separate sexes as a consequence of species selection. There is no reason to dismiss this hypothesis as impossible, but all the processes we observe cannot be explained by it alone.
In the Komodo monitor, parthenogenetic females sometimes appear. This is a representative of a fairly ancient evolutionary branch. We will assume that such monitors existed – existed, and now have reached the brink of extinction due to the appearance of parthenogenetic individuals? And why do mayflies and daphnia feel so well?
“Kondrashov’s axe” works under a certain pattern of interaction of deleterious alleles.
The next two slides illustrate Kondrashov’s hypothesis. We consider offspring from parents, each carrying 5 harmful alleles. In the offspring the number of harmful alleles is determined combinatorially; of course it can range from 0 to 10; the majority will have 5 defects. We consider a situation where the presence of each defective allele reduces the viability of the individual.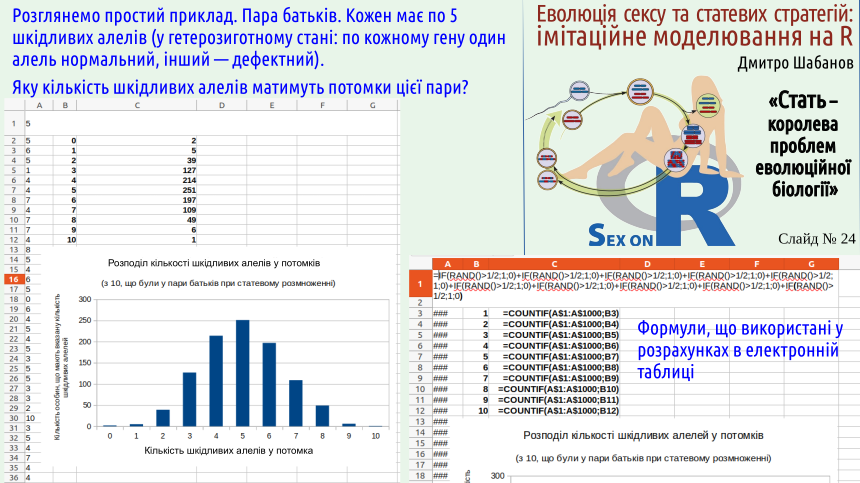
How much worse is it to have 2 defective alleles than 1? Different scenarios are possible. Perhaps having two defects is twice as bad as having one. This case would be observed if these genetic defects and their effects do not interact. This variant is shown on the slide in green. In it an individual with one defect has a mortality probability of 0.04, and with two defects – 0.08.
It may also be that having two defects is more than twice as bad as having one. This variant is shown in red‑violet, “beetroot” colour. A carrier of two defects has a mortality probability of 0.088. This corresponds to a genetic interaction called positive epistasis (mutual reinforcement of gene effects).
In raspberry colour the case is shown where harmful alleles significantly amplify each other’s action. Having two defects is three times worse than having one: mortality probability equals 0.12. Individuals with 7 or more defects become non‑viable under such conditions. On the distribution by number of genetic defects this part is seemingly cut off by the axe. These overloaded individuals can be compared to “cuckoo‑parasitic” birds that carry the “sins” of the whole population.
Why does this mechanism of purging genetic defects by selection outperform what can be observed in organisms with clonal reproduction? Of course, positive epistasis would also work in clonal organisms. But in clonal populations “cuckoo‑parasitic” individuals would not appear, and the population would not be cleansed of many harmful alleles.
