
Ecology: biology of interactions. IV-19. (supplement) Sex must be impossible! Why does it exist?
«The core» of the complex of phenomena, which includes the presence of sexes, the difference between them, and sexual acts, is precisely sexual recombination, the formation of new gene combinations that ensures the probabilistic combination of hereditary traits of parental individuals as a result of fertilization — their fusion...
We have mentioned several types of multiplication. For further discussion, we need to propose their typology. The typology we propose is shown in Table IV-19.1. It was developed (Shabanov, 2013, 2015) for discussing the features of hemiclonal population systems of the hybridogenetic complex of green frogs, and this explains the attention to Type IV of multiplication. Undoubtedly, the most ancient type of population multiplication is Ia, clonal with horizontal transfer of genetic information; this type is characteristic of prokaryotes. Probably, in most cases, representatives of Type Ib (clonal without horizontal transfer) are organisms that once had sexual recombination and then lost it. To which type the first organisms with sexual reproduction belonged is a question that can be discussed; perhaps it was a type not shown in Table IV-19.1 (monomorphic organisms that produced uniform gametes). In any case, it is clear that the origin of sexual recombination could not have been connected with Type IIIb; for significant ecological differences between sexes to arise, separate sexes had to already exist, that is, Type IIIb populations had to have developed from Type IIIa populations.
|
Biosystems |
Populations |
Hemiclonal population systems |
||||
|
Clonal populations |
Mendelian populations |
|||||
|
with horizontal transfer |
without horizontal transfer |
hermaphroditic |
dioecious |
|||
|
without sexual dimorphism |
with sexual dimorphism |
|||||
|
Ia |
Ib |
II |
IIIa |
IIIb |
IV |
|
 |
 |
 |
 |
 |
 |
|
|
Individuals |
Monomorphic |
Monomorphic, two sexes |
Two sexes that differ significantly |
Two sexes, different forms |
||
|
II |
IIIa |
IIIb |
Essentially linear (in diploid hybrids) |
|||
|
Genetic |
None; within a clone, individuals are not unique |
The vast majority of individuals are genetically unique |
Non-heritable (in hemiclones and clones) |
|||
|
Uniqueness of individual |
None; a set of clones |
Exists; a pool of genes that recombine |
Gene pool of the parental species + a set of hemiclones and clones |
|||
|
"Double |
None; all individuals can directly produce offspring |
Exists; only females can directly produce offspring |
||||
|
Sexual |
None |
Relatively less effective |
Relatively more effective |
Calymmian |
||
|
Intraspecific |
Non-homologous, sometimes — homologous |
None |
Homologous (between corresponding chromosomes and loci due to crossover) |
Exists in triploid hybrids between chromosomes of the parental species that are represented by two copies |
||
|
Interspecific |
Non-homologous |
None (due to sterility of interspecific hybrids, which is caused by failures in recognition of homologous chromosomes by each other during meiosis) |
Homeologous (between corresponding chromosomes of different species) |
|||
| Existence of "true" species |
Doubtful | Exist; in typical cases, species are communities with a certain gene pool that are genetically isolated from other species | Doubtful | |||
IV-19. (supplement) Sex Should Be Impossible! Why Does It Exist?
(in progress)
The evolution of sex is the most complex problem of evolutionary biology.
John Maynard Smith
Sex is the main challenge of modern evolutionary theory... the queen of problems of evolutionary biology.
Graham Bell
One of the strange features of sex is that, according to prevailing scientific understanding, it is impossible. To understand this, we must first consider what exactly we will mean by the term "sex" in this discussion; for this, we will have to unravel some not-simple terminological complexities. Let us distinguish three different understandings of "sex":
— sex in the everyday sense — sexual intercourse, "having sex"; this is the realization in ordinary life of the mysterious phenomenon connected with the reproduction of new genotypes and multiplication;
— sex sensu stricto (in the strict sense) — sex, the belonging of an individual to one of two categories that differ in the sex cells (gametes) they can produce; as is known, females produce relatively large gametes, egg cells, males — small sperm; the concept of "sex" in this sense should not be confused with "gender," which refers to a cultural construct describing notions about the characteristics and roles of representatives of different sexes;
— sex sensu lato (in the broad sense) — sexual recombination, the formation of new genotypes that combine the genetic information of parental individuals in new random combinations.
"By sex in eukaryotes we mean the more or less regular alternation of meiosis and syngamy. A natural consequence of this is the alternation of haploid and diploid phases in the life cycle."
