
IV. Amphibia sensu lato-11. Batrachofauna and Herpetofauna of Kharkiv Oblast: An Overview
An excerpt from a field practice manual for vertebrate zoology (with minor modifications). The section concerning reptiles was written primarily by O.I. Zinenko, and the section concerning amphibians primarily by D.A. Shabanov. Atemasova T.A., Vlashchenko A.S., Goncharov G.L., Zinenko ...
An excerpt from a field practice manual for vertebrate zoology (with minor modifications). The section concerning reptiles was written primarily by O.I. Zinenko, and the section concerning amphibians primarily by D.A. Shabanov.
Atemasova T.A., Vlashchenko A.S., Goncharov G.L., Zinenko O.I., Korshunov O.V., Tokarsky V.A., Shabanov D.A., Shandykov G.A. Field Practice in Vertebrate Zoology. – Kharkiv: V.N. Karazin Kharkiv National University, 2019. – 196 p.
Composition and characteristics of the herpetofauna of Kharkiv Oblast
In the scientific literature, the concept of "herpetofauna" is used in two senses: in the narrow sense it denotes only the fauna of reptiles (Greek herpeton — reptile), and in the broad sense — the fauna of all herpiles (i.e., amphibians and reptiles). Here we use the term "herpetofauna" in the second of these senses, encompassing both the batrachofauna and the herpetofauna proper.
Amphibians and reptiles constitute a notable component of the terrestrial vertebrate fauna of Kharkiv Oblast (as well as Ukraine and Eurasia as a whole), yet their diversity is limited to only a few groups. This is associated with the concentration of the greatest species richness in tropical regions, as well as with the peculiarities of the evolutionary history of this group, whose heyday lies in the distant past: in amphibians, parareptiles, and theromorphs it fell during the Carboniferous–Triassic, and in typical diapsid reptiles during the Triassic–Cretaceous. In those remote times, the aforementioned groups were represented by several subclasses and a large number of orders. Later, when other groups assumed leading positions, only a few orders remained of the herpiles, and representatives of each have a rather distinctive appearance. Some of these orders are currently ecologically successful as well. Thus, the number of species in the order Squamata worldwide as of April 2019 was 10,503 species, and the number of species in the order Anura was 7,058 (considerably more than the number of species in the class Mammalia). It should be noted that new research provides grounds for an increase in the number of species within these groups.
According to the traditional system, herpiles are divided into two classes: Amphibia and Reptilia. Amphibians are anamniotes, possessing naked skin that plays a substantial role in respiration; they lack a thoracic cage and ventilate the lungs by means of a buccal pump. Reptiles are amniotes; their skin is covered with cornified scales, and they are characterised by the presence of a thoracic cage.
Below is a systematic checklist of amphibian and reptile species inhabiting Kharkiv Oblast, based on modern systematics (Table 3.1). Since the new names do not correspond to most of the literature, the traditional (conventional) taxon names are given in parentheses after the "=" sign.
Checklist of amphibians and reptiles of the fauna of Kharkiv Oblast
|
Ukrainian name
|
Latin name
|
Russian name
|
|
Class Amphibia (Amphibians)
|
Amphibia
|
Class Amphibia (Amphibians)
|
|
Riad Khvostati
|
Caudata
|
Otriad Khvostatye
|
|
Rodyna Salamandry
|
Salamandridae Goldfuss, 1820
|
Semeistvo Salamandry
|
|
Genus Lissotriton Bell, 1839
|
Lissotriton Bell, 1839
|
Genus Smooth Newts