The author of this lecture would be interested in building a model where two competing species fight for a common resource. Are there conditions under which purging of deleterious mutations in a sexually reproducing species would offset the adverse “double‑sex‑value” effect?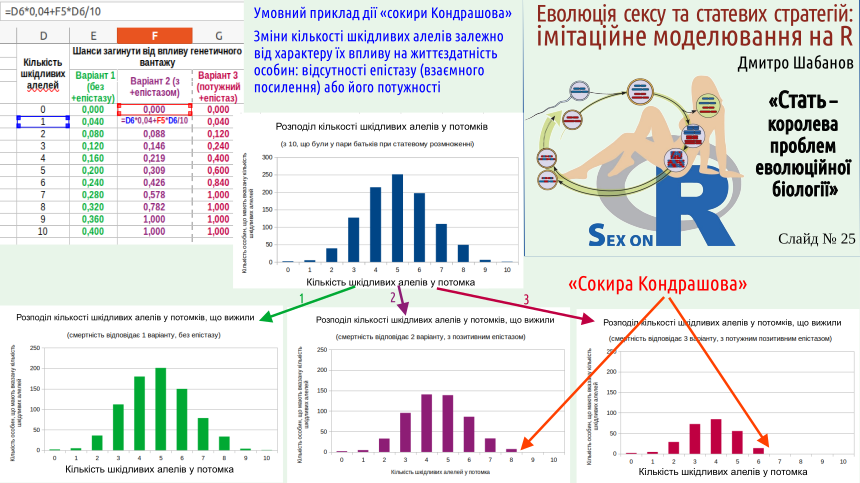
We began this lecture with Bell’s quote about sex as the queen of problems. Two of Bell’s hypotheses are to be discussed. The first is the “Brey’s vicar” hypothesis. Figuratively it can be described as: in a world where unexpected changes are possible, it is useful to produce a variety of offspring; some of them may be adapted to something still unknown.
In most cases individuals that produce offspring matching current conditions will increase their contribution to the next generation. Betting on low‑probability changes is a decision that requires foresight. As you recall, we discuss matters where there is no basis to expect predictive ability.
Could group selection support the diversity of bets envisaged by the “Brey’s vicar” hypothesis? In the author's view – perhaps (and without group selection this mechanism would not work). It would be worthwhile to test this using simulation modeling…
Williams’s lottery hypothesis develops a similar approach, but with a crucial difference. “Brey’s vicar” assumes betting on something that may happen in the future; if we talk about low‑probability changes, they must be waited for (and waiting will not work, because predictive individuals are those who increase the share of their offspring “here” and “now”). One drawback of the lottery hypothesis is that it considers adaptation to what already exists “now”, but not “here”.
Criticism of Bell’s hypothesis by Williams is partially indisputable. The models listed on the next slide are convincing. In the first two cases sexual and clonal reproduction alternate; in the third model only sexual reproduction is observed.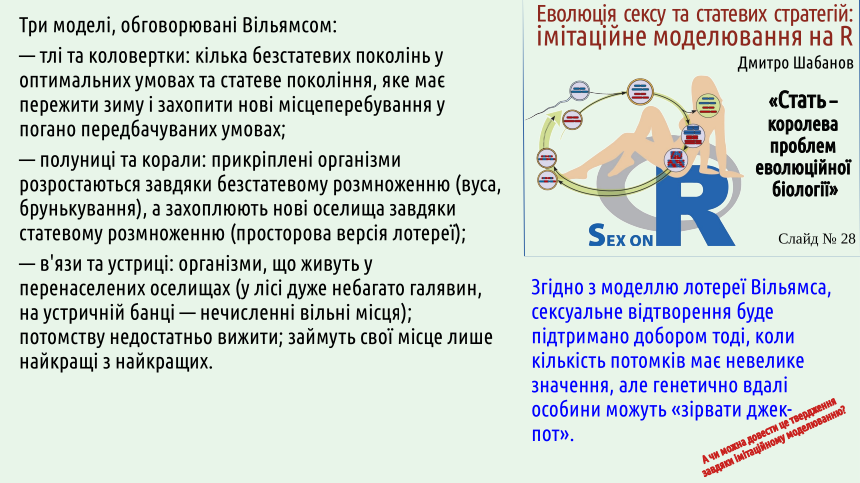
Bell’s counter‑arguments are also compelling. Indeed, suppose that the Darevskian lizards (clonal lizards we mentioned in the previous theoretical lecture) live under quite harsh conditions. Bell named this hypothesis with a nod to the famous quote from a more important book by Darwin.
