SexOnR–07_Evolution. Evolution and sex (Problem description I)
To consider the evolution of sex, one must first understand how evolution actually works. The next step — understand what sex is
Evolution and Sex The first lecture addressing the evolutionary problems of sex was video-recorded in the spring of 2022 and even edited, but... It turned out that the bombing of Kharkiv, which could be heard outside the window, did not contribute to high-quality recording. The author of this lecture did not like his own heavy speech and uneven pace of speaking very much. In the last days of winter 2023, the author decided not to record a video, but to write comments to the presentation slides. Perhaps, over time, a video recording of this lecture will appear, but for now, students will have to use the lecture in text form. We would like to make this lecture as clear and correct as possible. We ask all its readers to report any shortcomings they notice (from grammatical errors to "obscure" passages) and any ideas and arguments that come to mind.
In fact, the main topic we will be considering in this course is not even sex and gender, but evolution. To understand how evolution actually happens, it takes a lot of effort. This is a topic that could be dedicated to a separate large course. If we start to prove all the statements that we need to keep in mind, we will spend all or almost all of the time allocated for our course. Therefore, we will have to formulate a certain number of key ideas and offer them to the course listeners. If you disagree with something, write in the Telegram group, we will discuss it. In any case, you will have to get rid of common myths such as "survival of the fittest" in natural selection, "humans are as Nature wanted to create them," "there is an abyss between humans and animals," etc. Explain why this is nonsense? In natural selection, those who better fit their lifestyle in certain environmental conditions are more likely to survive. Strength is by no means a universal solution to all problems. There is no personalized "Nature" that could wish for something and plan. Humans are one of the animal species; like any species, they have certain peculiarities. Of course, the most important features of our species relate to the characteristic way we develop adaptive behavior. We are actually an unusual animal species.
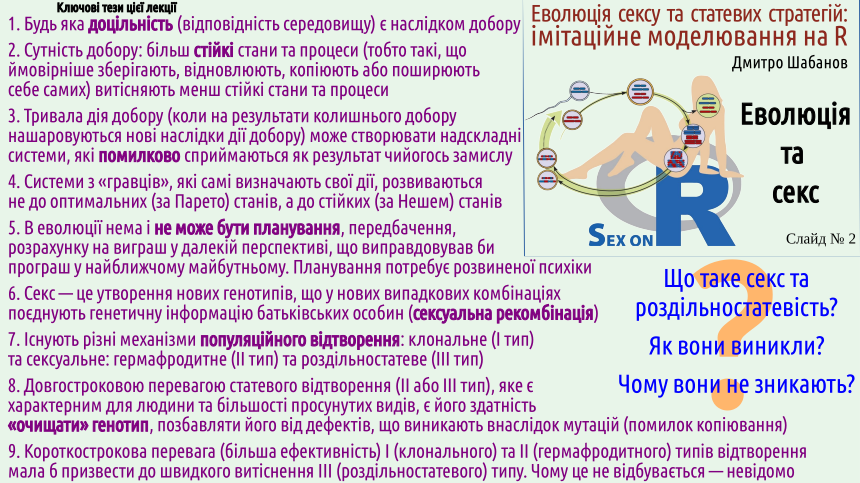
Among other things, the peculiarities of our sexual life and interactions between the sexes played a significant role in the origin and development of our characteristics. The next slide is a kind of content, a list of the main ideas discussed in this presentation... 1. Any expediency (adaptation to the environment) is a consequence of selection. It is a common misconception that natural selection is purely a biological phenomenon. One hears the argument against evolution: that since selection occurs exclusively between organisms, the very appearance of living organisms cannot be explained by selection. Of course, this is incorrect. Selection is a very broad phenomenon; its action can be observed among physical, biological, and mental and cultural phenomena (some examples will be given on slide 6). In general, information theory teaches that information arises as a result of remembering the results of random choice. If this memory is non-selective (i.e., devoid of selection), noise arises – a chaotic signal that has no value. Selective retention of information (i.e., the presence of certain selection, testing what has arisen for its suitability to the environment) is the cause of meaningful information. All complex, purposeful systems, even those that arise in the human psyche, arise as a result of selection. For example, what happens in the head of a person trying to solve a technical problem? Suppose you need to solve a puzzle. A less experienced person will try to do it by turning the puzzle in their hands. Turn it this way, that way, try turning it some other way, pull, push... After a series of unsuccessful attempts, they finally manage to find the movement needed to achieve the goal. What is this? An example of selection. The person randomly chose different movements, tested them for suitability for the goal, and retained (implemented) the one that suited the goal. A more experienced person may not make unnecessary movements. They look at the puzzle (an object of the external environment), create a model of it in their psyche, and then operate with this model.
What will happen if I do this? And this?

And what will this movement lead to? Oh, it seems the solution is found! This is also selection, but selection of random choices tested on a model. In fact, in both cases, not all possible movements are tested (whether in the process of manipulating the object or on a model), but only selected ones – those for which it can be expected that they might lead to the desired result. A certain selection is also at this level, but in any case, solving a technical problem is a consequence of selection. Here are the statements of three outstanding thinkers, each of whom evokes sincere admiration from the author – Boltzmann, Teilhard, Dennett. These are very different people: a classic of thermodynamics, a Jesuit paleontologist, a philosopher. Why do they all consider the idea of selection and evolution so important? Boltzmann's tombstone did not state who he was; above his sculptural image, a formula was given that relates entropy (a measure of disorder, S) to thermodynamic probability, W. In a more modern notation, it would look like this: S = k × ln(W). k is the Boltzmann constant, 2.07 × 10−21 J. W can be considered the number of microstates of a system corresponding to the macrostate being considered. In general, Boltzmann managed to bridge the gap between the microscopic and macroscopic states of the systems studied.

Boltzmann was a genius who was far ahead of his time and not understood by his contemporaries; the crisis caused by this led to the scientist's suicide. One of Boltzmann's most important results was that he managed to determine which changes can occur spontaneously (among other things, this direction of spontaneous changes determines the direction in which time flows). Isolated systems change in the direction of increasing their entropy, i.e., they move from less probable states to more probable ones. You understood that the thermodynamic probability of a system's state is determined by the number of microscopic ways in which that state can be achieved? 2. The essence of selection: more stable states and processes (i.e., those that are more likely to preserve, restore, copy, or propagate themselves) displace less stable states and processes. Another common (and incorrect!) argument against selection is that, according to Boltzmann's interpretation of the second law of thermodynamics, isolated systems must evolve towards increasing disorder, while living organisms are remarkably ordered. It seems that evolution contradicts the second law... No, it conforms to it!
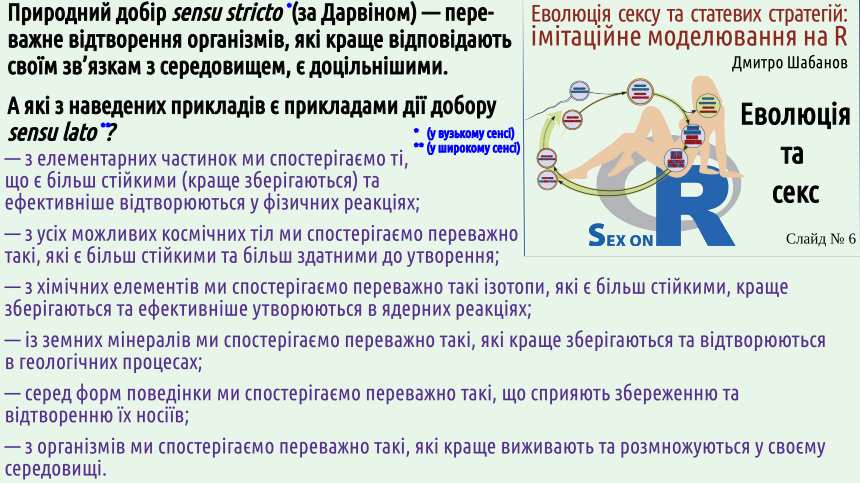
The second law requires the preservation of more thermodynamically probable states. Otherwise, one can say that more stable states are preserved. In the broadest sense, natural selection can be viewed as a process that predominantly preserves more stable states and predominantly destroys less stable states of various systems. The slide shows a number of examples of the preferential preservation of more stable systems and processes. If we consider the life cycles of organisms, which are realized in certain environmental conditions, it turns out that those cycles are stable that are more likely to be reproduced in given conditions (i.e., produce more adapted organisms), while those organisms that are reproduced with less probability are less adapted. Stable organisms are those that more effectively preserve, restore, copy, or propagate themselves! The environment in which the life cycles of organisms occur is not an isolated system: it is an open system through which energy flows.