John Maynard Smith, Eörs Szathmáry. The Major Transitions in Evolution. 1995.
The third of the listed understandings of "sex," and the meaning referred to in the quoted passage, is the same. The "core" of the complex of phenomena that includes the existence of sexes, the differences between them, and sexual acts, is precisely sexual recombination, the formation of new combinations of genes as a result of combining the hereditary traits of parental individuals through fertilization — the fusion of their gametes (syngamy). Gamete fusion leads to a doubling of the amount of hereditary information; therefore, in a life cycle where fertilization occurs, there must also be a two-fold reduction in the amount of hereditary information. Such reduction is probabilistic, and it is ensured by meiosis. Mitoses ensure clonal transmission of genetic information, meiosis in combination with fertilization — sexual.
Is sex a form of reproduction or multiplication? In a number of cases, sex is not connected with an increase in the number of individuals (actual reproduction); it is only the cause of the formation of new (multiplication) genotypes. We consider it appropriate to use the word "reproduction" when referring to organisms, and to use "multiplication" when discussing processes that occur in populations (at a level higher than the organismal) and with genotypes (at a lower level). Of course, when referring to multiplication, the concepts "sexual" and "asexual/clonal" are not used to describe the attractiveness or unattractiveness of individuals, but to indicate how new genotypes are formed.
Of course, the paradoxical nature of sex makes sense to discuss precisely with the described understanding of this concept (though the existence of sexes is also paradoxical!). In the case of two individuals of different sexes that have the possibilities and desire to engage in sexual intercourse, sex (in the everyday sense) becomes possible. But what are these possibilities and such desires based on? On the existence of morphological, physiological, and behavioral adaptations that ensure fertilization in representatives of two different sexes. On the existence of dioecious populations, where individuals of different sexes interact. On the fact that sexual recombination has become and remains the main means of forming new genotypes in the overwhelming majority of of highly organized species, including humans. Is such a situation the only possibility? Of course not; let us recall clonal populations that get by without sex; populations of hermaphrodites that get by without different sexes; the possibility of human multiplication through natural cloning — the formation of identical twins as a result of mitotic divisions of a fertilized egg cell.
We have mentioned several types of multiplication. For further discussion, we need to propose their typology. The typology we propose is shown in Table IV-19.1. It was developed (Shabanov, 2013, 2015) for discussing the features of hemiclonal population systems of the hybridogenetic complex of green frogs, and this explains the attention to Type IV of multiplication. Undoubtedly, the most ancient type of population multiplication is Ia, clonal with horizontal transfer of genetic information; this type is characteristic of prokaryotes. Probably, in most cases, representatives of Type Ib (clonal without horizontal transfer) are organisms that once had sexual recombination and then lost it. To which type the first organisms with sexual reproduction belonged is a question that can be discussed; perhaps it was a type not shown in Table IV-19.1 (monomorphic organisms that produced uniform gametes). In any case, it is clear that the origin of sexual recombination could not have been connected with Type IIIb; for significant ecological differences between sexes to arise, separate sexes had to already exist, that is, Type IIIb populations had to have developed from Type IIIa populations.