Again (as with “Kondrashov’s axe”) the preservation of sexual reproduction is linked to interspecific competition. But...The lecture author finds it hard to imagine how a sexual population could outcompete a population that does not pay the “double cost of sex.”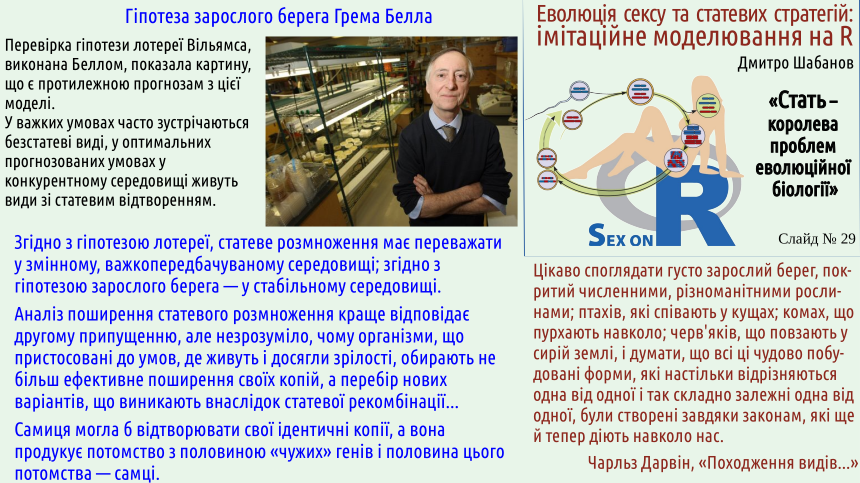
Mening’s hypothesis concerns a mechanism that unconditionally contributes to the stability of sexual reproduction. However… Neither the emergence of sexual reproduction nor
Interesting, what about possible sexual selection in a population of hermaphrodites? Imagine: each individual carries certain markers of its potential fitness and exhibits behavior that leads it to mate with individuals that have at least the same (preferably better) status. Yet, if a situation arises where it is difficult to find a partner, one must choose: mate with the one that is available, or wait for a prince on a white horse. In dioecious organisms waiting for a prince makes sense for attractive females; if they accept a neighbor’s courtship, when the prince arrives the attractive female may already be pregnant and uninteresting to the prince. And the prince may not be looking exclusively for princesses; an intriguing story with a dreamy girl from the periphery will not erase his prospects with a princess. Despite certain simplifications in the presented exposition, relationships between the sexes in dioecious organisms are based on their fundamental asymmetry (the physiological consequences of successful mating for female and male are completely different). What would happen if this asymmetry were removed in a hermaphroditic population? The problem of hermaphrodites should be examined in more detail in the next theoretical lecture.
Experimental confirmation makes Mening’s hypothesis much more weighty. The “health‑promoting” effect of sexual selection can be considered proven. The essence of the hypothesis is an attempt to explain the stability of dioecy by sexual selection. This is harder. Some influence is certainly present, but is it sufficient?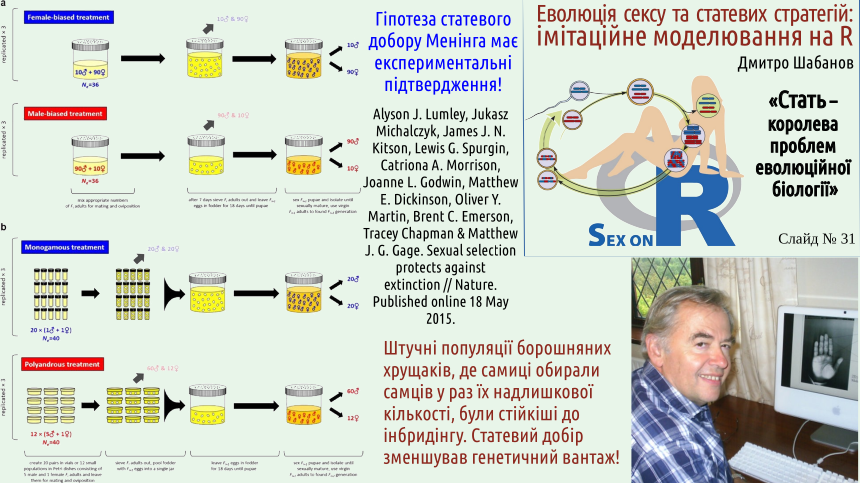
In the post‑Soviet space the views of Vigen Geodakyan are popular, among other reasons because of this brilliant scientist’s ability to promote his own ideas. Geodakyan’s scientific legacy is heterogeneous; in the author’s view, it contains undeniable biological errors. Even the quote that begins the next slide can raise many questions. Nevertheless, many of the ideas propagated by Vigen Artavazdovich are undeniably persuasive. However, these ideas do not refute the theory of sexual selection, but rather complement it.
