III. Pisces-01. Fishes: Their Characteristic Features and Archaic Groups
The evolutionary success of the phylum Chordata is associated with the emergence of two extremely successful groups—fishes and tetrapods (which are descendants of fishes).
I. Vertebrates Zoology II. Archaic groups: Acraniata, Tunicata, Agnatha III. Pisces
Fishes, Their Characteristic Features and Archaic Groups
IV. Amphibia sensu lato V. Reptilia sensu lato VI. Class Aves VII. Class Mammalia
← II.03. Comparative Anatomy of Lampreys
III.02. Class Chondrichthyes - Cartilaginous Fishes →
We are beginning the study of fishes. The evolutionary success of the phylum Chordata was largely associated with the appearance of two extremely successful groups - fishes and tetrapods. If we remember that tetrapods are descendants of fishes, the success of fishes becomes especially evident. Incidentally, consistent phylogenetic systematics does not permit paraphyletic groups, that is, groups that do not include all descendants of a given ancestor. From that point of view, we should consider ourselves fishes. This is inconvenient for studying vertebrate zoology, and therefore we will use paraphyletic taxa, but we will write their names in quotation marks. Thus, we move on to study the superorder “Pisces” - “Fishes”. However, in the Ukrainian text we will mostly avoid these quotation marks, because the true name of any taxon is its Latin name.
We begin with systematics, where we will show the position of fishes within the phylum and indicate their main groups. It should immediately be emphasized that ray-finned and lobe-finned fishes are very often considered separate classes. Here we return to the older system in which they are united in the class Bony Fishes. This is due to recent finds that confirmed the substantial phylogenetic affinity of these groups.
Phylum Chordata — Chordates
Subphylum Acraniata — Acraniates
Class Cephalochordata — Cephalochordates
Subphylum Tunicata — Tunicates
Class Ascidiacea (Ascidians) — Ascidians
Class Thaliacea (Salps) — Salps
Class Appendicularia — Larvaceans
† Subphylum Conodontochordata — Conodont chordates
† Class Conodonta — Conodonts
Subphylum Vertebrata — Vertebrates
Superclass “Agnatha” — Jawless vertebrates
† “Ostracodermi” — Shield fishes (~6 classes)
Class Myxini — Hagfishes
Class Petromyzontida — Lampreys
Superclass “Pisces” — Fishes
†Class “Placodermi” — Placoderms
†Class “Acanthodii” — Acanthodians
Class Chondrichthyes — Cartilaginous fishes
Class “Osteichthyes” — Bony fishes
Subclass Actinopterygii — Ray-finned fishes
Subclass “Sarcopterygii” — Lobe-finned fishes
Superclass Tetrapoda — Tetrapods
How and why did fishes appear? To answer this question, we must ask what way of life the further progress of the first vertebrates, which we united into the provisional group of shield fishes, could have been associated with. Most of the well-known representatives of that group moved to life near the bottom (and that is precisely why they more often entered the fossil record and became known to us). However, in the water column there remained some relatively small and lightly built shield fishes that preserved the original vertebrate mode of life: they shifted from filter-feeding to predatory feeding. When the resolving power of the senses of early vertebrates became sufficient to distinguish individual food particles, some of the detected particles turned out to be living. What was a living particle supposed to do after it entered the oral cavity of a primitive vertebrate - an actively swimming animal that was improving its ability to feed as a predator?
