
Ecology: the biology of interactions. IV-15. Ecological strategies
Populations of certain species spend the majority of their time in the r‑phase. In such species, the highest reproductive value is possessed by individuals capable of rapid reproduction and colonizing unoccupied habitats with their offspring. In other words, during this phase, selection will favor an increase in the parameter r — reproductive...
IV-15. Ecological strategies
How to determine the value of an individual for a population?
"Natural selection recognizes only one kind of ‘currency’—successful offspring" (E. Pianka, 1981).
We have said that a population is a potentially immortal entity composed of mortal individuals. To sustain the existence of the population, an individual must survive itself and leave descendants that can also survive. Note the duality of this task. Likely, the greatest chance of survival belongs to an individual that does not expend resources and the energy obtained from them on offspring production at all. But after a short time such an individual will disappear from the population without offspring. At the opposite "pole" is a hypothetical individual that, immediately after appearance, directs all its energy to producing offspring. Such a creature will die itself and, if its offspring inherit the same inefficient resource allocation, will give rise to offspring that have no chance of survival.
Thus, the greatest value for the population should belong to an individual that combines the costs of its own survival and offspring production in an optimal combination. This combination can be evaluated. For this, one must calculate under which combination in these conditions the individual will make the greatest possible contribution to the future generation. The measure used for this in mathematical population biology is called reproductive value. Reproductive value is a generalized measure of survival and fecundity that accounts for the relative contribution of an organism to future generations.
"It is easy to describe a hypothetical organism that has all the traits needed to achieve high reproductive value. It reproduces almost immediately after birth, produces numerous, large, protected offspring that it cares for; reproduces many times and frequently over a long life; it wins in competitive struggle, avoids predators and easily obtains food. Describing such a creature is easy, but imagining it is difficult..." (M. Bigon et al., 1989).
You understand that this impossibility stems from the contradictory tasks of self‑maintenance and reproduction (Fig. IV-15.1). One of the first to realize this was the English philosopher Herbert Spencer in 1870, who spoke of the alternative of an organism supporting its own existence and continuing itself in descendants. In modern terms, these parameters are linked by negative correlations, a relationship in which improvement in one parameter must be accompanied by deterioration in the other.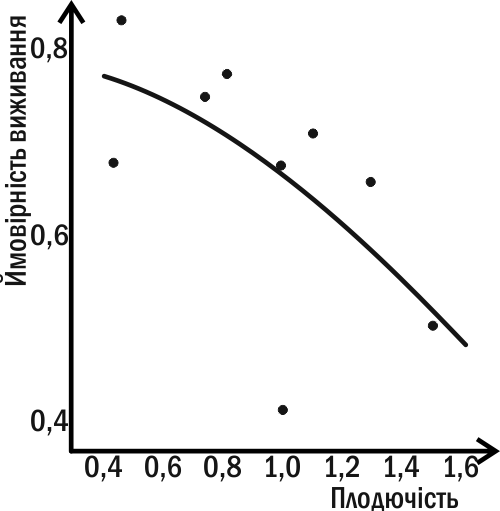
Fig. IV-15.1. In Asplanchna rotifers, survival chances decrease with increasing fecundity (Pianka, 1981).
Different species (and different populations) redistribute energy between self‑maintenance and reproduction in various ways. One can speak of a species strategy expressed in how members of the species acquire resources and how they expend them. Only a strategy in which individuals obtain enough energy to grow, reproduce, and compensate for all losses due to predator activity and various misfortunes can be successful.
Traits related to different adaptive strategies may be linked by a trade‑off relationship, i.e., insurmountable negative correlations (either‑or relationship). Thus, trade‑offs relate the number of offspring and their survival, growth rate and stress resistance, etc. American ecologists R. MacArthur and E. Wilson described in 1967 two types of species strategies that result from two different types of selection and are linked by trade‑offs. The designations of these strategies (r‑ and K‑) are taken from the logistic equation.
According to the logistic model, population growth can be divided into two phases: accelerated and decelerated growth (Fig. IV-15.2). While N is small, population increase is mainly driven by the multiplier rN, and growth accelerates. In this phase (the r‑phase) population growth accelerates, and its size is higher the greater the individuals' reproductive capacity. When N becomes sufficiently large, the multiplier (K‑N)/K begins to dominate population size. In this phase (the K‑phase) growth slows. When N=K, (K‑N)/K=0 and population growth ceases. In the K‑phase, population size is higher the larger the parameter K, which is higher the more competitive the individual.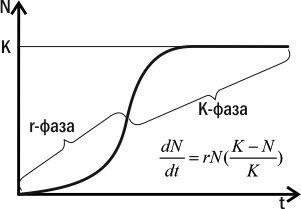
Fig. IV-15.2. r‑ and K‑phases of population growth according to the logistic model
It can be assumed that populations of some species spend most of their time in the r‑phase. In such species, the highest reproductive value belongs to individuals capable of rapid reproduction and of colonizing unoccupied habitats with their offspring. In other words, during this phase selection will favor an increase in the parameter r—reproductive potential. This selection is called r‑selection, and the resulting species are r‑strategists.