This slide shows another model that demonstrates how substantial the difference in fecundity between two competing forms can be. In this model one can vary the relative mortality of females and males and then see how this affects inter‑specific competition. The model can be downloaded either in LibreOffice Calc format or in Microsoft Excel format.
Can this mechanism explain the Maynard‑Smith paradox? No, because a population composed of clonal females would reproduce more efficiently than any population that includes males! Can this hypothesis explain the emergence of dioecy? No. For this effect to manifest, dioecy and ecological sex differentiation must already exist!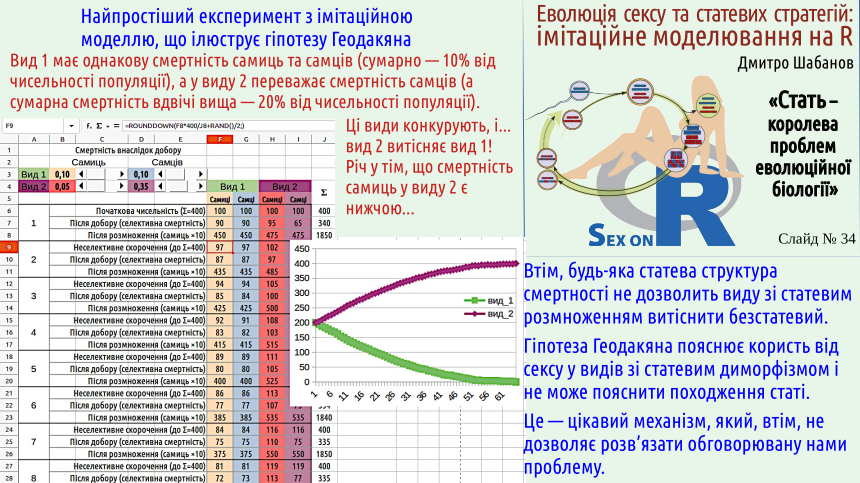
Another hypothesis that has received experimental support is the Red Queen hypothesis (sometimes translated as the Black Queen) of Li Van‑Walen. Here we see a reference to an episode from Lewis Carroll’s immortal fairy tale.
What mechanism can demand rapid evolutionary change? Defense against parasites, especially those that exploit specific biochemical “lock‑picks” to breach their hosts’ defense systems!
What happened in the experiment described on the next slide? First, pathogenic bacteria were selected. Bacteria can evolve quickly. Host defense systems act as a filter that selects for bacteria that overcome this defense and can proliferate effectively under such conditions. If the worms are genetically identical, bacteria that have been selected in one host are already optimally tuned to all others. If the hosts are genetically diverse (which is precisely what sexual reproduction provides), those individuals that differ from the diseased one from which they become infected have an advantage. Bacteria are poorly adapted to them, and while a relatively moderate infection and parasite evolution proceeds, the host can “learn” to resist the threat.
Note: in cenorabditids hermaphrodites reproduce by self‑fertilization. This is not normal sexual reproduction, nor typical clonal reproduction. One individual produces both oocytes (with a random loss of half the genetic material) and sperm (also with a random loss of half the genetic material). Fertilization occurs between the oocyte and the sperm of the same individual. This leads to half of the genes for which the parental individual was heterozygous (had different alleles) becoming homozygous in the offspring (having identical alleles). New alleles (which in true sexual reproduction would be supplied by a partner) cannot be obtained in this way; such hermaphroditic reproduction leads to a depletion of genetic diversity.