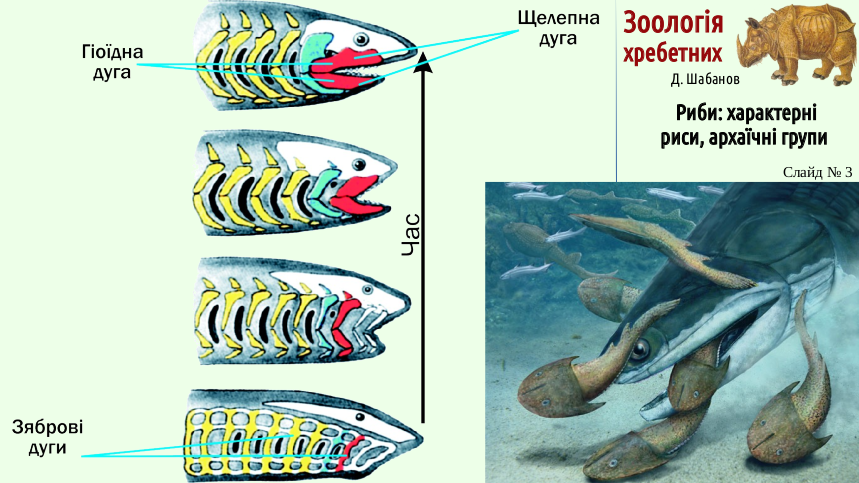
A small living organism, about to become a food particle, tried to escape from the oral cavity into the gill slit with the water current. The predator's task was to hold onto the food item and send it to the esophagus. The gill slits of such vertebrates were bounded by visceral (gill) arches. In early vertebrates these arches were segmented, consisting of separate cartilages (the continuous gill basket of lampreys is most likely a secondary acquisition associated with adaptation to breathing while attached to a substrate). Retention (and later deformation) of animals that had entered the oral cavity was added to the functions of individual elements of the visceral arches. Eventually one of the visceral arches (the jaw arch) became specialized for holding and sometimes even tearing apart prey.
The process of the origin of the visceral arch is schematically shown in the following video.
It should be emphasized that such animations, while showing some essential features of anatomical rearrangements, can mislead us about the course of evolution. There was no proto-fish in which one visceral arch simply enlarged and turned into jaws. Reality was more complex: for a long time, fishes with a more powerful and more mobile arch specialized for holding and deforming prey were the ones that mostly survived and left offspring. Between the states corresponding to adjacent frames of this animation, generations of primitive fishes passed.
The origin of jaws from visceral arches is also reflected in embryogenesis.
After effective jaws emerged, some of the animals that acquired them moved to life near the bottom and to feeding that no longer resembled filtration. This was true hunting.
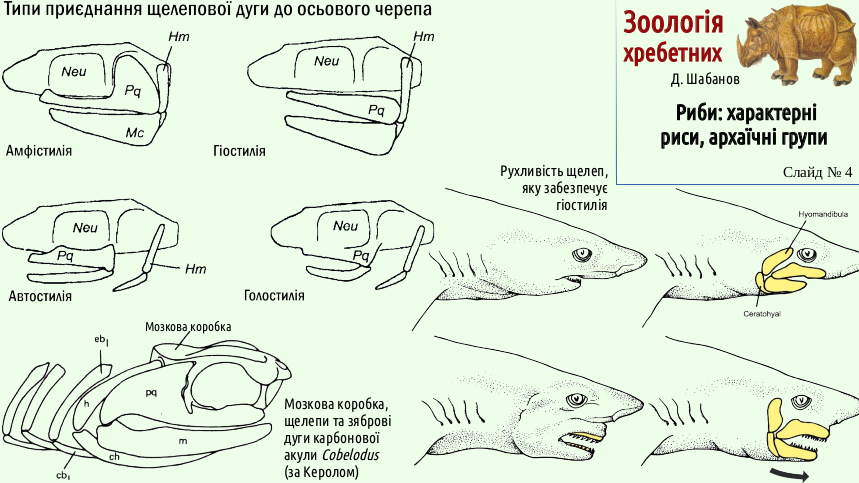
Specialization of the jaw arch for holding prey was effective when its connection with the braincase was weakened. If the victim was large enough, dealing with it required a reliable connection of the jaw arch with the axial skeleton (otherwise it might simply tear the jaws out of the unlucky predator). Strengthening the attachment of the jaw arch occurred with the help of the next visceral arch - the hyoid arch. Initially, the upper element of the jaw arch (the palatoquadrate cartilage) retained a movable connection with the braincase. At the same time, the attachment of the jaw arch was strengthened by the hyoid arch, which supported both the palatoquadrate cartilage and the Meckel's cartilage (the lower element of the jaw arch). This type of jaw-arch attachment (in which both the jaw arch and the hyoid arch are connected with the braincase) is called amphistyly.
For most fishes, hyostyly is characteristic (a variant in which only the hyoid arch is connected with the braincase, while the jaw arch “hangs” from it). It is this arrangement of the jaws that allows a shark, during a bite, to protrude its jaws beyond the normal contour of its body. For example, this makes it possible to bite a chunk out of a whale's side (this requires about the same ability as biting a chunk out of a bare wall; try it if you think it is easy).