None (due to sterility of interspecific hybrids, which is caused by failures in recognition of homologous chromosomes by each other during meiosis)
Exist; in typical cases, species are communities with a certain gene pool that are genetically isolated from other species
DoubtfulFirst, let us consider what the marvel of sexual reproduction entails. Through our mortal bodies flows a potentially immortal succession of germ cells—the embryonic line. Both the body and the “I” of each of us form a soma, a dead‑end lateral branch of the germ line (Fig. IV‑19.1). The success of the soma is the transmission of the germ line onward, a step toward its immortality… In the case of sexual recombination, each time a germ passes through the soma it is restored: each fragment of the parental nuclear genetic information is lost with probability 1/2 and transmitted to the next generation with probability 1/2. This is especially striking compared with cytoplasmic inheritance: mitochondrial DNA is inherited clonally; inheritance of the Y chromosome in humans occurs without recombination, but with probability 1/2.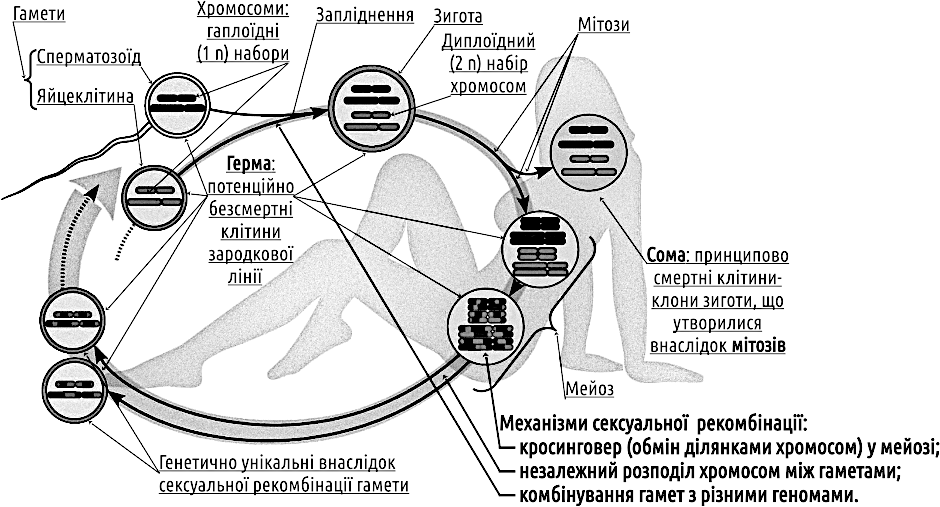
Fig. IV‑19.1. Typical life cycle with sexual recombination, fertilization, meiosis, and diploid and haploid phases
We noted that the existence of sexual recombination is paradoxical. To demonstrate this, we will split the explanation of observed natural phenomena into two stages: first, an explanation of how the discussed phenomenon arose, and second, an explanation of why it continues to exist, to be maintained. Let us examine the existence of sex (sexual recombination) and dioecy from this perspective. In doing so we encounter a series of unresolved circumstances. For convenience we will label them as paradoxes associated with eminent figures in the history of evolutionary biology.
The “Dawkins paradox” (as we shall call it) concerns the origin of sex. The processes shown in Fig. IV‑19.1 are the result of coordinated work of a suite of molecular‑genetic mechanisms. Their complex, mutually coordinated complex is likely to have arisen only once in the history of life. Complex adaptations of biosystems are the product of selection. Thus we see the outcome of prolonged selection that shaped a genetic complex that enables sex. How does selection act on genes (and on biosystems of other levels)? By increasing their fitness: their contribution to subsequent generations. Despite various ways of measuring fitness, all rest on the same premise: traits that increase the chances of preservation and spread in the future are retained. From this viewpoint it is paradoxical that the transition from asexual to sexual reproduction is associated with a two‑fold reduction in the fitness of all genes that enable it!
We associate this paradox with the name of the person who made the greatest contribution to the idea that selection operates at the genetic level (see section I‑18). At the organismal level (e.g., in hermaphrodites) and at the population level the shift to sexual recombination need not entail a halving of fitness, but at the gene level this effect is evident! Even without sharing Dawkins’s view of the omnipotence of gene‑level selection, it is hard to understand how organismal or group selection could have produced such intricate genetic adaptations. For Dawkins, who insists that all complex adaptations are tied to gene‑level selection, this paradox is especially painful.
“When reproduction is asexual, the stakes are equal and shared, because all replicators have the same 100 % chance of ending up in each offspring produced by their joint effort. In sexual reproduction the corresponding chance for each replicator is only half of the full amount, but the ritualized courtesy of meiosis, Hamilton’s ‘gavag’, largely guarantees each allele equal chances to reap the benefits of the collective reproductive enterprise. This, of course, raises the question: why is the chromosomal ‘gavag’ so courteous? This is an extremely important question, which I evade by a simple appeal to cowardice. It is one of a series of questions about the evolution of genetic systems that have been more or less fruitlessly wrestled with by the best minds after me (Williams 1975, 1980; Maynard Smith 1978a), a series of questions that prompted Williams to note that ‘a kind of crisis is maturing in evolutionary biology.’ I do not understand why meiosis occurs as it does, but given that it does, much follows. In particular, an organized honest process of meiosis helps explain the concordance and harmony that unites the parts of an individual organism. If, at the group level, a potential reproductive privilege were granted with the same scrupulous honesty in an equally well‑disciplined ‘gavag of organisms,’ group selection might become a more plausible theory of evolution.”
Richard Dawkins. The Extended Phenotype: The Long Reach of the Gene, 1999.