The “Red Queen” undoubtedly contributes to the maintenance of sexual reproduction. In cases where population survival critically depends on parasitic attacks, the influence of this mechanism can be sufficiently significant. But is it enough (even when combined with other mechanisms we have listed) to generate and sustain sexual reproduction? Incidentally, hermaphrodites that practice cross‑fertilization (i.e., normal sexual reproduction without dioecy) fully exploit all the advantages of the “Red Queen”…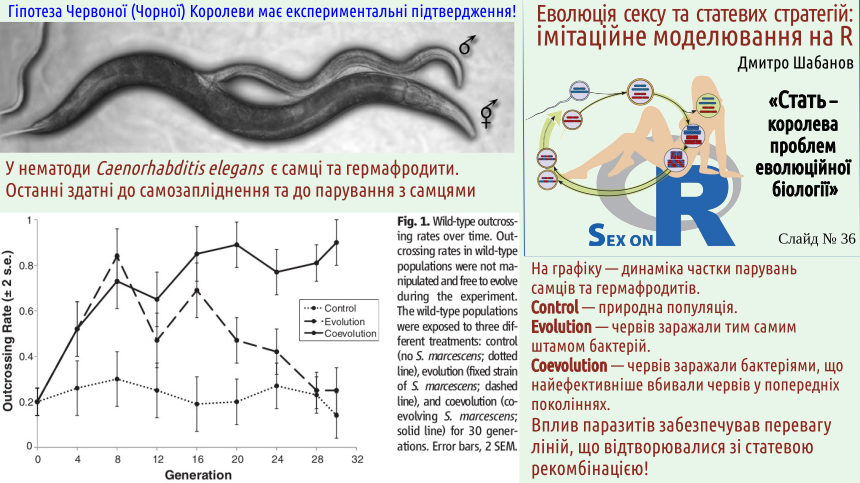
6. There are also known mechanisms that define the disadvantages of sexual reproduction compared with clonal reproduction. Among them is “Jenkins’s nightmare.”
After we discussed the advantages of sexual reproduction, we can also mention its disadvantages. One such disadvantage, the “double cost of sex,” has already been discussed. By the way, cross‑fertilized hermaphrodites do not “pay” this “cost.” However, at least one disadvantage of sex (as reproduction with recombination) also applies to hermaphrodites. Now we must examine a story that is repeatedly cited in a distorted form.
“Jenkins’s nightmare” is an argument against Darwin’s views that seemed the most compelling even to Darwin himself.
The author of this lecture was once surprised that books discussing “Jenkins’s nightmare” never quote Jenkins himself. Usually it is described as follows: “Suppose a red poppy appears in a field of white poppies; it will attract bees, but its seeds will produce pink poppies, then pale‑pink, and so on. But now we know that flower colour can depend on a single gene and be inherited as a unit! It is a pity that Darwin did not pay close attention to Mendel’s work, who discovered discrete inheritance.”
With great difficulty the lecture author managed to order a copy of Jenkins’s work from a library in the USA (now everything has become much easier – the work is available online).
Racist views at that time were widespread, almost universally accepted. Discussing this now is unnecessary. Something else is more interesting. The traits that Jenkins wrote about are clearly not those that could depend on a single gene. Under sexual reproduction these advantages would “blur” and be lost!
By the way, a shift to clonal reproduction in successful hybrids can be a way out of “Jenkins’s nightmare.” Here is an example.
A large portion of fruit‑crop varieties we grow in our orchards are hybrids that reproduce clonally. Once, through successful crossing, an exceptionally successful hybrid was noticed by breeders. In seed reproduction, the outstanding hereditary traits of this hybrid are lost, blurred in full accordance with the logic of “Jenkins’s nightmare.” That is why such successful hybrids are propagated clonally: by cuttings, grafts. Seeds produced by these plants are not sown, because the offspring from them do not retain the remarkable qualities of the parent.
