Usova (2010) Determining Natural Mortality of Sexually Mature Water Frogs
An important paper for understanding the population ecology of frogs. Frogs are by no means precocious creatures... Usova E. E. Determining the natural mortality of sexually mature water frogs (Pelophylax esculentus complex; Amphibia, Ranidae) using skeletochronology // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. ...
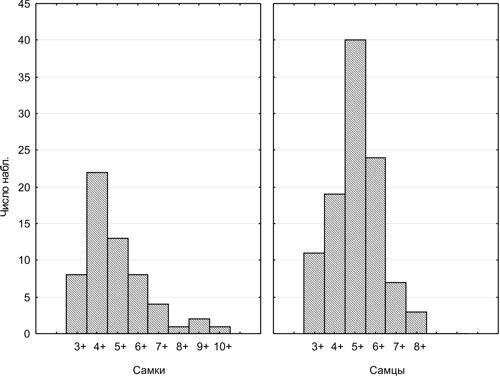
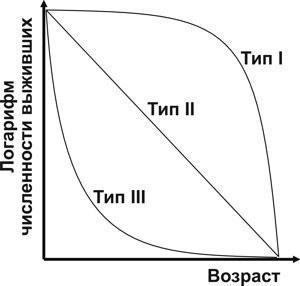
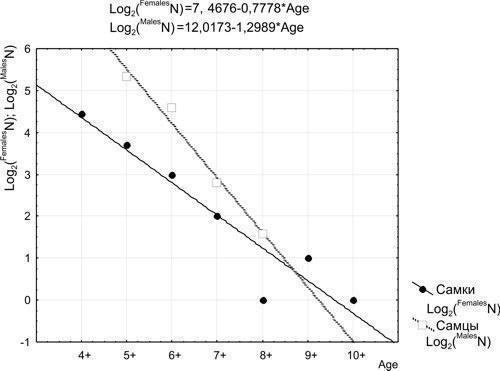
Usova E. E. Determining the natural mortality of sexually mature water frogs (Pelophylax esculentus complex; Amphibia, Ranidae) using skeletochronology // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. – 2010. – Issue 12 (No. 920). – P. 104–110. UDC (57.022+57.032):591.84:591.158.1:597.851 DETERMINING THE NATURAL MORTALITY OF SEXUALLY MATURE WATER FROGS (PELOPHYLAX ESCULENTUS COMPLEX; AMPHIBIA, RANIDAE) USING SKELETOCHRONOLOGY E. E. Usova Department of Biology, V. N. Karazin Kharkiv National University (Kharkiv, Ukraine) e_usova@mail.ru The pattern of mortality of water frogs (Pelophylax esculentus and Pelophylax ridibundus) from Kharkiv Oblast was studied using skeletochronological data from 146 individuals. Based on the age distribution of sexually mature frogs caught at spawning sites, it is hypothesised that the majority of females spawn for the first time at age 4+, and males at age 5+. Sexually mature water frogs are characterised by R. Pearl type II survival curve (mortality is age-independent). Keywords: Pelophylax esculentus (=Rana esculenta) complex, skeletochronology, sexual maturity, mortality. O. Ye. Usova. Determining the natural mortality of sexually mature water frogs (Pelophylax esculentus complex; Amphibia, Ranidae) using skeletochronology. The pattern of mortality of water frogs (Pelophylax esculentus and Pelophylax ridibundus) from Kharkiv Oblast was studied using skeletochronological age data from 146 individuals. Based on the age distribution of frogs caught at spawning sites, it is hypothesised that the majority of females spawn for the first time at age 4+, and males at age 5+. Sexually mature water frogs are characterised by R. Pearl type II survival curve (mortality is age-independent). Keywords: Pelophylax esculentus (=Rana esculenta) complex, skeletochronology, sexual maturity, mortality. E. E. Usova. Determining the level of mature water frogs (Pelophylax esculentus complex; Amphibia, Ranidae) natural mortality by use of skeletochronology. The rate of mortality of water frogs (Pelophylax esculentus and Pelophylax ridibundus) from the Kharkiv region was estimated on the basis of skeletochronology data on the age of 146 individuals. Taking into account the age distribution of the frogs caught in the spawning grounds, we suggest that females come on the first spawning by age 4+, and males – by age 5+. R. Pearl type II survival curve is characteristic for adult water frogs. Keywords: Pelophylax esculentus (=Rana esculenta) complex, skeletochronology, maturity, mortality. Introduction The hybridogenetic complex of water frogs, Pelophylax esculentus complex (=Rana esculenta complex), comprises two parental species, Pelophylax lessonae (Camerano, 1882) and Pelophylax ridibundus (Pallas, 1771), as well as various di-, tri-, and tetraploid hybrid forms (Plötner, 2005), which in the present work are treated under the name Pelophylax esculentus (Linnaeus, 1758). The coexistence of hybrids with parental species is maintained through hemiclonal inheritance (transmission by hybrids of certain genomes clonally, without recombination), which is of considerable theoretical interest. Some of the most complex hemiclonal populations of water frogs have been recorded in the upper Siverskyi Donets River basin within Kharkiv Oblast (Shabanov, Litvinchuk, 2010). To elucidate the mechanisms maintaining such complex biosystems, it is necessary to study the demographic processes occurring within them. The objective of this study is to determine the pattern of natural mortality