In our view, an explanation of the origin of sex is possible only if we predict a major role for group selection in this process. As Maynard Smith notes, the idea that the emergence of sex could be a consequence of group selection belongs to Ronald Fisher—the founder of the genetic theory of evolution. Dawkins does not wish to acknowledge a key role for group selection; although in The Extended Phenotype he concedes the principled possibility of group selection, he insists that no complex adaptations could arise. Consequently, Dawkins chooses to retreat into his own ignorance as the best solution. Why speak of group selection? Because we must construct arguments such as: “although members of the phylogenetic line that transitioned to sex reproduced less efficiently than members of related clonal lines, the probability of extinction of that line due to [choose appropriate: unpredictable environmental change; parasite evolution; genome degradation, etc.] is lower.” Thus we consider not the success of an individual organism or the preferential spread of certain genes, but the success or failure of particular groups of organisms.
Most likely, the emergence of sex was a unique event in the history of life. It is probable that the appearance of meiosis, fertilization, the cell nucleus, and the eukaryote‑typical genome organization constitute a suite of innovations that resulted from the origin of eukaryotes via symbiogenesis between archaea and alpha‑proteobacteria (which became mitochondria). According to the hypothesis of William Martin and Eugene Koonin (2006), the changes that occurred during eukaryogenesis were linked to the need to protect the host‑cell genome from mobile genetic elements (retrotransposons) introduced by the symbiont; these mobile elements became group II introns. Under this hypothesis, rapid evolutionary reorganizations driven by the necessity to safeguard the host genome led to a remodeling of the prokaryotic genetic recombination system and the emergence of meiosis. In such scenarios, what matters is not how the fitness of “sex genes” changes (given the virtual nature of such concepts), but whether organisms that combine the advantages of archaeal and bacterial metabolism persist or disappear due to destructive genomic processes. This hypothesis ties the origin of sex to group selection rather than gene‑level selection.
Investigating unique events that occurred under poorly reconstructible conditions is a difficult task. Yet even if we cannot explain how sex originated, we will be forced to acknowledge that it exists. We see numerous examples of the loss of sex during evolution. If some groups abandon sex, why do not all? The stability of sex is easier to study than its origin, because the maintenance of sexual stability is occurring right now in a huge number of modern groups.
The “Jenkin paradox,” as we understand it, concerns the stability of sexual recombination despite its continual disruption of successful hereditary trait combinations. It should be noted that the so‑called “Jenkin’s nightmare” (the name Charles Darwin used in a private letter for the argument of Fleming Jenkin, a Scottish engineering professor known as the inventor of the cable car and underwater telegraph cable) is usually misunderstood. To examine it in detail, we quote the primary source.
“Imagine a white man who suffers a shipwreck on an island inhabited by Negroes … Imagine that he possesses the physical strength, energy and abilities of the dominant white race … grant him all the advantages we can imagine for a white over a native … yet none of these assumptions leads to the conclusion that, after a limited or unlimited number of generations, the island’s inhabitants will be white. Our hero, the shipwreck survivor, will probably become king; he will kill many Negroes in the struggle for existence; he will have many wives and children, while many of his subjects will live and die as bachelors … and still he will not have a single generation to make his subjects’ descendants white. One could say that white colour is not the cause of advantage. That is true, but it can be used only to show how qualities belonging to one individual in large numbers must gradually be erased. In the first generation several dozen intelligent young mulattoes will appear, who by average intelligence will far outstrip the Negroes. One might expect that after a few generations the throne would be occupied by a more or less yellow king; but can anyone believe that the whole island would gradually be settled by whites, or even by a yellow population, or that the islanders would acquire the energy, courage, ingenuity, patience, self‑control, endurance that allowed our hero to kill so many of his ancestors and sire so many children; those qualities that, in fact, would be selected away by the struggle for existence, if it could select at all?”
Fleming Jackson. Review of “The Origin of Species.” North‑British Review, 1867.