Of course, in hyostyly the jaw arch is not rigidly fixed. If stronger closure force is needed, it is better to link the jaw arch more firmly to the axial skeleton. This is achieved in autostyly, when the jaw arch is rigidly fixed to the braincase. Finally, in holostyly the palatoquadrate cartilage fuses with the braincase into a single whole. This is the arrangement that allows chimaeras to crush the calcareous skeletons of coral polyps with their jaws.
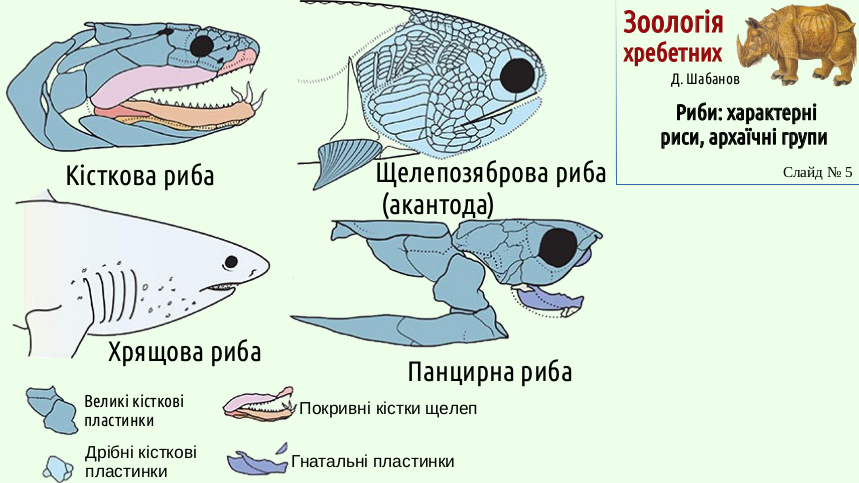
The four main groups of fishes (placoderms, acanthodians, cartilaginous fishes, and bony fishes) have different jaw structures and structures that strengthen their bite. However, these variants are probably united by common origin, and the typical representatives of different groups chose different options. In placoderms, the jaws were reinforced by strong gnathal (jaw) plates, parts of the external armor. Thanks to them, the jaws of placoderms could act like enormous shears. In cartilaginous fishes, the jaws remain cartilaginous, but teeth are located on them, and these teeth are homologous to placoid scales. In acanthodians the jaws are covered externally by small bony plates, and in bony fishes - by dermal bones.
However, predation requires not only strong jaws. What will potential prey do when a predator approaches? It will flee, first of all by moving sideways. Hunting moving prey requires a reorganization of the entire locomotor apparatus. The swimming-economy optimization characteristic of filter feeders becomes irrelevant: if a predator catches prey containing a valuable resource, it will justify all of its energy costs on high-speed swimming; if it does not catch it, no economy will matter anyway.
Take a look at this video clip. It is an emotional, powerful sight...
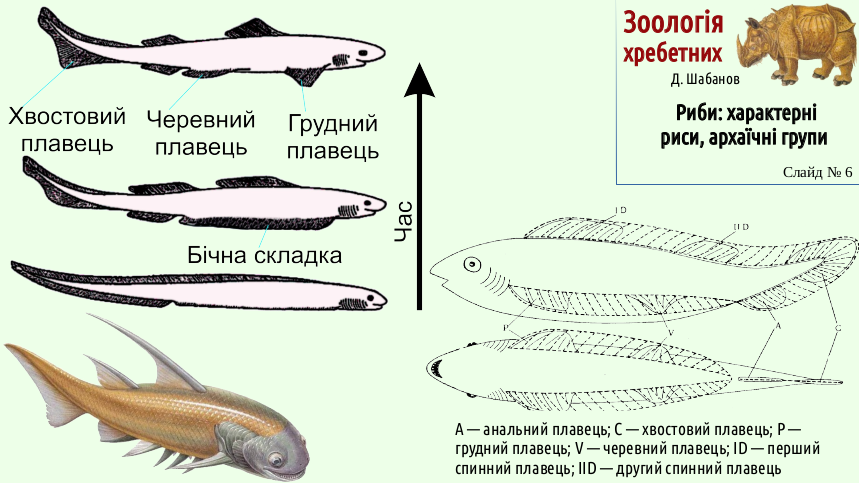
A sea lion in this video escaped from a great white shark by moving sideways. A smaller animal has difficulty competing with a large predator in speed, but it can win in maneuverability. So predatory feeding requires effective rudders that allow the body to turn during pursuit of prey. The predator's body is propelled forward by unpaired fins; the rudders must develop from other structures. The turns of fish ancestors were provided by paired lateral folds extending along the body (roughly where the metapleural folds run along the body of a lancelet).
