Beast-Pegasopher. Column in ComputerreOnline #61
Based on the assumption of evolutionary economy, we reconstruct its pathways using molecular data. And we become convinced of the whimsical, uneconomical course of changes in organismal structure.
Can you imagine Bosch’s “Garden of Earthly Delights”? This is the first panel of the triptych known by that title. The second and especially the third panels seem to me manifestations of mental illness, while I have long loved the first. In my childhood and youth a copy of this painting hung in my room and influenced my attitude toward animal diversity.
In the lower half of the painting the Creator brings Eve to Adam for the first time, but the main content of the canvas is not this event. Its overwhelming part is occupied by a whimsical landscape and a multitude of creatures. Among them are both quite realistic, slightly strange, and completely unusual ones, sometimes resembling each other, sometimes entirely different. I have always felt that the humans in this picture are less real than all the other animals.
“The Garden of Earthly Delights” by Hieronymus Bosch
The very possibility of comparing different organisms with each other stems from their fundamental similarity, manifested through their differences. Here it is, biodiversity! Why do these beings differ from one another? For Bosch and his contemporaries the answer was clear—it fits their design. The reason for their similarities and differences was seen in the same way as the reason for the similarities and differences of different beats of different instruments in a large and complex symphony.
Our methods of animal systematics go back to Carl Linnaeus, the Swedish genius of the 18th century. Linnaeus did not invent hierarchical classification, nor did he invent binary nomenclature (the designation of a species by a generic name and a specific epithet), but he was the first to build the whole system on simple and universal principles. In general, binary nomenclature was an application of Aristotelian logic to the description of animal diversity. Such names can be used for both living and non‑living objects. Tables can be writing or dining, toads—green and gray… The classic systematist tormented his housewife by achieving a precise distribution of socks and stockings in the linen closet according to a special system.
Linnaeus understood that systems could be different and proposed their division into artificial and natural. In school textbooks you may read that natural systems are those reflecting the kinship of organisms. This is a later reinterpretation that does not correspond to Linnaean approach. Artificial systems, according to Linnaeus, are convenient for identifying organisms but reflect only their individual features. In contrast, a natural system should reflect the essential similarity of organisms (if you wish, it can be interpreted as the Creator’s design).
Darwin was not the first to realize that similarity of organisms is a consequence of their kinship, but he was the first to grasp many implications of this fact. For example, it became clear that by examining the structures of similarity and difference in observed animals one can understand the pathways of their evolution. Incidentally, this implied that the diversity of organisms, in an important respect, differs from the diversity of tables, stockings and socks. They are linked by common origin and relate to each other like branches of a single tree. Hence, a system of organisms, as a way of describing their diversity, must fundamentally differ from the system of most objects. It was only after the Darwinian revolution that the natural system came to be understood as the one reflecting the kinship of the classified objects.
At first, building evolutionary trees required only a flight of fancy (especially in this field the German Darwinist Ernst Haeckel stood out), but over time biologists wanted more evidence. Then a vicious circle in the proofs was recognized. Note: we established that similarity is a consequence of kinship. And how do we judge kinship? On the basis of similarity. That does not look very good.
The best way out of this logical circle was offered by cladistics (more precisely, phylogenetic systematics), a school founded by the German entomologist Willi Hennig. Hennig step‑by‑step formalized a procedure in which, based on individual manifestations of similarity, kinship is reconstructed. Supporters of Hennig’s hypothesis about the most probable evolutionary history of a group build it on certain principles. The most debated are two of them. Evolution proceeds by the most economical path. Branching of the evolutionary tree occurs as follows: an old species ceases to exist and, in its place, two (exactly two!) new species appear.
Why does cladistics adopt such ambiguous principles? In a sense it looks for lost keys not where they were lost, but under the lamp, where it is more convenient to search. If evolution had proceeded that way, it would be easy for us to reconstruct it. So, let us assume that evolution did proceed that way…
You can read more about the relationship of the three schools in systematics and the reconstruction of evolutionary trajectories (evolutionary, numerical and phylogenetic) here, in my old text written several years ago for “Kompyuterr”. In this column, however, something else matters to me.
When using one set of approaches you have to assume one form of the evolutionary tree, when using another—a different one. If a system of organisms tries to reflect their kinship, then reshaping ideas about evolutionary pathways necessitates reshaping the system! This is one of the reasons that make debates about evolutionary trajectories especially heated.