inherent in sexually mature frogs using the results of skeletochronological age determination. Material Studied We determined the age of 163 sexually mature frogs: 59 females and 104 males (Table 1). Frogs were collected in Kharkiv Oblast from 2004 to 2009. Nearly half of them (73) were collected in the vicinity of the Biological Station of V. N. Karazin Kharkiv National University (Haidary village, Zmiyiv District, Kharkiv Oblast) and Homilshanski Forests National Nature Park. Species identity and genomic composition for the majority of specimens was determined by flow DNA cytometry at the Institute of Cytology of the Russian Academy of Sciences (St. Petersburg) by S. N. Litvinchuk and Yu. M. Rozanov. A portion of individuals (representatives of P. lessonae) were identified morphologically using a suite of characters (Lada, 1995; Shabanov et al., 2006). Table 1. Composition of the studied frog sample (L — genome of P. lessonae; R — genome of P. ridibundus) Genotype Females Males Of which from the vicinity of the Biological Station of KhNU and Homilshanski Forests NNP LL 8 9 - LLR 1 – - LR 18 73 47 LRR 5 8 11 RR 27 14 15 Methods of Skeletochronological Age Determination To determine the lifespan of individuals we employed the skeletochronological method (Smirina, 1983 and subsequent works; Usova, 2010). The third phalanx of the longest toe of the hind leg was cleaned of soft tissue, decalcified, and cross-sections of 20–22 μm thickness from the mid-diaphysis were obtained using a freezing microtome, stained with Ehrlich’s haematoxylin, and temporary preparations were photographed; measurements and counts of lines of arrested growth (LAGs) were subsequently performed from the photographs. These lines form during hibernation; their enumeration allows determination of the age of frogs. Since a certain portion of frog ontogeny occurs prior to the first hibernation, and age determination is generally performed on frogs that have emerged from hibernation, the age of a frog bearing, for example, 3 LAGs is designated as “3+”, i.e. “more than 3 years”. One of the principal difficulties in age determination arises from the fact that as new LAGs are added from the outer, periosteal surface of the bone, earlier ones are destroyed (resorbed) from within, from the endosteal cavity. In some cases periosteal rings are not completely destroyed, and their preserved fragments testify to their former existence. Even in cases of complete destruction of one or several lines, their number can be estimated with sufficient reliability. The diameter of the first LAGs differs less among individuals than the diameter of subsequent lines; although throughout life the differences in body size and LAG diameters in the phalanges between fast-growing and slow-growing frogs increase, the dimensions of young frogs and the diameters of their LAGs are sufficiently similar. By examining cross-sections of phalanges from sexually immature frogs, we established the typical dimensions of LAGs formed in the 1st and 2nd years of life. By measuring the endosteal diameter, one can estimate how many LAGs must have been accommodated within the cavity it now occupies. In the majority of sexually mature frogs, 2 LAGs were resorbed; in some individuals, 1 or 3. In some cases precise age determination is impossible. In such cases we indicated the most probable age, acknowledging that the true age may be one year less or more. Features of Statistical Processing of Recorded Age Distributions The results of age determination for the studied individuals are shown in Table 2. As can be seen, each category of frogs (e.g. individuals of a given sex, a given genotype, captured in a specific habitat in a particular year) is represented by a small number of individuals, insufficient for a well-founded assessment of their mortality pattern. At the same time, the entire dataset is sufficient for analysis by statistical methods. If the age distributions obtained for different categories of individuals do not differ significantly, they can be considered jointly. Table 