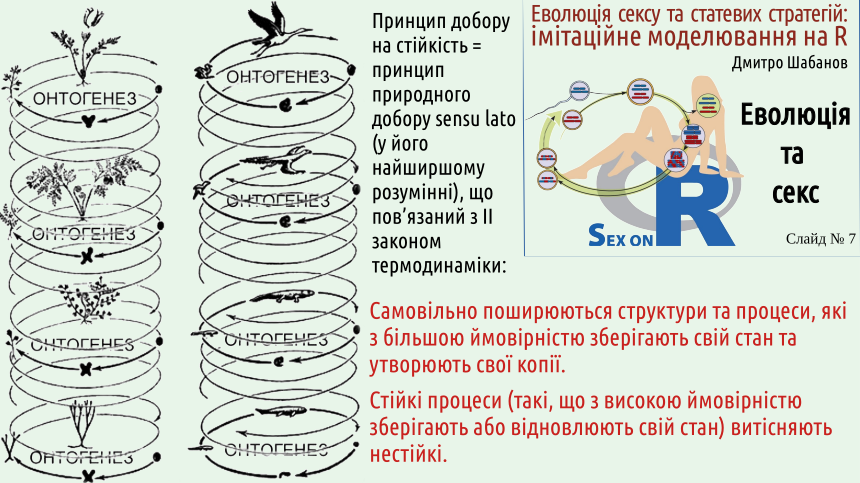
Selection, which is a consequence of the second law of thermodynamics, maintains more stable life cycles that correspond to more adapted organisms in such conditions. Ilya Prigogine was one of the outstanding researchers of self-organization in open systems through which energy flows. He showed that in so-called dissipative systems (highly non-equilibrium systems that dissipate the energy flowing through them), complexly organized structures can arise spontaneously. This is a consequence of the increase in overall disorder required by the second law of thermodynamics.

Processes in which Darwinian natural selection operates are an example of such spontaneous self-organization. 3. The prolonged action of selection (when new consequences of selection are superimposed on the results of previous selection) can create super-complex systems that are mistakenly perceived as the result of someone's design. Let's return to the first thesis. Can it be that expediency arises not as a result of selection, but solely by chance? Let's consider another common argument against evolutionary biology. They cite supposedly calculations of the probability of a random appearance of an amino acid sequence of a particular protein, or, for example, the probability that the molecules that make up a living cell will randomly combine into the structure we observe. Of course, these are fabrications. A result that requires a single instance of randomness can arise by chance. Let's consider a simple example. Do you know this game: one hides an object, and the other searches for it by suggesting various movements and receiving answers "hot" or "cold." By the way, sometimes the searcher reads the partner's unconscious reactions and, by comparing reactions to different movements, determines where the hidden "treasure" is. If the players have enough patience, very complex instructions can be conveyed in this way. It may be that first, through trial and error, one must understand which room to enter; then (in the same way) – which object to choose in that room, then – what to do with that object... The results of trial and error (=selection), which establish subsequent actions, are superimposed on the results of trial and error that led the player to the right place. Can it be done without selection? You thought of a specific unambiguous number, I tried to guess it, said, for example, "3" and... guessed it! Was there selection here? In any case, when we established that I guessed it (i.e., tested my guess, tested the trial for suitability to the environment, i.e., the number you thought of), we already used the selection procedure. That is why a living cell did not arise from molecules randomly "jumping" together in the right way. It is the result of prolonged, "multi-layered" selection. How this could have happened is a problem that can be discussed, but it would take us too far from the topic of our course. Interestingly, our intuition is not adapted to recognize the results of "multi-layered" selection. When we look at complex systems that are the result of prolonged selection, we perceive them as something pre-planned. The fact is that our intellect is evolutionarily formed not as a means to achieve truth, but as a means of adaptation to existing conditions.

It is better to see a design where there cannot be one than not to see it when it is there (for example, when it is necessary to understand the intentions of a cunning opponent). 4. Systems with "players" who determine their own actions do not evolve towards optimal (Pareto) states, but towards stable (Nash) states.

Vilfredo Pareto described what state of a system of several players is optimal for everyone. This is a situation of maximum mutual benefit.

A world built according to Pareto would be optimal. In cases where we deal with systems where planning is possible, Pareto states can be achieved. Unfortunately, if the players are independent, they act to maximize their own benefit, regardless of how their actions affect the interests of other players.
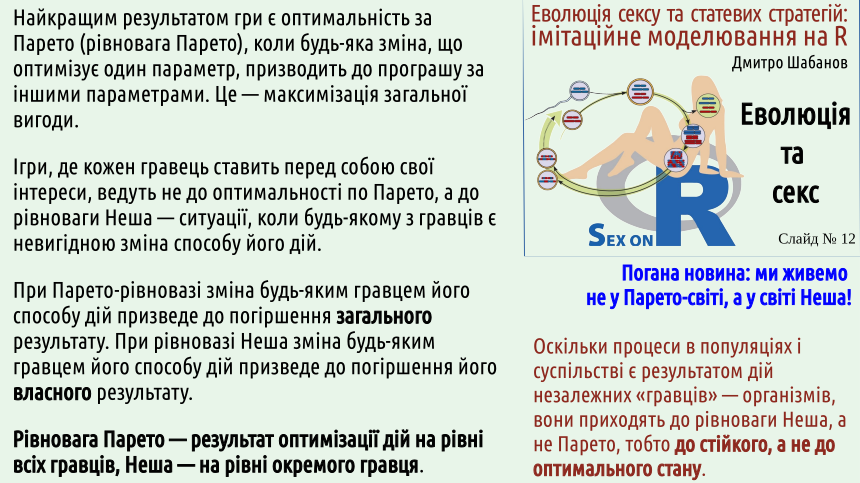
This was first shown by John Nash. Unlike Vilfredo Pareto, who was likely a very good person, Nash suffered from schizophrenia and was a rather difficult person to communicate with. A world living by Pareto would be much better than a world evolving by Nash. ...Unfortunately, we live in Nash's world...

Why do players choose Nash equilibrium and not Pareto optimality? Because they prioritize their own interests. This can be illustrated by the famous "prisoner's dilemma." Its formulation is given on this slide. The "prisoner's dilemma" was formulated in 1950, but investigators began to use it in their work much earlier. Maximizing mutual benefit, Pareto optimality corresponds to the case where both prisoners remain loyal to each other. Nash equilibrium corresponds to the case where both prisoners "betray" each other.