Naturally, at first cladistic supporters still worked with classical organismal characters, primarily structural features. At this stage they broke a lot of wood. The fact is that in the evolution of those characters linked to structure and function, parallel development is observed everywhere. The requirement to choose, from all possible developmental trajectories, the most economical one (the one that entails the minimal number of evolutionary changes) leads, under such conditions, to frequent errors.
May I give an exaggerated example? Given: two old men. Required: determine how they originated. There are two possible solutions. Option A. Each of these old men was once a child, then they independently grew up and aged, passing through similar stages of life in parallel. Option B. There was one child who grew up and aged, and then split into two old men.
The paradox is that if, following cladistic methodology, we look for the option with the minimal number of transformations, we must choose option B.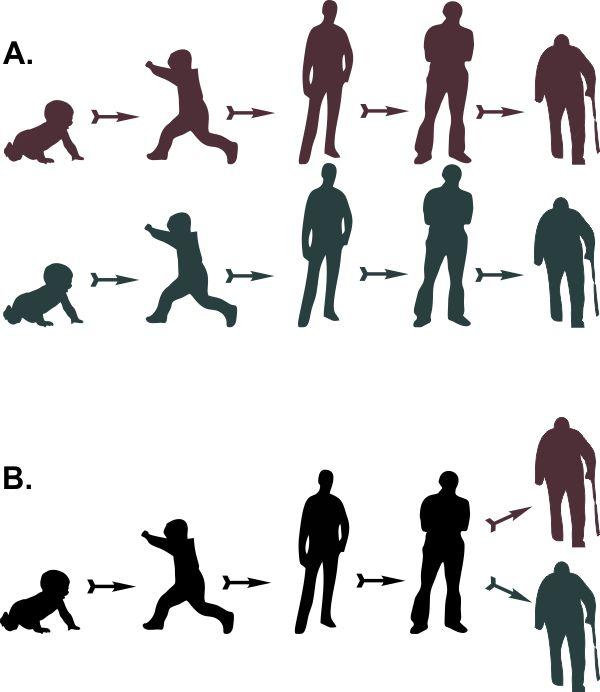
Which origin scenario for the two old men is more economical (requires fewer independent events)? Of course, option B! It involves 5 transitions, whereas option A requires a full 8.
Does this mean that a cladistic systematist will really insist on the second option? Of course not. By expanding the number of characters considered in our analysis, we will find that “splitting” a man requires many additional assumptions. When reconstructing ontogeny (the life path), no one reasons as in the given example.
However, in reconstructing phylogeny, the evolutionary history, cladists often insist on analogues of option B from our exaggerated example. Without going into details (more here), I will say that the colonisation of land by vertebrates could have occurred either through adaptation of several parallel branches to terrestrial life, or through a single branch. I find the latter (based on analysis of minor characters that may seem insignificant) better justified. Cladists who deem it necessary to choose the more economical hypotheses, however, argue for option B.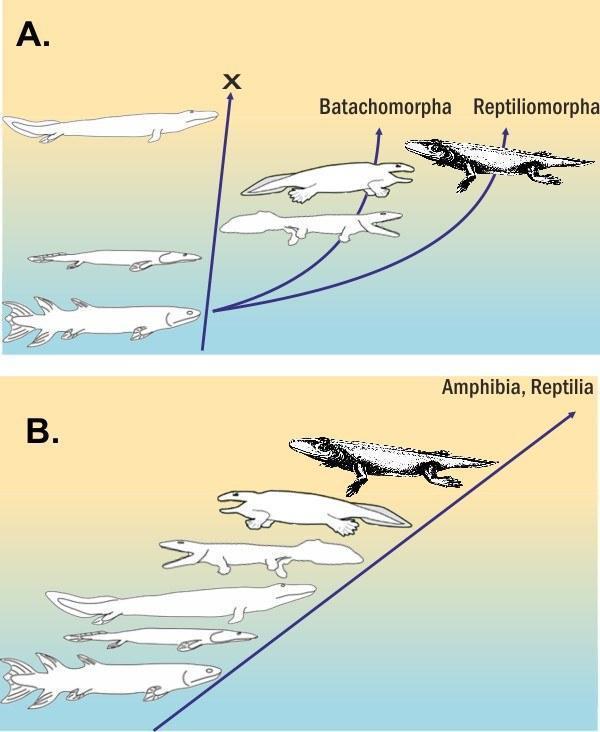
This illustration is analogous to the previous one, but here the choice between the proposed hypotheses is not so trivial. How did vertebrates colonise land: with a single stem that later split into several groups, or with several parallel stems?