2. Age distribution of the studied frog individuals Genotype Age of females Age of males 3+ 4+ 5+ 6+ 7+ 8+ 9+ 10+ 3+ 4+ 5+ 6+ 7+ LL – 3 3 1 1 – – – 3 5 1 – – LLR – – – 1 – – – – – – – – – LR 5 7 2 1 1 – 1 1 8 13 29 16 6 LRR – 2 1 2 – – – – – – 4 4 – RR 3 10 7 3 2 1 1 – – 1 6 4 1 First of all, for further analysis it is necessary to determine whether the data need to be processed separately for each sex, or whether they can be processed jointly. Using Statistica for Windows 6.5, we compared the observed age distributions of females and males against the expected distributions computed under the assumption of no association between sex and age structure (Table 3). Table 3. Example comparison of empirical and expected (based on the assumption of no association between the characteristics reflected in the rows and columns of the table) distributions. Age distributions of female and male frogs are shown. χ² = 16.07; degrees of freedom = 7; p = 0.0245; the difference between distributions is significant. Distributions Sex Age Total 3+ 4+ 5+ 6+ 7+ 8+ 9+ 10+ Empirical Females 8 22 13 8 4 1 2 1 59 Males 11 19 40 24 7 3 0 0 104 Total 19 41 53 32 11 4 2 1 163 Expected Females 6.88 14.84 19.18 11.58 3.98 1.45 0.72 0.36 59 Males 12.12 26.16 33.82 20.42 7.02 2.55 1.28 0.64 104 Total 19 41 53 32 11 4 2 1 163 Comparison of the empirical and expected distributions using the Pearson chi-squared (χ²) criterion allows assessment of the significance of the observed deviation of the distribution from random expectation. As is clear from Table 3, the age distributions of females and males differ significantly. This means that data for the two sexes must be processed separately. The next step is to determine whether data from individual genotypes can be considered jointly. For females, the difference between empirical and expected distributions is non-significant (p = 0.8). For males, such a difference is significant (p = 0.007), and is primarily attributable (see Table 2) to the difference between the age distributions of P. lessonae males and those of other genotypes. When P. lessonae males are excluded from consideration, the difference between the observed and expected distributions for the remaining genotypes becomes non-significant (p = 0.69). This means that data on the age distribution of P. ridibundus and various representatives of P. esculentus can be considered jointly, whereas P. lessonae is characterised by a different distribution, at least for males. The next step is to determine whether the age distribution data obtained from frogs from different populations can be considered jointly. As shown in Table 1, nearly half of the studied individuals (and more than half of the studied P. ridibundus and P. esculentus specimens) were obtained from a single metapopulation inhabiting the vicinity of the Biological Station of KhNU and Homilshanski Forests NNP. Comparing the age distributions of individuals from this metapopulation with the remaining studied representatives of P. ridibundus and P. esculentus from Kharkiv Oblast, we can confirm that the association between age and the geographic origin of frogs is non-significant (p = 0.43 for females and p = 0.29 for males). Comparison of the distributions of frogs caught in different years also provides grounds for their joint consideration. Consequently, all data pertaining to P. ridibundus and P. esculentus from Kharkiv Oblast can be considered jointly. Admittedly, joint consideration of data from frogs of different forms, captured in different habitats and at different times, represents a simplification of the actual picture. Such a simplification does not constitute a gross distortion, but it does afford an opportunity to assess the general character of the age distribution. Results and Discussion: The Character of Survival Curves for Sexually Mature Frogs The age distributions for all studied forms of water frogs, with the exception of P. lessonae, are shown in Fig. 1. [IMG_1] Fig. 1. Age distribution recorded for females and males of P. ridibundus and P. esculentus