In species whose populations spend most of their time in the K‑phase, the situation is opposite. The maximum reproductive value in these populations will belong to individuals that are so competitive that they can obtain their share of resources even under scarcity; only then can they reproduce and contribute to the next generation. A population composed of such individuals will have a higher K—environmental carrying capacity—than one composed of individuals that cannot "fight" for resources. At this stage K‑selection operates, producing K‑strategists. K‑selection aims to increase the investment in the development of each individual and its competitiveness.
Transitions between these strategies are possible, but they are intermediate rather than combining typical manifestations of both forms.
"One cannot be both salad and cactus" (E. Pianka).
An important factor for determining whether r‑ or K‑selection will act on a species is the dynamics of available resource quantity and the intensity of competition for it. With a sharp non‑selective reduction in population size caused by resource shortage due to external factors, r‑strategists gain an advantage; under competitive struggle for limited resources, K‑strategists prevail.
Choosing between an r‑strategy (increasing fecundity) and a K‑strategy (increasing competitiveness) is fairly simple, yet it touches many organismal parameters and life cycles. Compare these strategies in their typical form (Table IV-15.1).
Table IV-15.1. Features of r‑ and K‑selection and strategies
|
Early Maturation |
Table IV-15.1. Features of r‑ and K‑selection and strategies |
Characteristics |
|
r‑selection and r‑strategists |
K‑selection and K‑strategists |
Climate |
|
L | Life‑span | Life‑span |
Stable, predictable |
Mortality |
|
Catastrophic, density‑independent |
Competition‑induced, density‑dependent |
Mortality curve |
|
Usually type III |
Usually type I or II |
Population size |
|
Variable, unbalanced |
Stable, near environmental carrying capacity |
Free resources |
|
Appearance of free resources, filling an "ecological vacuum" |
Free resources are rare, occupied by competitors |
Intra‑ and interspecific competition |
|
Weak |
Intense |
Body size |
|
According to thermodynamic laws, the work of a system equals the decrease in free energy, which is directly related to internal energy — the sum of the energies of all structural elements. The specific manifestations of these regularities are highly diverse. Biological species specialize and lose evolutionary plasticity. Spiritual life develops from the omnipotence of the individual's magical will to dependence on impersonal will, fate, and further to voluntary spiritual enslavement. Rituals obscure faith; canonized beauty loses its appeal; morality, enshrined in law and protected by law enforcement agencies, exits the sphere of ethics, since a moral act is only possible in a situation of free choice. Conjugal love gives way to conjugal duties. In personal life, the romantic period is limited to early youth, replaced by a conservative phase, the prelude to death. |
Relatively large |
Development |
|
Fast |
Early |
Middle |
|
Early |
Late |
Reproductive rate |
|
High |
Low |
Reproduction over lifespan |
|
Often single‑event |
Multiple events |
Offspring per brood |
|
Many |
Reproductive rate |
Late |
|
**Viability parameters** |
High |
Lifespan |
|
Short |
Long |
Adaptations |
|
Primitive |
Table 3.13.1. Example of ecological pyramids for a hypothetical trophic chain |
Optimized for |
IV-15. Ecological strategies
How to determine the value of an individual for a population?
"Natural selection recognizes only one kind of ‘currency’—successful offspring" (E. Pianka, 1981).
We have said that a population is a potentially immortal entity composed of mortal individuals. To sustain the existence of the population, an individual must survive itself and leave descendants that can also survive. Note the duality of this task. Likely, the greatest chance of survival belongs to an individual that does not expend resources and the energy obtained from them on offspring production at all. But after a short time such an individual will disappear from the population without offspring. At the opposite "pole" is a hypothetical individual that, immediately after appearance, directs all its energy to producing offspring. Such a creature will die itself and, if its offspring inherit the same inefficient resource allocation, will give rise to offspring that have no chance of survival.
Thus, the greatest value for the population should belong to an individual that combines the costs of its own survival and offspring production in an optimal combination. This combination can be evaluated. For this, one must calculate under which combination in these conditions the individual will make the greatest possible contribution to the future generation. The measure used for this in mathematical population biology is called reproductive value. Reproductive value is a generalized measure of survival and fecundity that accounts for the relative contribution of an organism to future generations.