But at the end of the 20th century a revolution occurred. The development of methods for reading and comparing genetic texts of different organisms gave systematists and evolutionists many new characters. And it turned out that many of these characters behave exactly as required for cladistic analysis!
Today the technology of “tracing” past evolution through genetic texts has reached a high level of development. Studies performed on genetic material often confirm each other. But it is also important that they frequently overturn established ideas about the course of evolution based on comparisons of form and function.
A detailed discussion of this upheaval in systematics would take considerable time, and a talk about the problems of using new organismal systems in biology teaching would be a separate complaint. Here I will give only a few examples concerning mammalian systematics.
Could one, based on classical characters of modern animals, have guessed that the walrus is a closer relative of the dog than of the cat? In modern systematics the order Pinnipedia has been eliminated, because seals and their relatives turned out to be just one branch of the order Carnivora. The ancestors of cats split from the ancestors of dogs before the latter split from the ancestors of seals.
And which animals turned out to be the closest relatives of carnivores? Reptiles (pangolins): scaly Afro‑Asian ant and termite hunters. For the group uniting carnivores and reptiles, the name Ferae has been proposed (literally “wild ones”).
The next closest relatives of carnivores are the odd-toed ungulates: horses, rhinoceroses and tapirs. The wolf is closer to the horse than to the cow. Yet cetaceans are so close to even‑toed ungulates that they should be merged into a single order!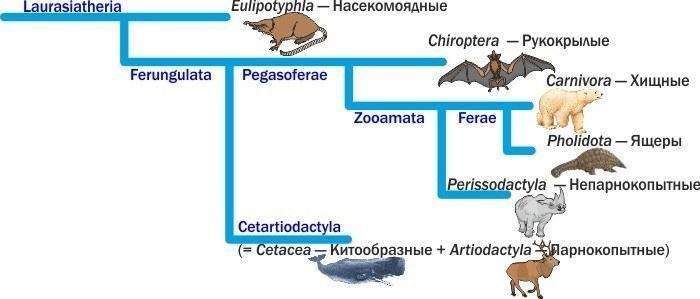
Fragment of the mammalian evolutionary tree (according to Nishihara et al., 2006). Order names are in black, clades (individual branches) in blue
If the examples I gave surprised you, the root of that surprise lies in the whimsical character of evolution, which would be hard to predict from external traits alone. Nevertheless, conjectures have been varied… Here Dawkins marvels at Haeckel’s insight, who “… classified hippopotamuses as a sister group to whales: in his view, hippos were more closely related to whales than to pigs, and all three were more closely linked to each other than to cows” (Epilogue to “The Hippopotamus Story” from “The Ancestor’s Tale”).
Thus, based on the assumption of economical evolution, we reconstruct its pathways from molecular data. And we become convinced of the whimsical, far from economical, course of the evolution of organismal structures and functions. But thanks to this situation we now have the (still largely untapped) possibility to try to understand the logic of evolution of vital traits of living beings. That is the main conclusion of the column.
And why is it called that? In one of the molecular‑data‑based versions, a group is distinguished that includes the orders Carnivora, Reptilia, Perissodactyla and Chiroptera (the inclusion of chiropterans is disputed; other interpretations exist). What to call this group? Its discoverers proposed the word Pegasoferae. Well, it is a decent way to unite horses, bats and carnivores in a single metaphor!
Ancient Greeks mistakenly thought that Pegasus should have bird wings with feathers. Recent scientific achievements show that they should have had a leathery membrane between the digits, like bats. And, of course, a Pegasus (being a quadruped) should have exactly four limbs, not six as in old, still imperfect reconstructions. When depicting the pegasifer beast, the quintessence of the Pegasoferae group, one only needs to add a wolf’s head and a pangolin’s tail to the reconstruction.
How do you like this reconstruction of the pegasifer beast? Thanks to Dmytro Bukhanovsky for helping create it. You understand that this is a joke, right?
Just as in Bosch’s time, the diversity of organisms remains for us a source of amazed admiration…