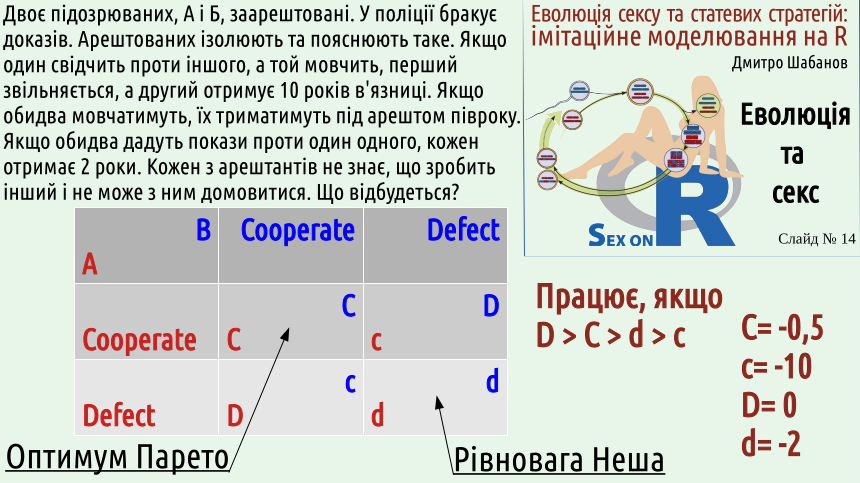
WHY? Put yourself in the place of one of the prisoners. Suppose your accomplice remains silent. If you testify, you will be released, and if you remain silent, you will serve six months. Thus, if your partner remains silent, it is more advantageous for you to testify. Another situation: your accomplice testified against you. If you testify, you will serve two years, and if you remain silent – all ten! Thus, if your partner testifies, it is more advantageous for you to testify. In both cases, it is more advantageous for you to testify! Well, tell the investigator everything, and be sure that your accomplice will do the same! 5.
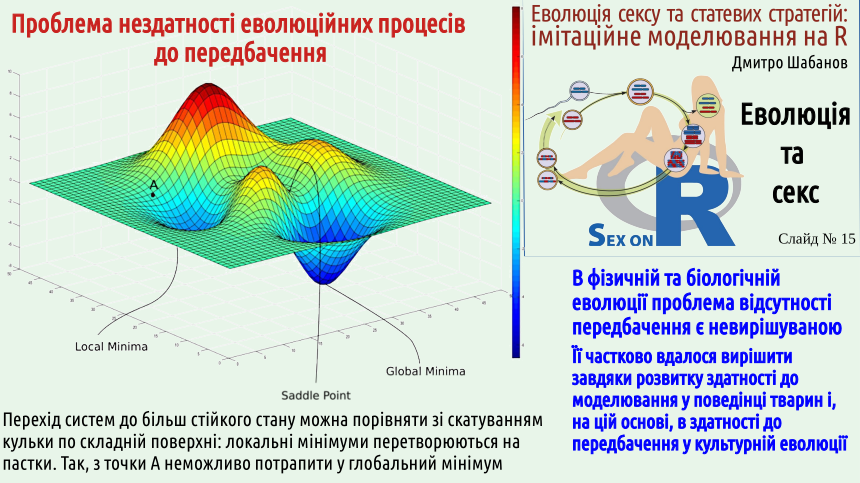
In evolution, there is no and cannot be planning, foresight, or calculation for a long-term gain that would justify a loss in the short term. Planning requires a developed psyche. When the fate of a system of several players is decided by one person capable of planning and predicting the consequences of their actions, there is hope that the result of the development of such a system will be Pareto optimality. But even in cases where we consider the actions of independent players, even if they can plan and predict the future, the system will fall into Nash's trap. Sad... Note: this applies even to the evolution of systems consisting of predictable players! And what will happen to systems where there are no planning and predicting persons, i.e., any physical and biological systems, except for those where the behavior of highly organized animals, such as humans, plays a decisive role? There is no basis to expect consideration of the possible consequences of the processes that will occur. Evolution does not proceed towards Pareto optimality; it proceeds towards Nash equilibrium! So, is it generally impossible to achieve Pareto optimality? It is possible when we deal with a system centrally controlled by an intellect capable of foresight. The evolution of the psyche of highly developed animals has gone in the direction of improving models of the environment. Using this model, the animal can build a forecast (remember the example with the crow!) and act accordingly. Humans have surpassed all other species in this ability and have even created artificial intelligence.
In the human psyche, as we have already discussed, a certain form of selection operates; selection is quite often used in the work of artificial intelligence. For example, to find global minima in cases similar to the one shown on the previous slide, algorithms start optimization from random positions on the studied plane, and then determine which attempt led to the best result. From the foregoing, there is an interesting consequence. When studying the consequences of evolution, it is not advisable to ask the "childish" question "why is this done?". The things that Matt Ridley talked about, quoted on slide 9, are generally not the result of any planning! Understandably, they are not the result of pure chance either; without selection, chance cannot form anything complex.
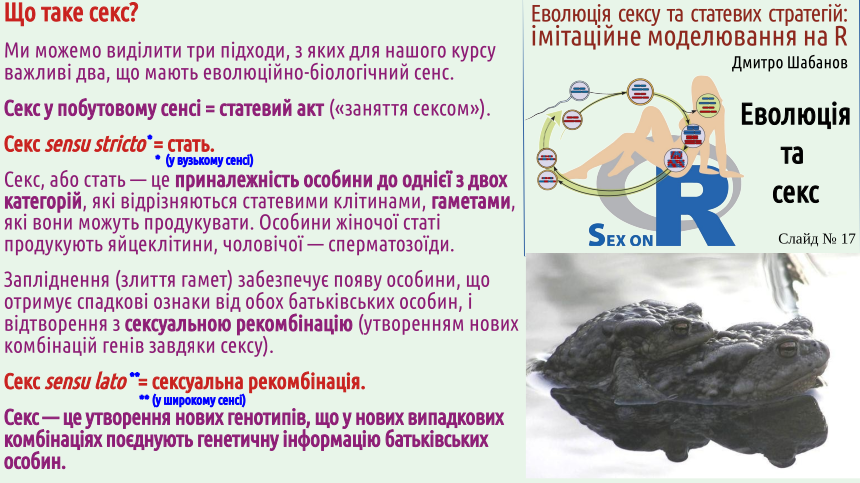
If we try to understand why certain phenomena exist, we should establish how and why stable states arise, the consequence of which is the existence of such phenomena. All of the above was an introduction (a prelude, if you will) to our main conversation. It is important that when considering sexual reproduction, you ask yourself not the question "for what purpose?", but the question "why?". Why does sex exist? 6. Sex is the formation of new genotypes that combine the genetic information of parental individuals in new random combinations (sexual recombination). We have finished our introductory talk about evolution and are finally moving on to consider sex! Let's start with definitions. We've already talked about this, but in this lecture, we should also provide these definitions.