We see that Jenkin’s argument (according to the views of that time) is openly racist; debating it with deeply compromised notions of racial capabilities is pointless. It is clear why Jenkin’s argument is avoided in direct citation and discussed only by analogy, for example: “Suppose a red poppy appears in a field of white poppies. It is more noticeable to pollinators and therefore gains an advantage over the white poppies. Sadly, its offspring will be only pink, in the second generation pale‑pink, and soon the new trait will disappear. Only Mendel’s discovery, that poppy colour is determined by a discretely inherited gene, helped solve this problem.” Yet using such analogies substitutes the argument! Poppy colour is a trait that may depend on a single gene, whereas human skin colour, let alone “energy, courage, ingenuity, patience, self‑control, endurance,” are not monogenic traits! The traits Jenkin mentions depend on a very complex interaction of many hereditary features (and on development under favourable conditions). Even if Jenkin’s argument is explained without specifying which traits are meant, it replaces traits that depend on one or a few genes with traits that are derived from almost the entire genotype.
“In essence, Mendelism undermined Fleming Jenkin’s claim that natural selection is ineffective because new traits would arise through crossing with unchanged individuals. If inheritance is partial rather than blended, a favourable new trait would remain intact and its frequency would gradually increase in the population … Where there was considerable genetic variability, because several genes or alleles affected the same trait, variation was maintained in the population only because inheritance kept gene‑variants untouched from generation to generation.”
Peter Bowler, “The Mendelian Revolution,” 1989.
The essence of Jenkin’s argument is that under sexual reproduction traits that secured the success of a particular individual can “dissipate” in subsequent generations if they depend on the interaction of many genes. A successful individual is more likely to leave successful offspring if it reproduces its genotype, which has been vetted by environmental interaction, rather than randomly combining a half of its genes with a half of another, even if that other is also highly successful. Does this phenomenon not seem odd to you? Consider it from another angle.
We view evolution as the refinement of programs that influence organismal development. Genotypes that promote phenotypes better suited to environmental interaction are more likely to be retained and reproduced. “Winners of natural selection” are those who survived and reproduced; their success was likely shaped not only by chance but also by the perfection and coherence of their genotype. And what do these “winners” do? They destroy their genotypes by mixing their fragments with fragments of other “winners of natural selection”! Imagine a mountaineer who has climbed Mount Jomolungma; he has overcome a difficult route, passing the bodies of the defeated, for it is reported that the slopes of Jomolungma are strewn with the corpses of failed climbers. Our “winner” at the summit—i.e., according to our analogy, survived and proceeds to reproduce—carries a certain set of equipment (genome); presumably, with this set he overcame all obstacles, proving its reliability and complementarity. The act of the climber beginning to exchange randomly selected parts of his equipment with someone else who has also reached the summit would seem surprising. Likewise, the characteristic re‑organization of their own genomes that contributed to their success should surprise us!
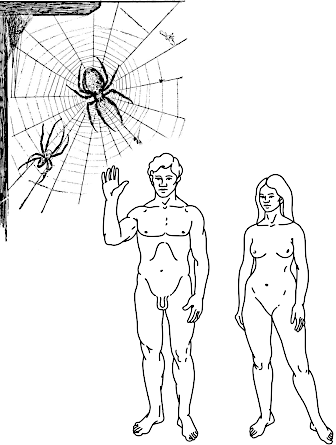