|
|
Smooth Newt
|
Lissotriton vulgaris (Linnaeus, 1758)
|
Smooth Newt
|
|
Rid Tryton
|
Triturus Rafinesque, 1815
|
Rod Tryton
|
|
Great Crested Newt
|
Triturus cristatus (Laurenti, 1768)
|
Great Crested Newt
|
|
Riad Bezkhvosti
|
Anura
|
Otriad Beskhvostye
|
|
Rodyna Kumky
|
Bombinatoridae Gray, 1825
|
Semeistvo Zherlianky
|
|
Rid Kumky
|
Bombina Oken, 1816
|
Rod Zherlianky
|
|
Fire-bellied Toad
|
Bombina bombina (Linnaeus, 1761)
|
European Fire-bellied Toad
|
|
Rodyna Chasnychnytsi
|
Pelobatidae Bonaparte, 1850
|
Semeistvo Chesnochnytsy
|
|
Rid Chasnychnytsi
|
Pelobates Wagler, 1830
|
Rod Chesnochnytsy
|
|
Chasnychnytsia Pallasa
|
Pelobates vespertinus (Pallas, 1771)
|
Chesnochnytsa Pallasa
|
|
Rodyna Kvakshi
|
Hylidae Rafinesque, 1815
|
Semeistvo Kvakshy
|
|
Rid Kvakshi
|
Hyla Laurenti, 1768
|
Rod Kvakshy
|
|
Eastern Tree Frog
|
Hyla orientalis Bedriaga, 1890
|
Eastern Tree Frog
|
|
Rodyna Ropukhy
|
Bufonidae Gray, 1825
|
Semeistvo Zhaby
|
|
Rid Ropukhy
|
Bufo Laurenti, 1768
|
Rod Zhaby
|
|
Common Toad
|
Bufo bufo (Linnaeus, 1758)
|
Common Toad
|
|
Genus Bufotes Rafinesque, 1815
|
Bufotes Rafinesque, 1815
|
Genus Green Toads
|
|
Green Toad
|
Bufotes viridis (Laurenti, 1768)
|
Green Toad
|
|
Rodyna Zhaby
|
Ranidae Rafinesque, 1814
|
Semeistvo Liahushky
|
|
Genus Rana Linnaeus, 1758
|
Rana Linnaeus, 1758
|
Genus Brown Frogs
|
|
Moor Frog
|
Rana arvalis Nilsson, 1842
|
Moor Frog
|
|
Genus Pelophylax Fitzinger, 1843
|
Pelophylax Fitzinger, 1843
|
Genus Green Frogs
|
|
Pool Frog
|
Pelophylax lessonae (Camerano 1882)
|
Pool Frog
|
|
Marsh Frog
|
Pelophylax ridibundus (Pallas, 1771)
|
Marsh Frog
|
|
Edible Frog
|
Pelophylax esculentus (Linnaeus, 1758)
|
Edible Frog
|
|
Class Reptilia
|
Reptilia
|
Class Reptilia (Reptiles)
|
|
Riad Cherepakhy
|
Testudines
|
Otriad Cherepakhy
|
|
Family Emydidae, 1815
|
Emydidae, 1815
|
Family Freshwater Turtles
|
|
Genus Emys Dumeril, 1806
|
Emys Dumeril, 1806
|
Genus Pond Turtles
|
|
European Pond Turtle
|
Emys orbicularis (Linnaeus, 1758)
|
European Pond Turtle
|
|
Riad Luskati
|
Squamata
|
Otriad Cheshuichatye
|
|
Pidriad Yashchirky
|
Sauria (Lacertilia)
|
Podotriad Yashcherytsy
|
|
Family Lacertidae Bonaparte, 1831
|
Lacertidae Bonaparte, 1831
|
Family True Lizards
|
|
Genus Lacerta Linnaeus, 1758
|
Lacerta Linnaeus, 1758
|
Genus Green Lizards
|
|
Sand Lizard
|
Lacerta agilis Linnaeus, 1758
|
Sand Lizard
|
|
Genus Zootoca Wagler, 1830
|
Zootoca Wagler, 1830
|
Genus Forest Lizards
|
|
Viviparous Lizard
|
Zootoca vivipara (Jacquin, 1787)
|
Viviparous Lizard
|
|
Rid Yashchurky
|
Eremias Wiegmann, 1834
|
Rod Yashchurky
|
|
Varied Lizard
|
Eremias arguta (Pallas, 1773)
|
Steppe Runner
|
|
Rodyna Veretilnytsi
|
Anguidae Gray, 1825
|
Semeistvo Veretenytsy
|
|
Rid Veretilnytsi
|
Anguis Linnaeus, 1758
|
Rod Veretenytsy
|
|
Slow Worm
|
Anguis colchicus (Nordmann, 1840) (=A. fragilis Linnaeus, 1758)
|
Slow Worm
|
|
Pidriad Zmii
|
Serpentes
|
Podotriad Zmey
|
|
Rodyna Vuzhopodibni
|
Colubridae Oppel, 1811
|
Semeistvo Uzheobraznye
|
|
Genus Natrix Laurenti, 1768
|
Natrix Laurenti, 1768
|
Genus True Grass Snakes
|
|
Grass Snake
|
Natrix natrix (Linnaeus, 1758)
|
Grass Snake
|
|
Dice Snake
|
Natrix tessellata (Laurenti, 1768)
|
Dice Snake
|
|
Rid Midianky
|
Coronella Laurenti, 1768
|
Rod Medianky
|
|
Smooth Snake
|
Coronella austriaca Laurenti, 1768
|
Smooth Snake
|
|
Rodyna Hadiuky
|
Viperidae Laurenti, 1768
|
Semeistvo Hadiuky
|
|
Rid Hadiuky
|
Vipera Laurenti, 1768
|
Rod Hadiuky
|
|
Nikolsky's Viper (= Common Viper)
|
Vipera nikolskii Vedmederja, Grubant, Rudaeva, 1984 (= Pelias nikolskii (Vedmederja, Grubant, Rudaeva, 1984); Vipera berus Linnaeus, 1758)