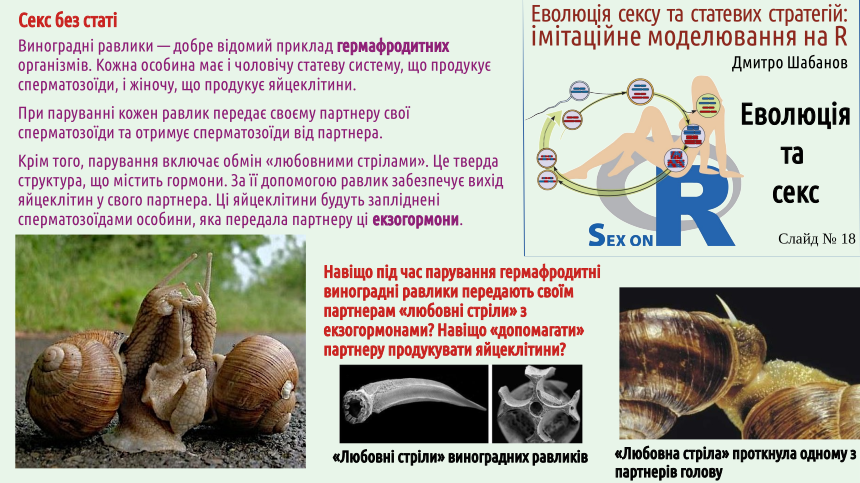
Students who came to our course to learn how to get more pleasure from sex may be disappointed: the pleasure that can be obtained from our classes is mainly intellectual. Does it seem to you that sex is inextricably linked with gender? There are many creatures around us that practice sex but do not belong to the female or male sex. These are not only the snails shown on the slide, but also earthworms. A significant portion of angiosperms (flowering plants) are monoecious, meaning they produce both pollen and seeds (i.e., they are effectively hermaphrodites). We will not delve into the discussion of whether hermaphrodites have sex (in the opinion of the lecture author – no, but some of his colleagues disagree). We can also mention other examples. One of them is as follows. The green alga Spirogyra is one of the most common freshwater algae; it often participates in water "blooms." The body of Spirogyra is an unbranched filament. Two such filaments approach each other, and conjugation occurs. The contents of a cell from one filament flow into the cell of the other filament and merge with it. Not gametes, but simply cells participate in fertilization! A spore (zygospore – a spore formed as a result of fertilization) is formed, which overwinters and undergoes meiosis the following year. The vast majority of cell divisions in the human life cycle (which can be considered a typical example of a life cycle with sexual reproduction) are mitoses, i.e., divisions that result in the formation of two genetically identical (clonal) cells. Almost all cells in our body are clones (genetic copies) of the zygote, the fertilized egg cell. These cells have a set of chromosomes inherited from the mother and a set of chromosomes inherited from the father. These cells form the soma, the body itself. However, the zygote itself is not part of the soma; it belongs to the set of potentially immortal cells, the germ line or germ. Most descendants of the zygote form the soma, but some of them become cells that form gametes (sex cells). In the body of a young woman, there are several hundred thousand cells that can develop into a female gamete, an egg cell. These precursor cells were formed when the future woman was still a fetus and was inside her mother's uterus. As a woman ages, the number of egg precursor cells irreversibly decreases. Throughout a woman's life, her body may produce several hundred egg cells – quite a few, if this number is compared to the number of sperm. Men need to produce a huge number of gametes. A mature man can produce several trillion sperm in his lifetime (remember that the current human population recently exceeded 8 billion). The phenomenon of sex is precisely related to the formation of gametes. During gamete formation (gametogenesis), the amount of genetic material in the cells is halved.
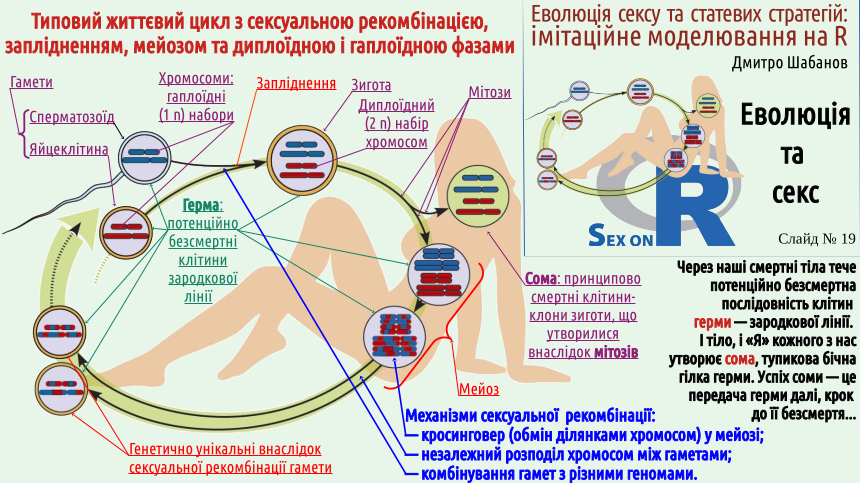
This is ensured by a special type of cell division – meiosis. During meiosis, random exchange of fragments of maternal and paternal chromosomes occurs (during a process called crossing over). The result is that all gametes are genetically unique. It will be easier for you to believe that a woman does not have two identical egg cells, because as a result of crossing over and independent distribution of hybrid chromosomes formed by crossing over between daughter cells, each cell receives its own version of genetic information. Look at the diagram on the slide: it conditionally shows this process! And what is the probability that out of the trillions of sperm produced by a man, two will be genetically identical? It is less than one divided by the number of atoms in the universe! This diversity is the result of sexual recombination (the formation of new combinations of genetic information). If you look at the slide, you will see that different features of the life cycle of organisms with sexual recombination are related. Fertilization doubles the amount of genetic information, and therefore it must be halved during meiosis. The mechanism of this reduction is such that it causes the formation of genetically unique gametes; the probabilistic nature of gamete combination is an additional source of genetic diversity of organisms formed as a result of fertilization.
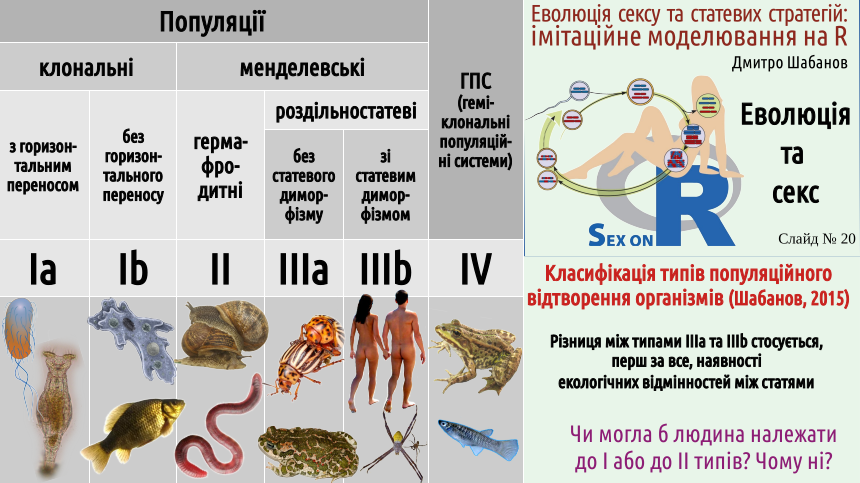
7. There are different mechanisms of population reproduction: clonal (Type I) and sexual: hermaphroditic (Type II) and gonochoric (Type III). It's time to propose a classification of organism reproduction methods that we will use. We will distinguish between reproduction (what happens to organisms) and replication (a phenomenon characteristic of the population level of biological systems). Population replication occurs through organism reproduction... Why are we considering population replication, not organism reproduction? Because populations evolve, as they are potentially immortal, while organisms are fundamentally mortal. So, populations can be clonal (replication in which occurs without sexual recombination) or Mendelian (named after Gregor Mendel, who made a huge contribution to the study of sexual reproduction). The diagram, in addition to populations, shows hemiclonal population systems – biosystems that can arise as a result of a special form of interspecific hybridization. These systems are the object of research by the author of this lecture, and he would not want to remove them from the diagram. However, in this course, we will not consider hemiclonal population systems. We will be interested in populations of Type I, II, and especially III. As you can see, the classification of reproduction types also shows subtypes of the main types. The boundary between these subtypes is more blurred than the boundary between the types. Nevertheless, it should be taken into account that clonally reproducing organisms can, to a greater or lesser extent, practice what can be a certain analogue of sex – the transfer of genetic information from organism to organism. This process occurs much less controlled than in true sex and is called horizontal gene transfer (vertical transfer is considered the transfer from ancestors to descendants). Most likely, the ability for horizontal transfer is typical for Type I populations; cases where this ability is lost are most likely dead ends. Among gonochoric populations, we can distinguish those in which females and males lead approximately the same lifestyle, and those where the ecological roles of representatives of the two sexes are sufficiently different. The difference in ecological roles of the sexes causes sexual dimorphism – a difference between the sexes not directly related to reproduction. Sexual dimorphism is also characteristic of humans, but, for example, in many spiders, it is more pronounced. After this introduction, we can compare the named types of reproduction. Remember Robinson Crusoe, the hero of Daniel Defoe's famous novel, written at the beginning of the 18th century? The classic English novel began with this book...