In some acanthodians, we can see that the lateral folds were transformed into an entire series of paired fins. However, the effect of these fins as rudders was different. The anterior and posterior sections served as rudders, while the intermediate ones became mere spines that protected these predators from even larger predators. Thus, the fact that the body of the fish was turned more effectively by the anterior and posterior parts of the lateral fold, from which the pectoral and pelvic fins arose, is simply due to their greater efficiency under the lever rule. One consequence of this is that we belong to the group Tetrapoda: our four limbs are transformed pectoral and pelvic fins of fishes.
The acquisition of jaws and paired fins marked the formation of the characteristic body organization of fishes.
The first fishes looked roughly like this.
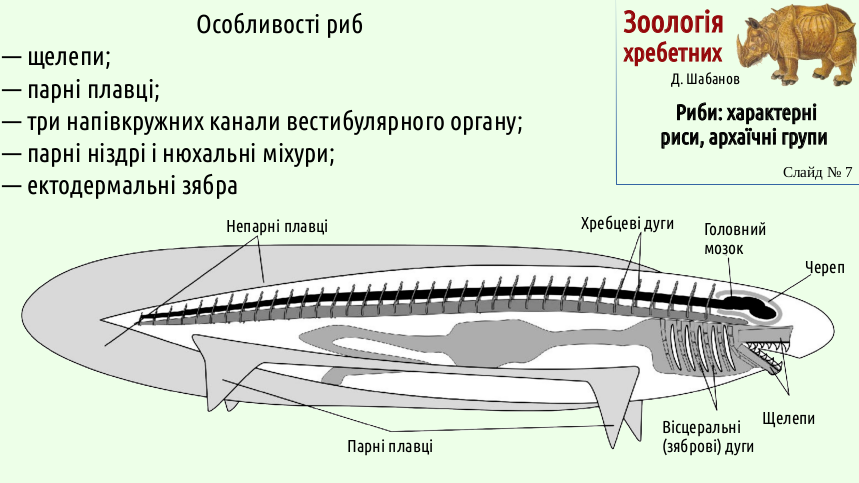
The slide below shows the archetype of a fish. In addition to jaws and paired fins, it shows a developed brain protected by a skull, well-developed sense organs, and vertebral arches providing support for powerful lateral muscles. Thus, we have discussed the main regularities in the emergence of fish organization. Jaws and paired fins are the result of predatory feeding. Three canals of the vestibular system are the result of increased control over movement; to track body position in three-dimensional space, one semicircular canal is needed for each spatial dimension.
Different groups of jawless vertebrates had different sets of sense organs. For example, lampreys descend from shield fishes that had a single nostril. Fishes descend from luckier shield fishes. Perhaps the fact that paired nostrils make it possible to determine from which direction a stronger smell is coming also played a role here. Are ectodermally derived gills more effective than endodermal gills? The author does not know the answer to this question. Maybe ectodermal gills give their owners some advantages, or maybe the development of gills in that group of shield fishes that became the ancestors of fishes was simply a random event.
With the emergence of the fish body plan, large invertebrate predators (such as sea scorpions and orthocerids mentioned in earlier lectures) soon disappeared: fishes became the dominant predators in the seas. Only much later did fishes give up part of the ecological space they had conquered to their descendants, tetrapods, who later returned to the water.