Another aspect that can be illustrated by this analogy concerns the need for another “winner” for the reproduction of a “winner of natural selection,” most often of the opposite sex. The advantages of victory will be used by the one who climbs the allegorical Jomolungma not alone… In literary terms this limitation can be denoted as the Robinson Crusoe paradox (Fig. IV‑19.2). The prototype for the character in Daniel Defoe’s novel was the British sailor Alexander Selkirk, who was stranded on a deserted island off the coast of Chile from 1704‑1709. According to the novel, Robinson Crusoe spent 24 years in solitude on the island, then rescued a native whom he named Friday. After four years of living together, Crusoe was able to leave the island. If human populations belonged to type I in the classification shown in Table IV‑19.1, Crusoe would have colonised the island with his clonal offspring in 24 years. If human populations belonged to type II, Crusoe would have had to wait for Friday, but their offspring would have been recombinants and would have enjoyed all the benefits of sexual reproduction. However, human populations belong to type III, which imposes important ecological constraints on the search for reproductive partners.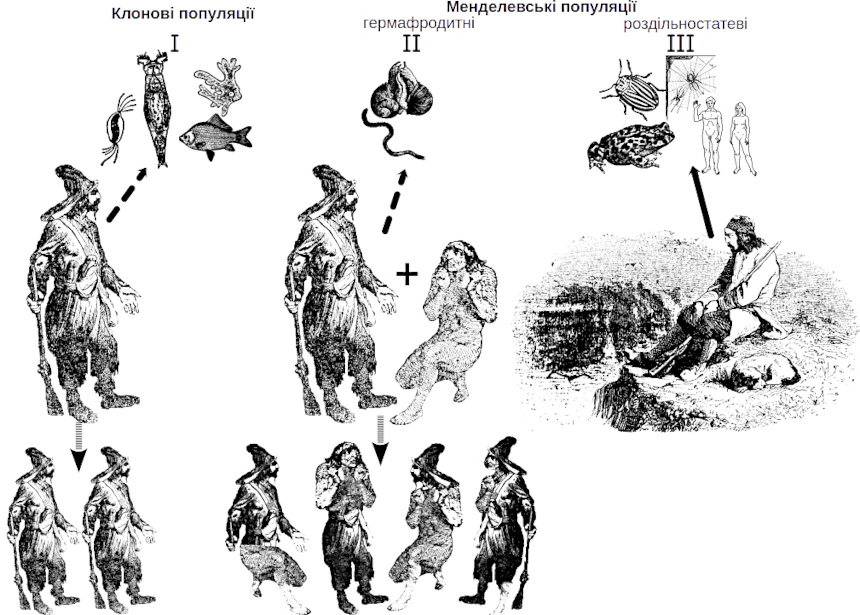
Fig. IV‑19.2. Robinson Crusoe paradox: if humans reproduced clonally, Crusoe would have colonised the island with clonal offspring; if humans were hermaphrodites, recombinant offspring of Crusoe and Friday would have appeared. But humans reproduce “the least successful” way for colonising new territories, i.e., dioeciously, and therefore Crusoe could not populate the island with his offspring.
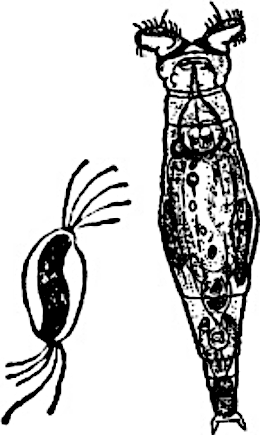
In such a case, while things are going well (or there is no suitable company), it is better to reproduce clonally, and sexual reproduction should be invoked only when problems begin to accumulate, and only if a suitable partner is available. This assumption can be supported by numerous examples. For several groups that alternate sexual and asexual reproduction (e.g., Daphnia, mayflies, etc.) a typical pattern is that under favourable conditions asexual (parthenogenetic) reproduction occurs, while under deteriorating conditions (e.g., autumnal cold) a shift to sex and the production of overwintering eggs takes place (Fig. IV‑19.3). Life cycles similar to that shown in Fig. IV‑19.3 can help understand the next paradox (which we will call the Williams paradox and discuss later in connection with problems related to the origin and maintenance of dioecy).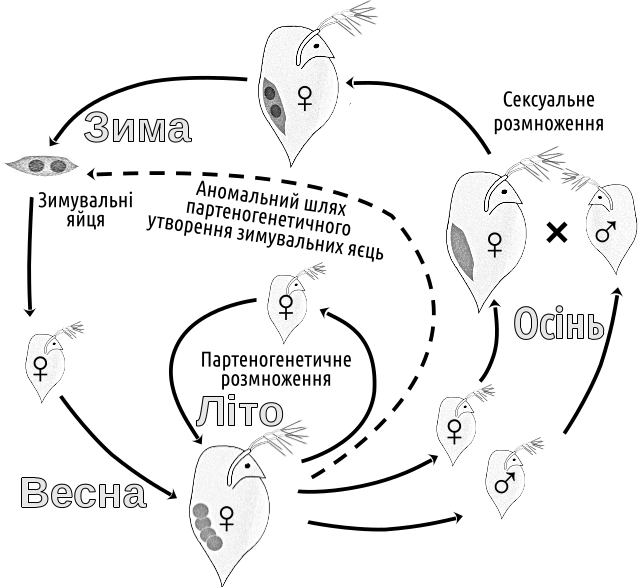
Fig. IV‑19.3. Typical annual cycle of Daphnia
We have already noted that we lack sufficient data to confidently describe the first organisms that used sex. It is entirely possible that sex was not directly linked to reproduction in them; it could have been a means of generating hybrid genotypes whose reproduction occurred at a later life‑cycle stage. Which cell nuclei fused at fertilisation? It is quite plausible that they were nuclei of unspecialised cells. The diversity of sexual processes observed today in various algae and fungi illustrates this. For example, fertilisation in which two unspecialised vegetative algal cells fuse is called conjugation, and the fusion of two vegetative cells bearing flagella (observed in some algae and some fungi) is called hologamy. After the spread of sex, the cells participating in fertilisation became specialised and turned into gametes. There is no reason to expect that these gametes were immediately divided into two types, large and small, eggs and sperm. If the evolution of sex passed through a hologamous stage, the next step could have been isogamy, a case in which two motile gametes that are morphologically identical take part in fertilisation. Anisogamy is the case in which one of the two motile gametes is larger and the other smaller. Anisogamy could have been a transition to oogamy—the most common case, in which fertilisation occurs between a relatively large, non‑flagellated egg and a small motile sperm.