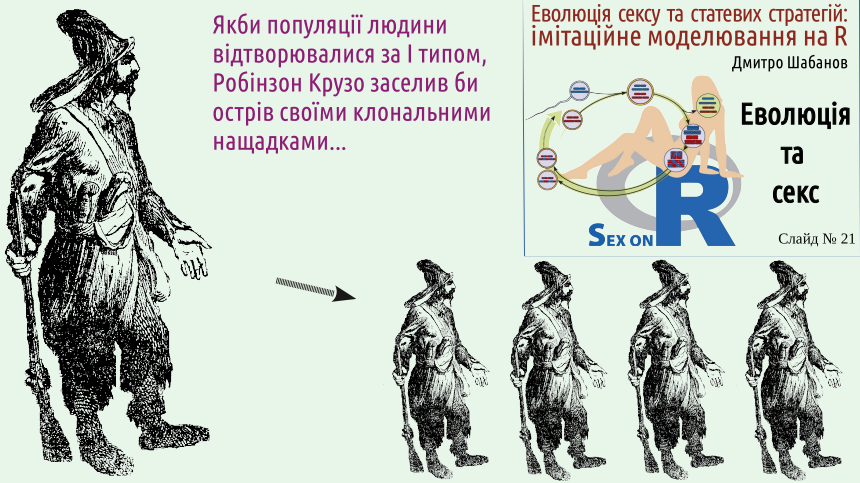
So, Robinson was a sailor who ended up on a desert island and lived there for 28 years. Thanks to his hard work, he built a micro-world perfectly suitable for life. By the way, the stories of people who found themselves on desert islands in the real world were not as successful as the story of the fictional Robinson; humans are biological beings adapted to function as part of populations. But even for the literary hero, his loneliness became the main problem. How good it would be for Robinson Crusoe if human populations reproduced according to Type I! Even a lonely individual could create a certain number of their genetic descendants. For an isolated individual, this method of reproduction would be an advantage.
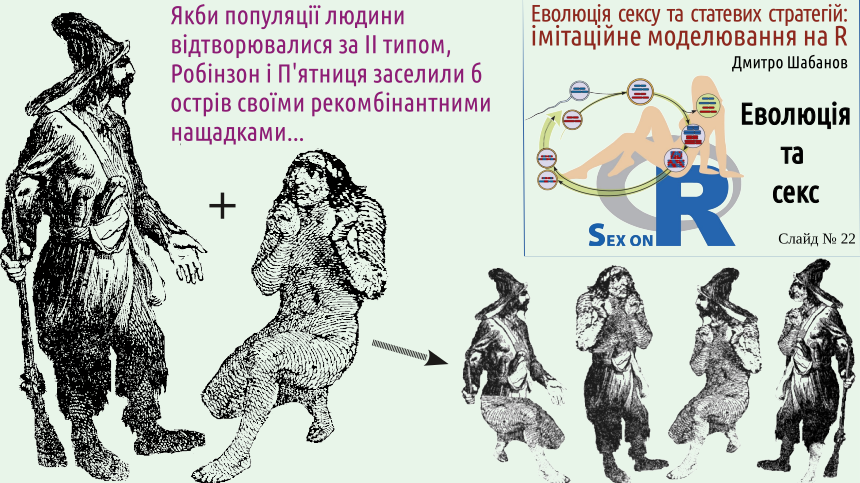
Of course, clonal descendants would appear without fur clothes, a rifle, or even a beard and mustache, but copying the image of Robinson on the slide helps to show that his descendants, in case of clonal reproduction, would be genetically identical to him. After many years of living alone, Robinson observes cannibals arriving on the island, who are about to eat prisoners they brought with them. Robinson saves a prisoner whom he names Friday.
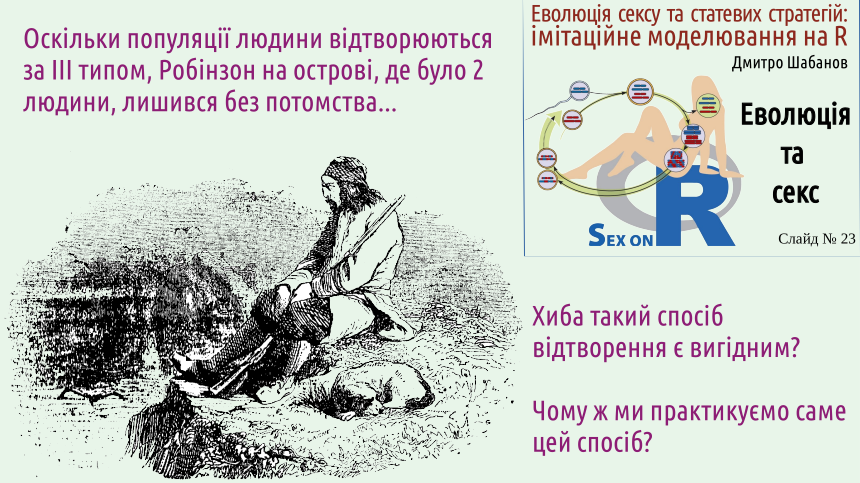
Over time, Friday becomes Robinson's loyal companion. Imagine if humans reproduced according to Type II. In hermaphrodites, any two individuals at the appropriate stage of their life cycle can become sexual partners and leave offspring. Unlike Type I reproduction, in this case, the offspring will be genetically diverse. This diversity is shown on the slide by combining the traits of Robinson and Friday. This is very inconvenient: a potential partner for a human (among individuals of the appropriate age) can only be half of the individuals! With a probability close to 1/2, two randomly chosen individuals from a Type III population are unable to reproduce together! Isn't it strange that our method of reproduction is so "inconvenient"? 8. The long-term advantage of sexual reproduction (Type II or III), which is characteristic of humans and most advanced species, is its ability to "cleanse" the genotype, to rid it of defects arising from mutations (copying errors). Let's separate a few questions. The first is why humans arose among creatures with sexual reproduction. The answer to this question is known, and we will discuss it now. The second is why humans are specifically gonochoric, reproducing by Type III. The answer to this question is more complex, but the author has a hypothesis that explains this phenomenon. We will discuss this problem in the next lecture dedicated to the issues of our course. An interesting tool for determining how logical this hypothesis is is simulation modeling. The third is why humans retain gonochoric reproduction. There is no generally accepted answer to this question; in the author's opinion, to answer this question, one must abandon the viewpoint common in modern evolutionary biology. Simulation modeling will also be very useful for researching this problem. Understanding that a comprehensive answer to the first question does not resolve the second and third, let's start from the beginning... The more complex the system, the more possibilities it has to break down. To create such a complex creature as a human, it is necessary that its development is controlled by a very complex and perfect genome (a complex of hereditary information). Let's think in the most general terms, what affects genome evolution? When considering any genome, we can see that it becomes more complex as a result of selection and degrades as a result of the mutational process (random changes). Humans are perfect because their hereditary information contains the concentrated experience of survival and reproduction of all their ancestors who have existed on our planet for almost 4 billion years! Think about it: we know that your parents (mother and father) successfully reached sexual maturity and successfully left at least one offspring. Not all people, not all organisms in general, are so lucky! Each of you can understand that your parents are winners in natural selection, whatever fate awaited them after they gave you life. But if there are winners, there must be losers. These are representatives of your parents' generation who did not reach sexual maturity or did not leave offspring. Not only your parents are winners in natural selection, but all four of your grandparents. In their generation, there were also losers; by the way, in addition to those who did not reach the required age or did not reproduce, those who left offspring that eventually died or did not reproduce also lost in natural selection. The one who loses in natural selection not only becomes a loser himself; he takes away the victory associated with his existence from all his ancestors! The winners in selection are all eight of your great-grandparents, all sixteen of your great-great-grandparents, all 32..., 64..., 128..., 256..., 512..., 1024... We now live in a world where most newborns reach sexual maturity, but a few generations ago the situation was completely different. We are descendants of a huge line of winners! And now we can explain why we mentioned this. The genomes of your ancestors differed from the genomes of their contemporaries. Some of these differences increased the chances of winning in natural selection. Of course, sometimes individuals with brilliant potential lost, and carriers of genetic defects won. However, generation after generation, selection preserved genetic potentials in the genomes that contributed to victory (i.e., those that increased the expediency, the adaptation of organisms to their lifestyle in their environment), and sex "shuffled" these potentials and caused testing of new combinations for perfection. We have described the process that causes the improvement of our genome. Unfortunately, the reverse process, leading to its degradation, operates continuously. The genetic information copying system operates with fantastic efficiency, but it also inevitably makes mistakes. The consequences of these mistakes should be eliminated by negative selection: their carriers should be less likely to win in selection than individuals without such defects.