from Kharkiv Oblast (data for different genetic forms, different habitats, and different years of collection are considered jointly) In the analysis of the data presented in Fig. 1, it is noteworthy that for females the most numerous age class is 4+, and for males it is 5+. This likely indicates that it is at these ages that the majority of individuals of the respective sexes attend spawning grounds for the first time. For example, there are no grounds to assume that the population contains more females aged 4+ than aged 3+. The predominance of older age classes in the studied sample can be explained by the fact that it consists primarily of individuals caught at spawning sites. Presumably, a substantial proportion of three-year-old individuals do not attend spawning sites because they have not yet reached sexual maturity. From the age of four years onward, females regularly attend spawning grounds year after year, and the decline in their numbers reflects natural mortality. Since the reproductive products in anurans are formed in autumn, attending spawning grounds at age 4+ implies that sexual maturity was attained at age 3+. Analogously, it can be assumed that the first spawning for the majority of males occurs at age 5+. Therefore, owing to the absence of complete data on the relative abundance of younger age classes, mortality of females in our results can be analysed starting from the age of 4 years, and of males from the age of 5 years. For the interpretation of the results obtained, one may employ the classification of survival curve types proposed by the American classical population ecologist Raymond Pearl (Pearl, 1937). Predominantly late-life mortality (Fig. 2) is characteristic of populations with Pearl type I survival curves, for example, human populations in civilised society. This type of curve is termed the Drosophila type, referring to adult Drosophila in laboratory cultures. Type III, the oyster type, with mortality concentrated in early life, is characteristic of many animals with high fecundity. Age-independent mortality, which appears as a straight line on a graph with a logarithmic abundance scale, is characteristic of Pearl type II survival curves, the Hydra type. This type of survival curve is observed in organisms that have already passed through the most vulnerable stage of their ontogeny (Shabanov, Kravchenko, 2009). [IMG_2] Fig. 2. Types of survival curves according to R. Pearl (source of illustration: Shabanov, Kravchenko, 2009) To determine the type of mortality characteristic of sexually mature frogs, the abundance data for their age classes must be transformed to logarithmic form (Table 4). Table 4. Summary data for constructing survival curves. The two rightmost columns show only the data points used for constructing the curves.
Genotype
Females
Males
From the vicinity of the KhNU biological station and the Homilshanski Lisy National Nature Park
LL
8
9
-
LLR
1
–
-
LR
18
73
47
LRR
5
8
11
RR
27
14
15
Age Number of females (FemalesN) Number of males (MalesN) Logarithm of female abundance Log2(FemalesN) Logarithm of male abundance Log2(MalesN) 3+ 8 11 4+ 22 19 4.46 5+ 13 40 3.70 5.32 6+ 8 24 3.00 4.58 7+ 4 7 2.00 2.81 8+ 1 3 0.00 1.58 9+ 2 0 1.00 10+ 1 0 0.00 The survival curves constructed from the data in Table 4 are shown in Fig. 3. [IMG_3] Fig. 3. Survival curves for females and males of water frogs As can be seen from Fig. 3, the survival curves for sexually mature individuals of both sexes conform to Pearl type II (with the exception of the outlying data point corresponding to eight-year-old females). The regression line equations approximating the recorded age distributions are given at the top of the figure. In the regression equations obtained, the first term determines the level at which the curve lies (the abundance of the first age class under consideration, i.e. effectively the size of the studied sample), while the coefficient in the second term determines the slope of the curve (the annual mortality rate). On a logarithmic scale, the abundance of each successive