The transition from isogamy to anisogamy and oogamy has a solid theoretical explanation often referred to as the PBS theory (PBS theory, after the initials of the authors G. Parker, R. Baker, and V. Smith (1972) “The origin and evolution of gamete dimorphism and the male‑female phenomenon”). According to the PBS approach, in a population where the norm is the production of gametes that carry roughly half of the nutrient reserves needed for zygote development, individuals that produce a somewhat larger number of slightly smaller gametes may gain an advantage.{"translated_text": "Under such conditions, increasing the number of potential offspring may have a stronger effect on the fitness of such individuals than reducing offspring survival. Under conditions where small gametes are dispersed, individuals producing enlarged gametes containing sufficient nutrient reserves may gain an advantage, regardless of how much the other gamete provides.\nHowever, it should be emphasized that the transition from isogamy to oogamy may be associated with the transition from the absence of sex (and gamete production not divided into two types) to gonochorism (production of a specific gamete type) or to hermaphroditism (production of two types of gametes either simultaneously or sequentially). We cannot be certain whether hermaphroditism is primary (hermaphrodites first arose, followed by separate males and females) or secondary (using the designations used in Table IV-19.1, it can be suggested that type II populations could potentially arise from type IIIa populations). Regardless, the mysterious transition to gonochorism was important in the history of a huge number of groups. Let us begin with the paradox associated with the appearance of two sexes.\nMaynard Smith's paradox or the paradox of the double cost of males («two-fold cost of males») or the paradox of the double cost of sex («two-fold cost of sex») consists in the fact that the transition to gonochorism leads to a twofold reduction in the reproductive potential of a population. Let us compare the number of offspring produced by two individuals in the case of sexual (gonochoristic organisms, type III according to Table IV-19.1) and asexual (for example, clonal parthenogenetic, type Ib) reproduction. Let us denote the brood size produced by a gonochoristic or parthenogenetic female as n. In the case of sexual reproduction, the number of offspring in the parental pair will equal n, in the case of abstaining from sex — 2×n. In the next generation, sexual organisms can obtain n×(n/2) offspring (since the number of offspring depends on the number of females, which constitute half of the individuals in each generation), and asexual organisms — 4×n²! This is a very significant difference! Discussing the origin of sex, we agreed with Fisher's assumption that it could have been supported by group selection. According to the «two-fold cost of males» paradox, group selection should powerfully «play» against sex and gonochorism!\n«In effect, a parthenogenetic female has a double selective advantage because she does not expend energy on producing sons. Of course, this argument is valid only under ceteris paribus conditions, that is, if there are no genetic differences affecting fitness between sexual and asexual females.»\nJohn Maynard Smith. «The Evolution of Sex», 1978\nBy the way, which of the designations used, «two-fold cost of males» or «two-fold cost of sex», is better? The latter is more widespread, but the former is more precise. For example, hermaphroditic populations, type II populations according to the classification shown in Table IV-19.1, do not pay this double cost: they have sex, but no males.\nA logical question may arise — can the «cost of males» be less than double, not double? Let us imagine a population where 9/10 are females and only 1/10 are males. In such a gonochoristic population, the number of offspring will reach 90% of that in a clonal one. In this case, the «cost of males» will equal not 1/0.5=2, but only 1/0.9=~1.11 — much less. If the «cost of males» can be reduced so easily, why do we so often observe sex ratios close to 1:1? This question has a universally accepted explanation, which Fisher provided in 1930.\nFisher's equilibrium (or Fisher's principle) — the principle whose cause lies in the fact that in gonochoristic populations each individual has one mother and one father. As a result, in a population where 9/10 are females and only 1/10 are males, the male will have on average 10 times more offspring than the female! In this case, selection will favor those individuals in whose offspring the proportion of males is higher than 1/10. The proportion of males will increase, but not indefinitely. When will its increase stop? When the average number of offspring in a male equals that in a female. In fact, the situation is somewhat more complex. In cases where producing a male offspring and producing a female offspring require different energy expenditures for the parents, selection will equalize not the expected number of further offspring per one offspring, but such a number of offspring per equal energy expenditure. However, in most cases the expenditures on offspring of different sexes are approximately equal, and this is the reason why the sex ratio in most gonochoristic populations approaches equality. Another interesting consequence is that hermaphroditic individuals with cross-fertilization must expend equal amounts of energy on producing female and male gametes!