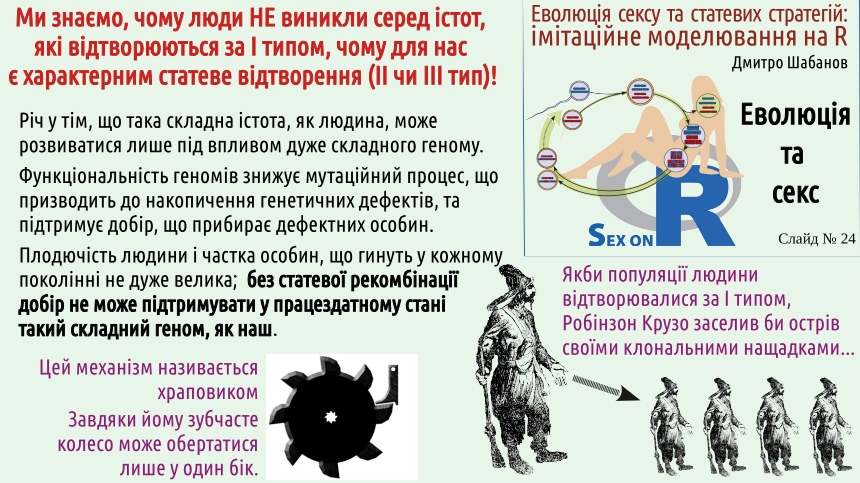
The larger the genome, the more errors it contains, the more effective the selection that purifies the genome from them must be. How to measure the effectiveness of such purifying selection? By the number of individuals excluded from reproduction, i.e., those who lose in selection. The larger the genome, the more individuals in each generation must lose in natural selection to keep it in order! Do you understand, by what mechanism can handcuffs only tighten on an arrested person's hands, and cannot release his hands on their own? This mechanism is called a ratchet.
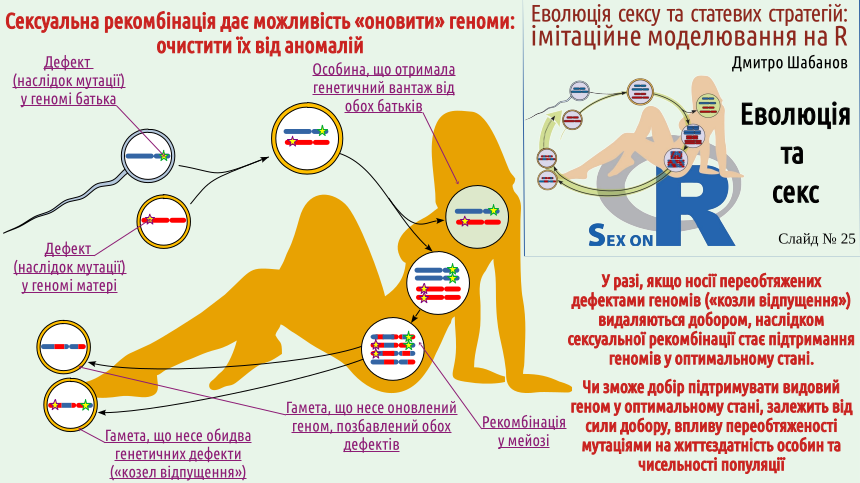
Now you will understand why we mentioned it... Let's return to the diagram of sexual reproduction (the same one that is on the logo of our course). Consider the case where both the mother's chromosome and the father's chromosome carry a defect – a consequence of mutation. Each somatic cell will carry both defects, but among the gametes, there will be those that carry one defect, and those that carry both or neither! If the discussed defects reduce the chances of their carriers winning in natural selection, carriers of both defects (they can be compared to the biblical "scapegoats") are more likely to disappear from the population. On the other hand, individuals free of both defects, purified by sex, will also appear in the population! And what will happen in populations that reproduce without sex? Let's consider this using the example of clonal lizards common in the Caucasus (as well as in an artificial population that exists on the Southern Bug in Ukraine). Several species of such lizards are now known, which were later classified into the genus Darevskia, named after Ilya Sergeevich Darevsky (1924–2009), who discovered the mechanism of their reproduction. Darevsky is considered a Russian scientist, although he was born in Kyiv and studied at Kyiv University before moving to Leningrad. One of Darevsky's main achievements was the discovery (in the mid-20th century) of parthenogenesis in Caucasian rock lizards. This refers to a mechanism of reproduction by females that does not require the participation of males. This is an example of Type I population reproduction, and a secondary one. The ancestors of Darevskia were certainly gonochoric animals, but at some stage of their evolution, they switched to parthenogenesis. All their cells, both those forming the soma and those forming the germ, divide only by mitosis. No sex – no purifying effect on the genome level!
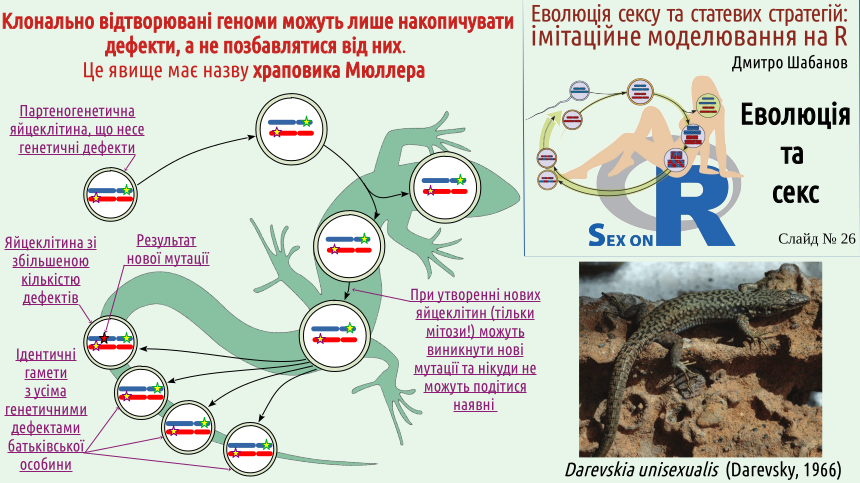
There is no genetic recombination (formation of new gene combinations), and the genetic defects that Darevskia acquired cannot go anywhere (the probability of a reverse mutation that restores a mutant region of a chromosome to its normal state is critically low and can be simply neglected). Mutations can only lead to an increase in the number of defects that parthenogenetic Darevskia eggs will carry. These parthenogenetic eggs do not require fertilization; individuals that develop from them will, at best, receive all the genetic defects carried by the maternal individual, and at worst, will receive some new defects in addition to the old ones.