|
Nikolsky's Viper (= Common Viper)
|
|
Steppe Viper
|
Vipera renardi (Christoph, 1861)
|
Steppe Viper
|
Several more amphibian species have been mentioned in the literature for Kharkiv Oblast, but the question of their presence in the region remains open. Thus, at present no reliable record of the Common Frog Rana temporaria Linnaeus, 1758 is known, although it has been noted in a number of authoritative sources both in the past (O.M. Nikolsky) and in the present (S.L. Kuzmin). It is unclear whether this represents the disappearance of this species from Kharkiv Oblast or misidentification of the Moor Frog.
Contradictory data concern the European Green Lizard Lacerta viridis (Laurenti, 1768). It was recorded from the vicinity of Krasnohrad; the nearest isolated population exists near Poltava (Sukhov, 1928). A single specimen of the Green Lizard was brought together with Sand Lizards and Green Frogs to the Museum of Nature of V.N. Karazin Kharkiv National University by university students in the 1970s from Zachepylivka district, but special searches by Museum of Nature staff O.M. Rudyk and V.I. Vedmederya in that district yielded no results. It should be noted that records of the Viviparous Lizard, whose presence in the oblast has now been confirmed, were considered doubtful for almost 100 years.
In historical times, the ranges of three ratsnake species approached or even entered the territory of Kharkiv Oblast: the Steppe Ratsnake Elaphe dione (Pallas, 1773), records of which are known from Luhansk and Donetsk oblasts (possibly still living in the south of Kharkiv Oblast in Izium district); the Pallas's Ratsnake, or Sarmatian Ratsnake, Elaphe sauromates Pallas, 1814; and the Caspian Whipsnake Dolichophis caspius (Gmelin, 1789). Thus, a specimen of Pallas's Ratsnake caught in 1905 between Lozova and Blyzniuky in Kharkiv Governorate is preserved in the Zoological Institute of the Russian Academy of Sciences (St. Petersburg). Local residents claim that some snakes matching the description of Pallas's Ratsnake or the Caspian Whipsnake lived near the village of Varvarivka in Blyzniuky district but disappeared in the 1950s following the onset of mass use of pesticides (according to V.I. Vedmederya). In the 1980s, a "Caspian Whipsnake" was observed on a railway embankment near the Samara River in Blyzniuky district of Kharkiv Oblast. Any record of a ratsnake in Kharkiv Oblast is of considerable interest both for faunistics and for nature conservation, as all ratsnake species are listed in the Red Data Book of Ukraine.
When identifying ratsnakes, the following characteristics should be used as guides. From other snakes of Kharkiv Oblast, the Steppe Ratsnake and Pallas's Ratsnake are distinguished by the large number of dorsal scale rows around mid-body (23–28) and ventral scutes (171–234), and by the presence of paired apical pits on the body scales. Although there are distinct differences in pattern and pholidosis between these two ratsnakes, their identification requires considerable experience. The Caspian Whipsnake typically has 19 scale rows around mid-body and bears apical pits on the scales. From the Smooth Snake it differs by its considerably larger size (up to 2 m, the largest snake in Europe), uniform dorsal colouration without pattern in adult individuals, and a large number of ventral scutes (189–192). Any identification is best confirmed by a specialist, and in the event of a find the specimen or high-quality photographs should be submitted. This last piece of advice applies, however, to any other animal that proves difficult to identify.