\n«Let us first consider a hermaphroditic animal species with external fertilization (for example, ascidians), or, if you prefer, a self-incompatible hermaphroditic or monoecious plant. Such species, as a rule, allocate the resources designated for reproduction equally between eggs and sperm or between seeds and pollen. The reason for such equal distribution is essentially the same that allowed Fisher (1930) to predict the 1:1 sex ratio in gonochoristic species <...> Hence, a parthenogenetic individual, who does not need to allocate any resources to performing male functions, could produce twice as many eggs or seeds.»\nJohn Maynard Smith. «The Evolution of Sex», 1978\nBy the way, Fisher's equilibrium is an example of selection at the gene level «winning» over group selection in many cases. Although populations with significantly skewed sex ratios could be much more successful, we do not observe them around us (in any case — in the typical case). Selection for increasing the reproductive success of each individual (or, from another perspective, — for more efficient reproduction of its genes affecting the sex ratio in offspring) supports those individuals that produce relatively equal proportions of offspring of both sexes.\nThus, populations that transitioned to gonochorism had to overcome a high threshold: to halve the efficiency of reproduction. Moreover: if at any stage gonochoristic populations abandon their paradoxical method of reproduction, they should obtain corresponding advantages! This phenomenon was also examined in detail by John Maynard Smith, but he referred to George Williams. Williams considered situations where two methods of reproduction are realized in one population, as in Fig. IV-19.2. Of course, in the Daphnia shown in the figure, the coexistence of sexual and asexual reproduction is the norm. On the other hand, it is possible to consider another case, where asexual reproduction appears in a sexual population (for example, as a result of the appearance of a parthenogenetic form that abandoned meiosis). The advantages of sexual reproduction are «slow», they will manifest themselves in an evolutionarily significant perspective. The advantages of asexual reproduction must be «rapid», manifesting within a short period.\n«The strongest argument for a rapid advantage of sex was advanced by Williams (1975) as the «balance» argument. If parthenogenetic and sexual modes of reproduction coexist in one population, then there must be some immediate advantage of sexual reproduction that would balance its double disadvantage. Indeed, one could expect that natural selection would alter the relative frequencies of sexual and asexual reproduction until the selective advantages are balanced. Let us formulate the same argument differently. Group selection can support sex by eliminating parthenogenetic variants if the latter arise sufficiently rarely, but cannot maintain a balance between these two processes if group extinction is not extremely widespread.»\nJohn Maynard Smith. «The Evolution of Sex», 1978\nWilliams's paradox or Williams's balance argument consists in the fact that in the case of combining sexual and asexual reproduction in one population (as in Fig. IV-19.3) asexual reproduction will have a «rapid» (such as will manifest within a few generations) advantage. For sexual reproduction not to disappear, it must be constantly supported by some other «rapid» advantage.\nIs the appearance of a parthenogenetic female in a gonochoristic population something realistic? Yes; such phenomena have been recorded in a large number of groups (with the exception of mammals, in which parthenogenesis is probably blocked by epigenetic mechanisms that provide genomic imprinting). Let us give an example of Burmese pythons (Python bivittatus), which were previously considered a subspecies of the tiger python (obsolete name — P. molurus bivittatus). These are large and beautiful snakes that are not difficult to keep and breed in captivity. Thanks to this, extensive data have been accumulated on the reproductive biology of this species. It is known that pythons sometimes hatch from unfertilized eggs laid by a female that has not mated with males; recently it has been proven that such individuals are females, and that they are genetically identical to their mother. How heritable the ability for parthenogenesis is in such females is unknown, but it can be assumed that, as with many other similar traits, there must be a significant heritable component in the propensity for parthenogenesis."}