Now you can understand why Hermann Muller, who described this phenomenon, compared it to a ratchet. In the absence of sexual recombination, the number of genetic defects can only change in one direction – towards an increase in their number, like a ratchet that can only rotate in one direction. The reasoning presented may seem detached from reality... Let's consider an example related to our genome. The next slide shows the karyotype (chromosome set) of a human. As in many other animal species, human sex is determined by a pair of sex chromosomes. There are two types of sex chromosomes. The X chromosome is a relatively normal chromosome – not the largest, not the smallest. This chromosome carries a sufficiently large number of genes whose function affects a number of individual characteristics. In contrast, the Y chromosome is essentially a fragment of the X chromosome; it has a much smaller number of genes, the main one being the SRY gene (Sex-determining Region Y). This gene's function is to switch development to the male type. A male will develop from a fertilized egg with the genotype XY (i.e., carrying one X chromosome and one Y chromosome) because at some stage of embryonic development, the Y chromosome will activate the function of all genes that make a person male. From an egg with the genotype XX, a female will develop. Sperm can carry either an X or a Y chromosome; egg cells only carry the X chromosomes present in the female. The sex of the child is determined by which sex chromosome the sperm carries, as the egg cell will in any case bring an X chromosome (of course, we are considering the typical case here; various anomalies associated with deviations from it are also known).
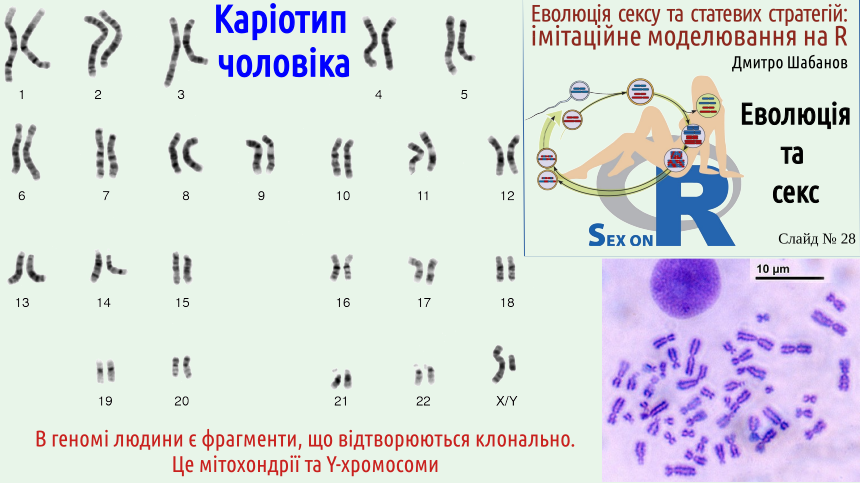
You understand that the next slide shows the karyotype of a male, not a female, because this karyotype includes both an X and a Y chromosome. But here arises an interesting question: why does the male sex chromosome look like a fragment of the female one? As shown in our course logo, for crossing over to occur between the maternal and paternal chromosomes, they must approach each other, and for this, they must "recognize" each other. Genetic sequences that correspond to each other are located next to each other, and then exchange fragments. In males and other typical mammals, X and Y chromosomes "recognize" each other, but the exchange of segments over most of their length is almost completely blocked. Let's start from the beginning: from the appearance of mammalian sex chromosomes. About 150 million years ago, the SOX3 transcription factor gene, located on the X chromosome, duplicated. One of its copies changed and turned into the SRY gene. This gene took over the initiation of male-type development. Selection supported the location of other genes necessary for male sex determination near the SRY gene – for example, genes that control sperm production and some other male characteristics. If crossing over broke this "male" complex, its effectiveness decreased. Therefore, selection supported a simple anomaly that interferes with crossing over: an inversion, a 180° rotation of a key region of the Y chromosome. The chromosomes as a whole recognize each other, but oppositely directed regions do not. In females with the XX genotype, sexual recombination maintained the functional state of the X chromosomes. In males, recombination in the inverted region of the Y chromosome turned out to be significantly suppressed, because the Y chromosome pairs with the X chromosome, where the genetic sequences are oppositely directed. If a defect appeared in the X chromosome, it was removed by sex (i.e., sexual recombination). If a non-critical defect appeared in the Y chromosome, it remained there, and the necessary functions were provided by the genes of the X chromosome. After selection stopped supporting the functional state of Y-chromosome genes, the mutational process quickly destroyed them (you remember that the functional state of a genetic program requires its constant support by selection?). And what happens to the Y chromosome if mutations damage the same SRY gene that initiates male development? Such a phenomenon is known and is called Swyer syndrome. Patients with this syndrome appear externally as women but have undeveloped gonads. Of course, they do not leave offspring, effectively removing the defective variant of the SRY gene from the population. In rare cases, crossing over between regions important for sex development of X and Y chromosomes still occurs. As a result, the SRY gene can end up on the X chromosome, but even this anomaly will not have long-term consequences. What happens if, in an individual with a female genotype, XX, one (or both) chromosomes carry the SRY gene? An individual will develop who appears externally as a man and is infertile; this is called de la Chapelle syndrome. As a result, when an X chromosome with an uncharacteristic fragment enters a genetic female, the infertility of this woman will ensure the removal of the altered chromosome from the population's gene pool. What is the fundamental difference between most genes on the Y chromosome and the gene that is the main switch for sexual development? If defects in the former are not critical, they are filtered by selection inefficiently. "Breakdowns" that lead to impaired sperm production will be removed by selection even under the Müller ratchet. But a defect in the sex development switch SRY will be removed by selection quite effectively. Unfortunately, a large number of people do not understand this last point. Look at fragments of a Russian popular science article (unfortunately, such reasoning can be found in other languages as well). Not very insightful biologists, who have registered a decrease in the number of functional genes on Y chromosomes (but do not understand how selection works), are trying to calculate when the last gene will disappear. And what will this lead to? To the disappearance of men, i.e., to the extinction of humans, or to the transformation of women into "Amazons" who will reproduce "unnaturally"!
A-a-a-a, catastrophe awaits us! Of course, the fundamental difference between the consequences of mutations in different genes on the Y chromosome is the reason why the process of their "disabling" should not proceed at a constant rate. The Y chromosome becomes simple enough that its functionality can be maintained by selection even without sexual recombination. Now you understand why the human genome as a whole requires sexual recombination for its evolution, while the much simpler Y chromosome (whose breakdowns lead to its rejection by selection) does not? But, remember, at the beginning of this, 8th part of the lecture, we noted three different questions related to the characteristic type of human population reproduction? We will leave the second question for further discussion, and move on to the third (why humans retain gonochoric reproduction) right now. 9. The short-term advantage (greater efficiency) of Type I (clonal) and Type II (hermaphroditic) reproduction should lead to the rapid displacement of Type III (gonochoric). Why this does not happen is unknown. Almost half a century ago, Maynard Smith (this is a double surname, and the name of this outstanding English biologist is John) pointed out one huge question related to gonochorism. Look at the diagram shown on the yellow background. In gonochoric organisms, only females directly leave offspring. Males are necessary for this process, but they do not give birth to offspring. The consequence is a phenomenon that Maynard Smith called the "double cost of sex" or the "cost of males."