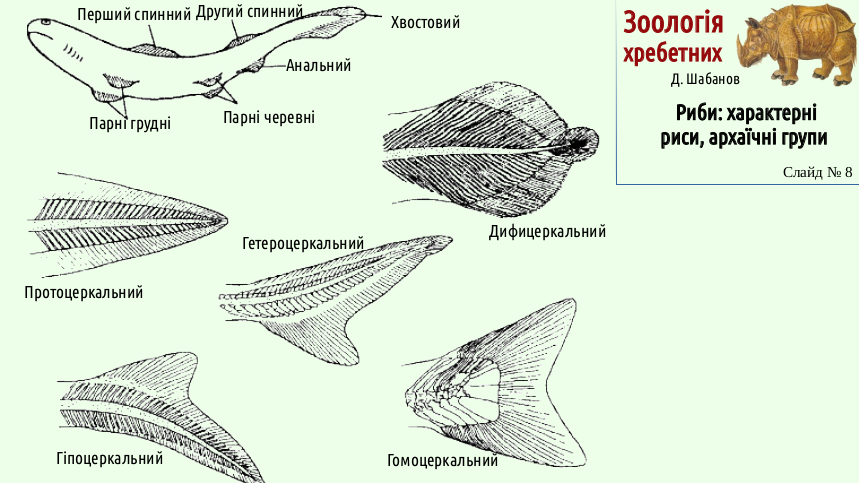
The changes did not affect only the paired fins. Intensification of swimming required a change in the shape of the caudal fin and affected the other unpaired fins.
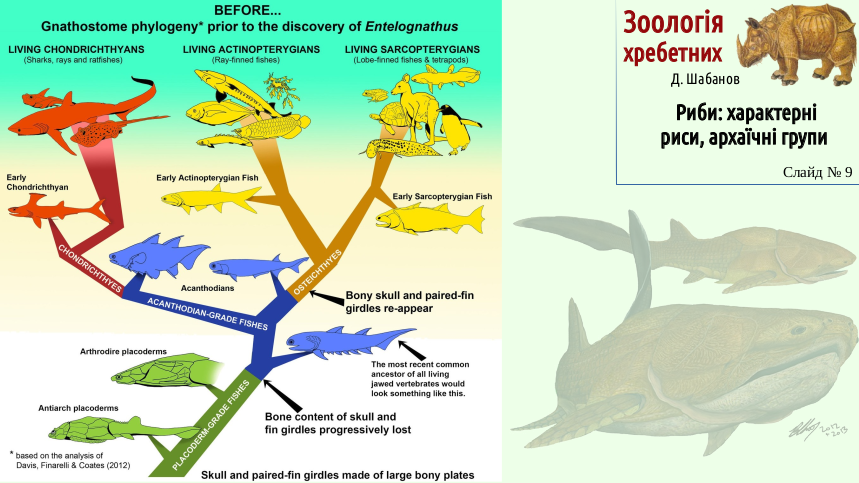
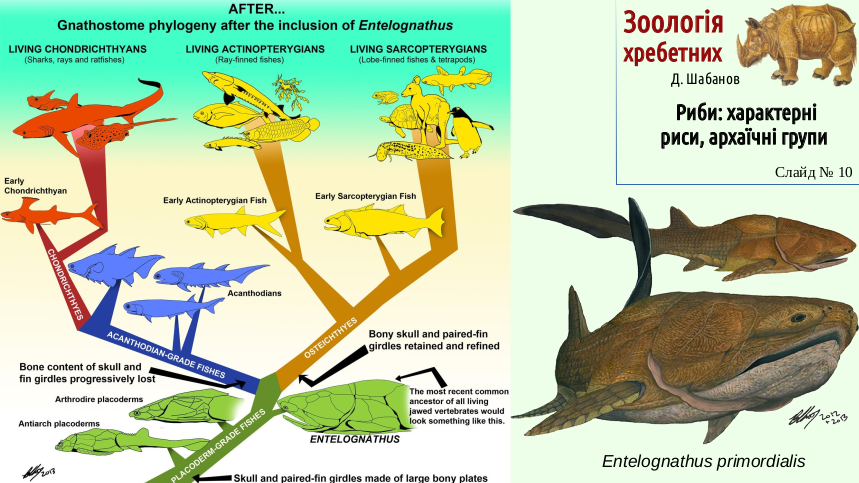
The phylogenetic relationships among the main groups of fishes remain controversial. New finds can substantially influence our ideas. The next two slides show ideas about the relationships among the main groups of fishes before the 2012 description of the primitive placoderm Entelognathus primordialis and after it. Entelognathus showed that primitive placoderms had important traits of bony fishes and may be regarded as an ancestral group for all fishes.