"It is easy to describe a hypothetical organism that has all the traits needed to achieve high reproductive value. It reproduces almost immediately after birth, produces numerous, large, protected offspring that it cares for; reproduces many times and frequently over a long life; it wins in competitive struggle, avoids predators and easily obtains food. Describing such a creature is easy, but imagining it is difficult..." (M. Bigon et al., 1989).
You understand that this impossibility stems from the contradictory tasks of self‑maintenance and reproduction (Fig. IV-15.1). One of the first to realize this was the English philosopher Herbert Spencer in 1870, who spoke of the alternative of an organism supporting its own existence and continuing itself in descendants. In modern terms, these parameters are linked by negative correlations, a relationship in which improvement in one parameter must be accompanied by deterioration in the other.
Fig. IV-15.1. In Asplanchna rotifers, survival chances decrease with increasing fecundity (Pianka, 1981).
Different species (and different populations) redistribute energy between self‑maintenance and reproduction in various ways. One can speak of a species strategy expressed in how members of the species acquire resources and how they expend them. Only a strategy in which individuals obtain enough energy to grow, reproduce, and compensate for all losses due to predator activity and various misfortunes can be successful.
Traits related to different adaptive strategies may be linked by a trade‑off relationship, i.e., insurmountable negative correlations (either‑or relationship). Thus, trade‑offs relate the number of offspring and their survival, growth rate and stress resistance, etc. American ecologists R. MacArthur and E. Wilson described in 1967 two types of species strategies that result from two different types of selection and are linked by trade‑offs. The designations of these strategies (r‑ and K‑) are taken from the logistic equation.
According to the logistic model, population growth can be divided into two phases: accelerated and decelerated growth (Fig. IV-15.2). While N is small, population increase is mainly driven by the multiplier rN, and growth accelerates. In this phase (the r‑phase) population growth accelerates, and its size is higher the greater the individuals' reproductive capacity. When N becomes sufficiently large, the multiplier (K‑N)/K begins to dominate population size. In this phase (the K‑phase) growth slows. When N=K, (K‑N)/K=0 and population growth ceases. In the K‑phase, population size is higher the larger the parameter K, which is higher the more competitive the individual.
Fig. IV-15.2. r‑ and K‑phases of population growth according to the logistic model
It can be assumed that populations of some species spend most of their time in the r‑phase. In such species, the highest reproductive value belongs to individuals capable of rapid reproduction and of colonizing unoccupied habitats with their offspring. In other words, during this phase selection will favor an increase in the parameter r—reproductive potential. This selection is called r‑selection, and the resulting species are r‑strategists.
In species whose populations spend most of their time in the K‑phase, the situation is opposite. The maximum reproductive value in these populations will belong to individuals that are so competitive that they can obtain their share of resources even under scarcity; only then can they reproduce and contribute to the next generation. A population composed of such individuals will have a higher K—environmental carrying capacity—than one composed of individuals that cannot "fight" for resources. At this stage K‑selection operates, producing K‑strategists. K‑selection aims to increase the investment in the development of each individual and its competitiveness.
Transitions between these strategies are possible, but they are intermediate rather than combining typical manifestations of both forms.
"One cannot be both salad and cactus" (E. Pianka).
An important factor for determining whether r‑ or K‑selection will act on a species is the dynamics of available resource quantity and the intensity of competition for it. With a sharp non‑selective reduction in population size caused by resource shortage due to external factors, r‑strategists gain an advantage; under competitive struggle for limited resources, K‑strategists prevail.
Choosing between an r‑strategy (increasing fecundity) and a K‑strategy (increasing competitiveness) is fairly simple, yet it touches many organismal parameters and life cycles. Compare these strategies in their typical form (Table IV-15.1).
Table IV-15.1. Features of r‑ and K‑selection and strategies
Climate
Variable, unpredictable
Stable, predictable
Mortality
Catastrophic, density‑independent
Competition‑induced, density‑dependent
Mortality curve
Usually type III
Usually type I or II
Population size
Variable, unbalanced
Stable, near environmental carrying capacity
Free resources
Appearance of free resources, filling an "ecological vacuum"
Free resources are rare, occupied by competitors
Intra‑ and interspecific competition
Weak
Intense
Body size
Relatively small
Relatively large
Development
Fast
Slow
Sexual maturity
Early
Late
Reproductive rate
High
Low
Reproduction over lifespan
Often single‑event
Multiple events
Offspring per brood
Many
Few, often one
Resource per offspring
Low
High
Lifespan
Short
Long
Adaptations
Primitive
Advanced
Optimized for
Productivity
Efficiency
It may seem odd that r‑strategists typically have single‑event reproduction, while K‑strategists have multiple events. This can be illustrated with an example. Imagine mice colonizing a granary with abundant resources and no competition. Consider the strategies of two species.