Main research challenges in the study of the herpetofauna of Kharkiv Oblast
The study of the herpetofauna of Kharkiv Oblast is of interest from many perspectives. Within our region or close to it pass the range boundaries of two spadefoot toad species and of subspecies of the Sand Lizard. Two forms of the Moor Frog have been reported from the oblast. Unique in composition, populations of green frogs occur here (discussed in greater detail below). Nikolsky's Viper, the type series of which was collected in the vicinity of Kharkiv, is a species whose taxonomic status continues to be debated. In Kharkiv Oblast or nearby pass the range limits of the Common Toad (occurring in Dnipropetrovsk Oblast in the floodplain of the Oril River, in Donetsk Oblast near Sviatohirsk, and in Luhansk Oblast near Kreminna), the Pool Frog, the Slow Worm, the Varied Lizard, the Viviparous Lizard, the Dice Snake, and the Steppe Viper, which provides opportunities to study the peculiarities of species existence at the range margin; effects related to isolation and population fragmentation; and the factors determining species distribution.
Each animal species is characterised by a unique population structure that reflects the nature of interactions among individuals and of the population as a whole with its environment. Differences between populations reflect the evolutionary history of the species and its strategy of adaptation to the environment. This makes the comparison of populations (and of individuals within them) particularly interesting, both across different regions and within a single one. Due to their relatively limited mobility, herpiles form rather small, well-structured populations. Their study is a fascinating problem in herpetology, ecology, and evolutionary theory.
The destruction of the natural state of the environment and global climatic change affect the abundance of herpiles more than that of any other group of vertebrates. For many amphibian species, catastrophic reductions in abundance and the extinction of populations and species as a whole have been recorded. The foregoing makes constant monitoring of the abundance and population status of reptiles and amphibians of Kharkiv Oblast a necessity.
Recommendations for field research on amphibians and reptiles
Although amphibians and reptiles hold a notable position in terms of abundance among vertebrates, locating them in nature and observing their lives is no easy task. Small size, a short period of summer activity, a secretive mode of life, the absence of acoustic signals in most anurans outside the breeding season, and the complete "silence" of reptiles and caudates make them rather difficult subjects for observation. However, with a little experience it is possible to find almost all of them within just a few days of summer fieldwork.
In general, success in locating amphibians and reptiles depends on several factors.
Seasonal activity. The figures show phenological charts for amphibians and reptiles of Kharkiv Oblast. Unlike mammals and birds, amphibians and reptiles are capable of substantially curtailing their activity in winter or during the hottest period of the year, entering winter dormancy — hibernation — or summer dormancy — aestivation. In temperate latitudes there is naturally no chance of encountering them in winter. At other times their activity, and hence the probability of encountering them on an excursion, depends on their physiological needs. The most favourable months for searching for amphibians and reptiles are the spring months — April and May — the time of mate-searching, reproduction, and replenishment of nutritional reserves following prolonged hibernation or the breeding season.