Thus, we consider fishes according to the following system.
Class Placodermi - Placoderms (Armored fishes, Plated-skinned fishes)
A group known from the Lower Silurian, that flourished in the Devonian and existed until the Permian. A diverse group that included about 9 orders. The largest reached 10 meters in length and were apex predators in Silurian and Devonian seas.
Placoderms had a body armor of dermal plates (tesserae), which most often covered the front part of the body. This armor is well preserved in sedimentary rocks. The main propulsive force of placoderms was the tail, which was usually less protected than the front part of the body. Overall, the typical body shape of placoderms resembled that of widespread groups of shield fishes.
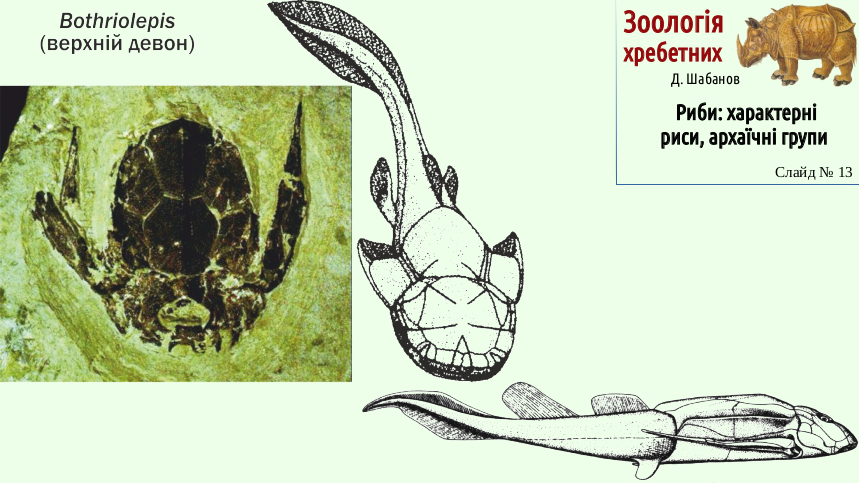
Placoderms were better swimmers than the shield fishes. The next slide shows one of the most famous placoderms - the Devonian Bothriolepis. Although the front of the body of Bothriolepis was covered by armor, it was probably not such a bad swimmer.
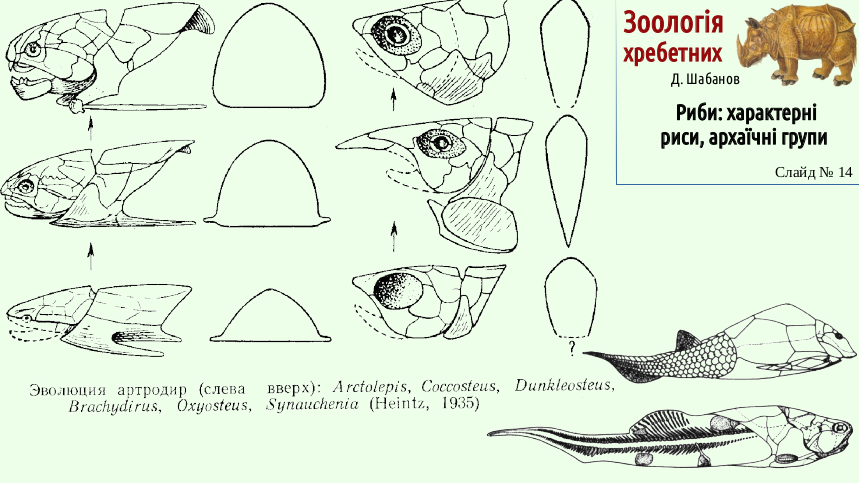
The diversity of placoderm body shapes is shown on the next slide. Among them were both bottom-dwelling forms with a flattened ventral side and inhabitants of the water column with laterally compressed bodies. Many placoderms resembled well-known modern ray-finned fishes. This was a consequence of belonging to similar life forms (a life form is a stable complex of adaptations to a particular way of life).