Species #1: Sexual maturity at 3 months, 10 offspring per brood, female lives one year and can reproduce every three months.
Species #2: Sexual maturity at 3 months, 15 offspring per brood, female dies from exhaustion after raising them.
In the first case, after three months, 10 offspring and their parents (12 individuals total) will reproduce; in the second case, 15 offspring will. The higher rate of occupying free resources will be provided by the second species. A typical r‑strategy forces individuals to reproduce as early and as intensively as possible, often limiting them to a single breeding season.
Conversely, it is easy to understand why typical K‑strategists reproduce many times. In a competitive environment, only the offspring that received substantial resource investment will survive. Moreover, an adult must allocate considerable energy to its own maintenance and development to survive and reproduce. Thus, in the extreme case K‑strategists produce one offspring at a time (e.g., elephants, whales, and, in most cases, humans). But however perfect these animals are, the parental pair will eventually die. For the population not to cease, the pair must leave more than two surviving offspring. Hence, repeated reproduction is a necessary condition for K‑strategist survival.
In 1935 Soviet botanist L.G. Ramenko identified three groups of plants, which he called ceno‑types (the concept of strategies had not yet been formed): violents, patients, and exploiters. In 1979 English ecologist J. Grime rediscovered these groups (under other names) (Fig. IV-15.3). The strategies are as follows.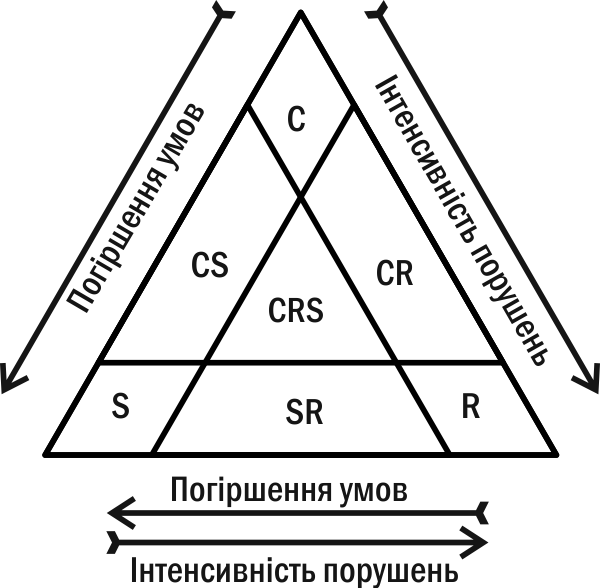
Fig. IV-15.3. "Grime's triangle"—classification of species strategies
– Type C (competitor), violents per Ramenko; allocate most energy to maintaining adult life, dominate stable communities. Among plants, this type includes trees, shrubs, or robust grasses (e.g., oak, reed).
– Type S (stress‑tolerant), patients per Ramenko; through special adaptations endure adverse conditions; use resources where competition is minimal. Usually slow‑growing organisms (e.g., Sphagnum, lichens).
– Type R (from Latin ruderis, ruderal), exploiters per Ramenko; replace violents in disturbed communities or exploit temporarily unused resources. Among plants, these are annuals or biennials that produce abundant seeds. Such seeds form a seed bank in the soil or disperse over long distances (e.g., dandelion, fireweed). This allows them to wait for resource release or quickly colonize open patches.
Many species can combine different strategy types. Pine belongs to the CS category, as it grows well on poor sandy soils. Stinging nettle is a CR strategist, dominating disturbed sites.
Species strategy can be plastic. The chestnut oak is a violents in broad‑leaf forests and a patient in the southern steppe. The Japanese bonsai technique (cultivating dwarf trees in pots) can be seen as converting violents into patients.
An interesting task is to compare the strategies of MacArthur‑Wilson with those of Ramenko‑Grime. Clearly, r‑strategists correspond to R‑type organisms, exploiters. K‑strategists correspond not only to C‑type organisms, violents, but also to S‑type, patients. Violents maximize competitiveness (and environmental capacity) under intense competition for readily consumable resources, while patients do so under constrained resource availability. In other words, the problems solved by an oak competing for light in a dense forest and a fern surviving in dim cave light share much: the need to optimize resource consumption and improve individual adaptation.