Phenology of amphibians of Kharkiv Oblast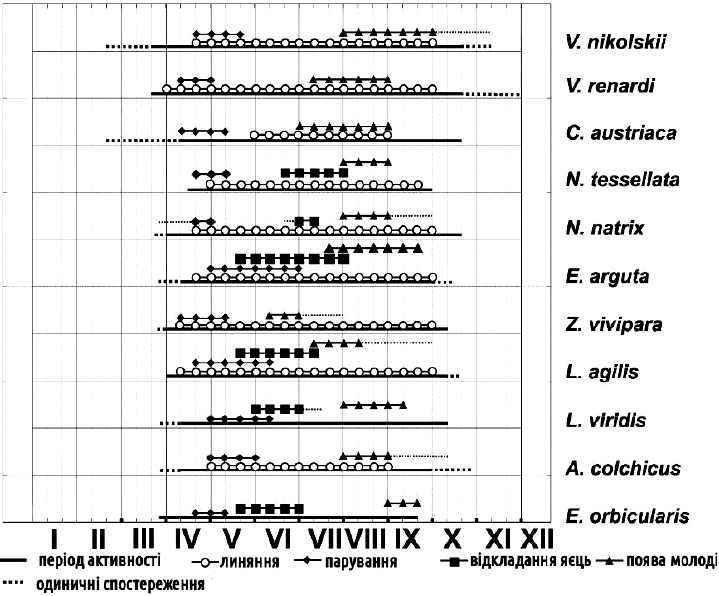
Phenology of reptiles of Kharkiv Oblast
Many amphibians are characterised by a very short spawning period (1–2 weeks), the timing of the onset of which depends on the course of spring. The typical spawning sequence of anurans in Kharkiv Oblast is as follows: Moor Frog (late March — April) — Common Toad (late March — April) — Spadefoot Toad (late March — April) — Tree Frog (April–May) — Fire-bellied Toad (April–May) — Green Frogs (May–June) — Green Toad (May–June), with the specific dates varying considerably. The duration of the breeding period can differ: for example, the breeding of the Common Toad or Moor Frog lasts about one week, that of the Green Frogs about a month, and that of the Green Toad (under stable weather conditions) sometimes up to two months.
After the breeding period, the activity of amphibians and reptiles declines, and instead of hundreds or even thousands (amphibians at spawning sites) or dozens (reptiles) of individuals per day’s excursion, one can barely encounter tens and single individuals respectively.
The beginning of summer is characterised by the continued reproduction of some amphibians (green frogs). Also at this time, amphibian larvae at various stages of development can be encountered in water bodies. During this period, females of oviparous reptile species — lizards and snakes — which are feeding and basking, show a high level of activity (and thus a high probability of encounter). Finally, only during egg-laying in mid-June can the European Pond Turtle itself, or its tracks, be seen in any numbers.
After egg-laying and a brief recovery period in the second half of summer, the visible diversity of amphibians and reptiles declines still further. This is also associated with an increase in mean daily temperatures and a decrease in precipitation. As a rule, the activity optima of our herpiles fall at temperatures up to 30 degrees Celsius. Whereas in spring or autumn months the probability of encountering them increases towards midday, in summer they can only be seen in the morning and evening hours; many amphibians switch to nocturnal or crepuscular activity, when temperatures are lower and humidity is higher.
The middle-to-late summer period is characterised by the appearance of young-of-the-year — neonates and hatchlings of reptiles, or recently metamorphosed amphibians — which, against the background of declining adult activity, briefly become the most commonly encountered finds on excursions. For example, young-of-the-year of certain amphibian species can be very numerous on the shores of water bodies on days of mass emergence onto land.
Finally, in early autumn a second peak of amphibian and reptile encounters is observed. More suitable weather conditions, concentration near hibernation sites, or reduction in extent through desiccation of water bodies makes searching for them almost as productive as in spring.
Weather conditions. The activity of amphibians and reptiles, owing to their poikilothermy, depends greatly on weather conditions. Briefly, the influence of weather conditions on activity can be characterised by several patterns. In cool seasons the probability of encountering them increases with rising temperature. When transitioning from a sharp cold spell and overcast weather to warm and clear conditions, the activity of reptiles increases, and vice versa. Summer excursions in clear weather following rainfall can be particularly productive. The activity of amphibians is higher in damp but warm weather.
Microhabitat affinity. Besides the general patterns of herpile distribution across biotopes, microhabitat affinity plays a major role. When searching for a given species, the researcher is guided more by the characteristics of the site where the species can be encountered at a given time of year and under the prevailing weather conditions. Providing an exhaustive description of such biotopes is extremely difficult, but the general patterns are as follows: almost all reptiles are associated with ecotones (boundaries between different types of biocoenoses), shelters (shrubs, fallen trees), and sun-warmed sites. For amphibians that are exclusively tied to water bodies throughout the season (Green Frogs, Fire-bellied Toads), the type of water body is of great importance, and even closely related species may prefer different water bodies. Additional data on the microhabitat preferences of individual species are contained in the species accounts.
Nevertheless, the majority of species outside the breeding period will not be available in large numbers — obtaining a sufficient number of specimens requires too great an effort in searching for them. Therefore, in practical field zoology, great importance attaches to the choice of study timing, as well as to the use of traps, drift fences, and other devices that substantially increase the efficiency of collection.