age class is smaller by a fixed amount. Denoting Log2(FemalesNA) as the binary logarithm of female abundance at age A (given A > 4), and correspondingly Log2(FemalesNA+1) as the binary logarithm of their abundance at age A+1, we can assert on the basis of the data shown in Fig. 3 that Log2(FemalesNA+1) = Log2(FemalesNA) – 0.7778. Hence FemalesNA+1 = FemalesNA × 2−0.7778 and, finally, FemalesNA+1 = 0.58 × FemalesNA. Analogously, Log2(MalesNA+1) = Log2(MalesNA) – 1.2989, whence MalesNA+1 = MalesNA × 2−1.2989 and, ultimately, N(Male)A+1 = 0.406 × MalesNA. To simplify: of 10 sexually mature females, 6 will survive to the following year, and of 10 sexually mature males, 4 will survive. It should be emphasised that the age-independent mortality of frogs is not characteristic of their entire ontogeny, but only of its final stages. The result obtained pertains to the pooled sample comprising P. ridibundus and P. esculentus. Unfortunately, the available data on the age structure of P. lessonae are insufficient for analysis of their mortality pattern. Although the survival curves obtained are of an averaged character, smoothing out differences between habitats, frog forms, and years, they represent the first estimate of the mortality of water frogs in natural habitats. Such an estimate is, among other things, necessary for the mathematical modelling of processes in hemiclonal population systems of water frogs (Kravchenko, 2010). Further research may refine the established patterns and reveal the specifics of demographic processes characteristic of individual forms of these animals. Acknowledgements The author expresses sincere gratitude to S. N. Litvinchuk, Yu. M. Rozanov, and A. V. Korshunov for assistance in identifying the study material, and to D. A. Shabanov for participation in the interpretation and statistical processing of the results. This work was carried out with support from the Foundation for Fundamental Research of V. N. Karazin Kharkiv National University and the joint grant of the State Foundation for Fundamental Research of Ukraine and the Russian Foundation for Basic Research (Russia). References Kravchenko M. A. Mathematical modelling of the dynamics of various karyogenetic forms of water frogs in hemiclonal population systems // Proceedings of the 11th International Scientific-Practical Ecological Conference. — Belgorod, 2010. — P. 210. Lada G. A. Central European water frogs (hybridogenetic complex Rana esculenta): introduction to the problem // Flora and Fauna of the Central Chernozem Region. — Tambov, 1995. — P. 88–109. Smirina E. M. In vivo age determination and retrospective body size estimation in the common toad (Bufo bufo) // Zoologichesky Zhurnal. — 1983. — Vol. 63, No. 3. — P. 437–444. Usova E. E. Age structure of representatives of the hybridogenetic complex of water frogs in Kharkiv Oblast // Proceedings of the 11th International Scientific-Practical Ecological Conference. — Belgorod, 2010. — P. 131. Shabanov D. A., Zinenko A. I., Korshunov A. V., Kravchenko M. A., Mazepa G. A. Study of population systems of water frogs (Rana esculenta complex) in Kharkiv Oblast: history, current state, and prospects // Visnyk of V. N. Karazin Kharkiv National University. Series: Biology. — 2006. — Issue 3 (No. 729). — P. 208–220. Shabanov D. A., Kravchenko M. A. Study materials for the course in general ecology with foundations of environmental science and human ecology. — Kharkiv: V. N. Karazin Kharkiv National University, 2009. — 292 p. Shabanov D. A., Litvinchuk S. N. Water frogs: life without rules or a special mode of evolution? // Priroda. — 2010. — No. 3. — P. 29–36. Pearl R. On biological principles affecting populations: human and other // Amer. Natur. — 1937. — Vol. 71, N 1. — P. 50–68. Plötner J. Die westpaläarktischen Wasserfroesche. Bielefeld: Laurenti-Verlag, 2005. – 161 S.
Genotype
Вік самок
Вік самців
3+
4+
5+
6+
7+
8+
9+
10+
3+
4+
5+
6+
7+
LL
–
3
3
1
1
–
–
–
3
5
1
–
–
LLR
–
–
–
1
–
–
–
–
–
–
–
–
–
LR
5
7
2
1
1
–
1
1
8
13
29
16
6
LRR
–
2
1
2
–
–
–
–
–
–
4
4
–
RR
3
10
7
3
2
1
1