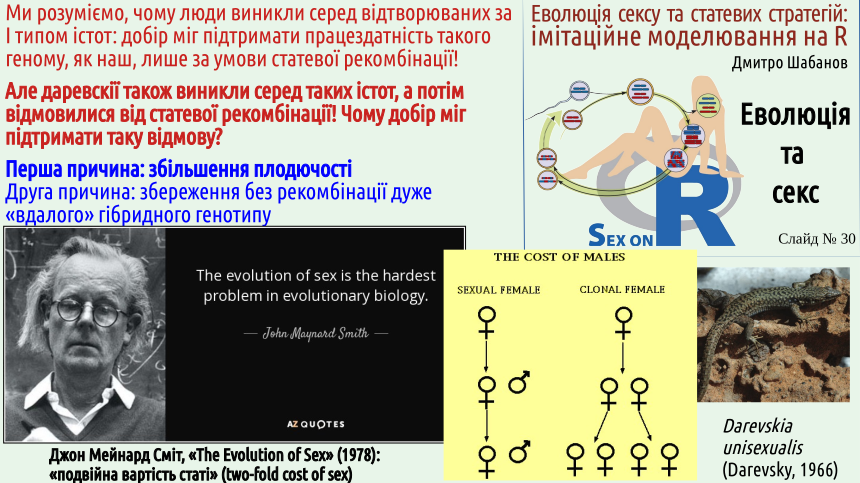
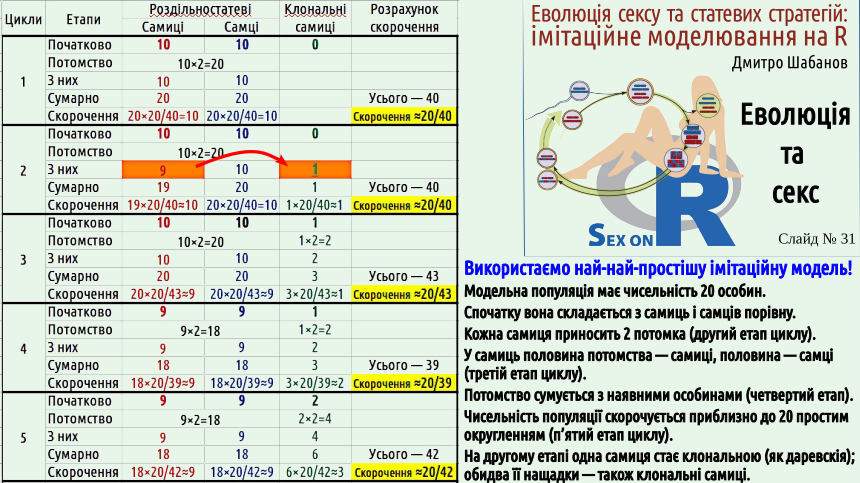
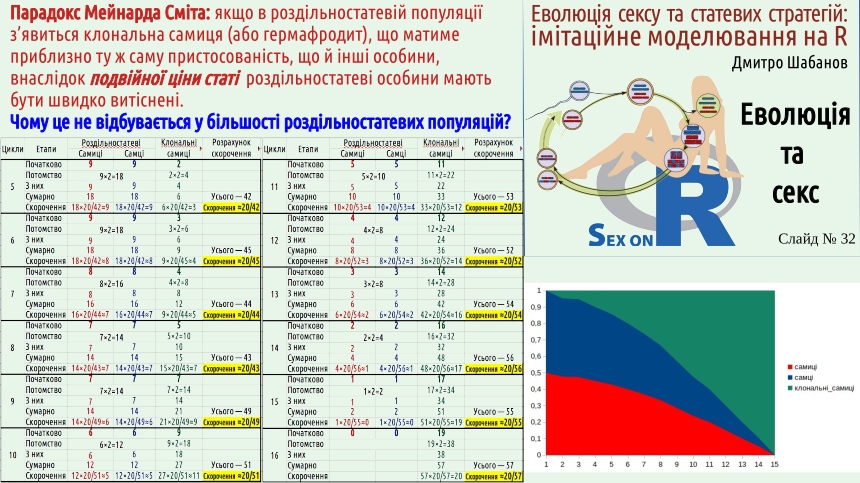
The circumstance that Maynard Smith drew attention to raises a number of questions. How can a gonochoric species withstand competition with a similar species that reproduces clonally, since the reproductive efficiency of a clonal species is twice as high? Let's consider two common answers to this question. First. A gonochoric species will use all the advantages of sexual reproduction, and a clonal species will lose due to the Müller ratchet. This argument does not work for two reasons. Firstly, hermaphroditic populations are not forced to bear the costs of the "double cost of sex." In hermaphrodites, any individual can directly leave offspring, and all the advantages of sexual recombination will work. Secondly, even a clonal species would initially have to win against its gonochoric competitor, and only then would it feel the adverse effects of the Müller ratchet. Population dynamics, interspecific competition are relatively fast processes, their consequences will manifest themselves in the short term. Genome degeneration due to the Müller ratchet will only manifest in the long term, and as you remember, there is no foresight in evolution. Second answer. It may be that the reproduction rate of both clonal and gonochoric species competing is excessive; it is as if in such a case, the reduction in reproductive efficiency of the gonochoric species by half would only benefit it. But even in the case where most offspring die, natural selection, with the same number of breeders, will involve twice as many clonal offspring. If all young individuals have equal chances of winning in natural selection, this will lead to an increase in the number of clonal organisms in the next generation. But the "Maynard Smith paradox" can be formulated even more strangely. Based on observations of many species, we know that in populations of gonochoric species, parthenogenetic females sometimes appear, such as we can observe in Darevskia lizards. The appearance of such females is a transition from Type III reproduction to Type I. It can be assumed that such a transition is possible for many gonochoric populations. According to simple calculations, a clonal female that appears within a gonochoric population should displace her gonochoric relatives in a very short time. Yes, in the long term, the offspring of such a female will suffer from the adverse effects of the Müller ratchet. However, as the example of the same Darevskia shows, clonally reproducing organisms can still exist for a sufficiently long time. To win in intra-population competition, a much shorter time is sufficient. Let's formulate the "Maynard Smith paradox" as follows: why does the appearance of parthenogenetic females in populations of gonochoric organisms not lead to a rapid abandonment of sexual reproduction? To demonstrate how such an abandonment can occur, we will use a simulation model, but not in the R language, which we will use in most of our course models, but a much simpler one created using spreadsheets. This model can be downloaded, both in Calc format (spreadsheets as part of the free LibbreOffice package, in which this model was created) and in Excel format (spreadsheets as part of Microsoft Office). Unlike spreadsheet models, which students of the Department of Zoology and Animal Ecology learn to build, this model does not even use the ability of spreadsheets to perform calculations based on formulas entered into the table. Here, the spreadsheet space is simply used as a sheet of paper: it indicates which calculations are performed. Let's consider this model in more detail. We simulate a population consisting of 20 individuals. Initially, there are 10 females and 10 males. In each cycle, each female produces 2 offspring (a female and a male). In the second cycle, one of the females that appears during reproduction turns out to be clonal (parthenogenetic). After reproduction in each cycle, the total population size is reduced to approximately 20 individuals. All groups of individuals are reduced in the same proportion. Since the population reduction is carried out according to rounding rules, sometimes there may be, for example, 19 or 21 individuals left. The same procedure is repeated 15 times. We see that parthenogenetic females reproduce faster, and are reduced in the same way as gonochoric individuals. At the beginning of the 16th cycle, only parthenogenetic females remain in the population... Pay attention to the graph in the lower left corner of the last slide of this lecture.
It shows the proportion of females, males, and clonal females in our model population. Why doesn't this happen in most gonochoric populations?
You won't believe it: there is still no answer to this question. There are a number of arguments that need to be considered. There are assumptions that contradict the prevailing view. This is one of the problems that would be very interesting to investigate using simulation modeling. To be continued...