Presumably, the origin of such clonal species, as discussed in the previous lecture on Darwinski, is also explained by a similar mechanism. Through crossing (often interspecific) an extremely successful hybrid arises. If its mode of reproduction does not change, it simply disappears—either because of disruption of gamete formation or because of “Jenkins’s nightmare.” If such a hybrid switches to clonal reproduction, its copies spread under suitable conditions and will persist until conditions change (or until “Müller’s ratchet” does its grim work). The capacity for further evolution in such hybrids is reduced, but for a time their existence will be relatively stable…
7. The emergence of sexual reproduction (which halves the probability of any gene being transmitted to the next generation) is most likely not explainable by selection at the gene level alone. The explanation of the discussed phenomena probably requires taking group selection into account.
Even larger theoretical problems are linked to explaining the origin of sexual reproduction itself, the origin of sex.
The quote from Dawkins on the next slide demonstrates that he is an honest person. From Dawkins’ easy hand spread the notion that evolution is the result of selection of “selfish genes,” exclusively the outcome of gene‑level selection. For Dawkins’ supporters it is obvious that the origin of sex can also be explained by such selection. The author of this lecture believes that the origin of sex cannot be explained without group selection. What is Dawkins’ own point of view?
Dawkins is a proponent of the view that selection operates exclusively at the genetic level. He understands that the transition from clonal reproduction (where each gene is passed to offspring with 100 % probability) to sexual reproduction (where genes have only a 50 % chance of being transmitted) cannot be explained by gene‑level selection. How could a selective process that is supposed to increase the fitness of genes (their ability to be passed on to the next generation) lead to a halving of their fitness? Before forming gametes, genes apparently politely give way to their competitors (the reference to Gavot) and disappear from half of the resulting cells. It seems this can be explained only by a higher‑level selection. Understanding these problems, Dawkins simply admits that he has no answer to the problem of the origin of sex…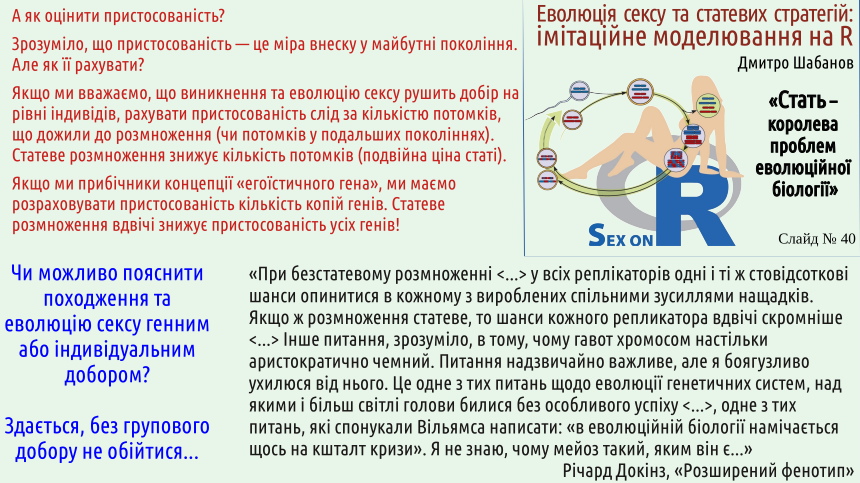
8. The hardest thing to explain is why the most common strategy among advanced organisms is not the one that might seem optimal—hermaphroditism with cross‑fertilization. Such hermaphrodites obtain almost all the benefits of sexual reproduction and reproduce with maximal efficiency because they do not incur the extra costs associated with the “double cost of sex.”
In summary, the nine listed explanations of the advantages of sexual reproduction do not fully resolve the Maynard‑Smith paradox, although they likely contain many correct ideas. However, they do not address the problem discussed on the final slide of this lecture. In general, explaining why hermaphrodites are far less widespread than they could be requires a separate lecture.