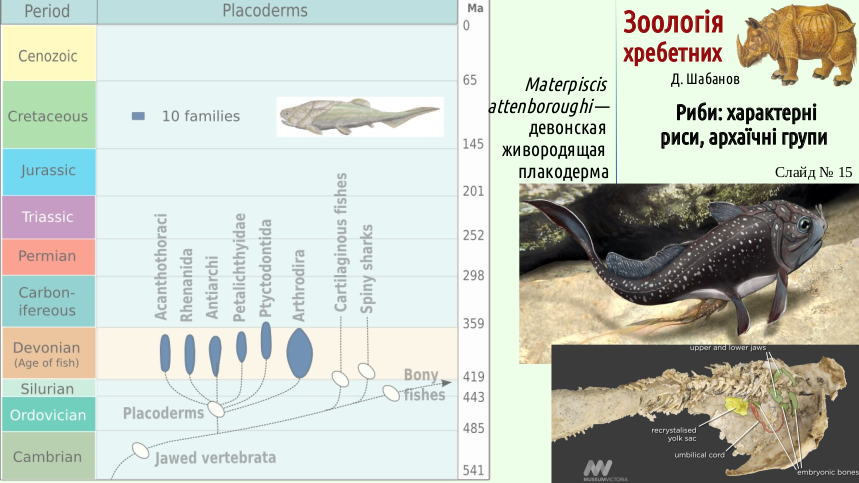
No matter how the phylogeny of placoderms is reconstructed, it is certain that they achieved considerable diversity. Among them were even viviparous representatives, one of which is named after Sir David Attenborough.
In the next video (from a fantasy series that imitates travels into the past), British television presenter Nigel Marven searches in the Devonian for the largest placoderm - Dunkleosteus (Dunkleosteus). Nigel offers Dunkleosteus a wrapped-up Bothriolepis to see whether Dunkleosteus's jaws can crush the armor. Placoderms had no true teeth, and prey was destroyed either by slanted plates that formed the head armor.
Class Acanthodii - Acanthodians (Spiny sharks)
A group that existed from the Silurian to the Permian. It included no truly large animals; the largest representatives reached 2 meters, but many were good swimmers that fed on plankton or smaller animals.
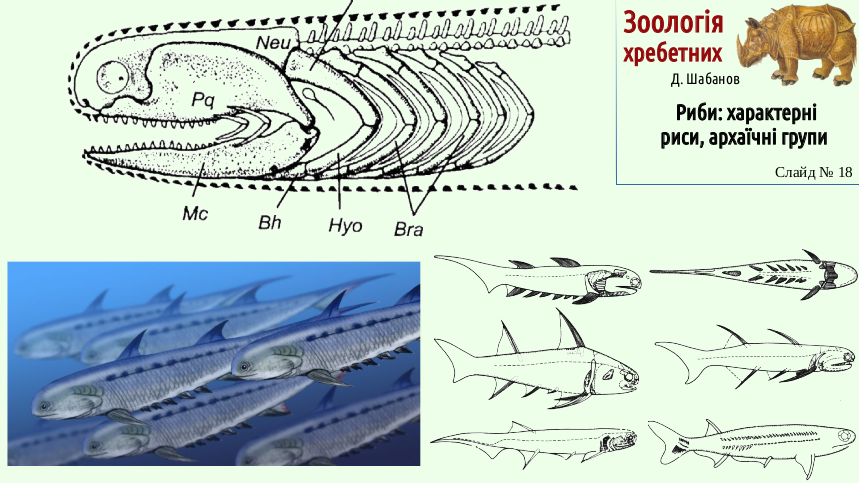
The name acanthodians emphasizes the similarity between the jaw arch and the gill arches; in acanthodians one can observe primitive variants of the visceral skeleton of fishes. Next to the jaw arch are labial cartilages - remnants of visceral arches located in front of the jaw. Notice the opening between the jaw arch and the hyoid arch. This is the spiracle, a rudiment of the gill slit that lay between these arches. The body shape of acanthodians was relatively uniform. A characteristic feature of their appearance was the large number of spines; this is reflected in their English name, spiny sharks.
It is quite likely that acanthodians, which fed on small prey, could form schools.