
Volume 6. Amphibians. 01. What are amphibians?
Grzimek`s Animal Life Encyclopedia. — 2nd ed. — Farminfton Hills, MI: Gale Group, 2003. Volume 6. Amphibians. — 507 p. What is an amphibian? — pp. 3-6. Working translation of the section from the largest international animal encyclopedia
Who are amphibians? Almost everyone knows fish, birds, or mammals, even reptiles. But what about amphibians? Most people recognize frogs and toads as amphibians, but these are not the only animals belonging to Amphibia, the class of vertebrate animals. There are three modern groups of amphibians. The most generalized (reflecting the original features of this group – note D.Sh.) are the salamanders, order Caudata (= tailed), which have a cylindrical body, a long tail, a distinct head and neck, and usually well‑developed limbs of roughly equal length. Most salamanders are terrestrial, but some are aquatic, a few are burrowing, and others are arboreal. Frogs, order Anura (= tailless), have a powerful torso merging into the head, lack a tail, and possess long hind limbs. Most tailless species are terrestrial or arboreal, but many are aquatic, and few are burrowing. The third group contains caecilians, order Gymnophiona, also called Apoda (= limbless). These limbless amphibians externally resemble earthworms and have blunt heads and tails, and their elongated bodies are encircled by grooves (rings). Several caecilians are aquatic, but most burrow in soil in tropical regions of the world. Description of features In a sense, amphibians occupy an intermediate position between fully aquatic fish and terrestrial amniotes (reptiles, birds, and mammals), but they are not a simple transition in their morphology, biology, ecology, and behavior. Over almost 350 million years of evolution, amphibians have undergone remarkable adaptive radiations, and their living groups demonstrate greater biological (life‑cycle) diversity than any other vertebrate group. In principle, amphibians can be defined as tetrapod vertebrates with a skull having two occipital condyles (articulating surfaces with the first element of the vertebral column). Attachment of the pelvic girdle to the spine is provided by a single sacral vertebra. In tailless amphibians (frogs and toads), the sacral vertebrae are fused into a rod‑like structure, the urostyle (coccyx), and the tail is absent. Caecilians and some salamanders lack limbs and their girdles, whereas in tailless amphibians the hind limbs are elongated and adapted for jumping. The skin is glandular and contains mucus and toxic glands, but lacks external structures such as scales, feathers, and hair that are characteristic of other tetrapod groups. The heart consists of three chambers, two atria and one ventricle, which may be partially divided. The aortic arches are symmetrical. Typically, amphibians have two lungs, but lungs may be reduced or absent in some salamanders, and the left lung is relatively smaller in most small caecilians (as in snakes). Some features are unique to amphibians, all of which have teeth composed of a base and crown, and specialized papillae for sound perception in the inner ear. Amphibians are ectothermic (cold‑blooded). They cannot physiologically regulate their body temperature as birds and mammals do, so their body temperature approximates that of their immediate environment, primarily the substrate. Biology (life cycle) Amphibian life cycles are highly diverse. The classic amphibian life cycle with aquatic eggs and larvae is only one of many reproductive modes, which also include direct development of terrestrial eggs (without an aquatic larval stage) and viviparity. Amphibian eggs lack shells and embryonic membranes (e.g., amnion, allantois, and chorion) found in reptiles, birds, and mammals. Instead, amphibian eggs are protected only by mucous capsules that are highly permeable; thus, amphibian eggs must develop in moist conditions. Phylogenetic relationships and classification Modern amphibian groups are closely linked with diverse fossil basal tetrapod vertebrates, usually placed in the class Amphibia. Phylogenetic links among these fossil groups are uncertain. Based on morphological and molecular evidence, salamanders and tailless amphibians form a monophyletic group (i.e., share a common ancestor) and together are called batrachians. Batrachians and caecilians form another monophyletic group, the lissamphibians. Classification reflects biologists’ knowledge of relationships among organism groups. Consequently, the acquisition and analysis of new data—morphological, molecular, behavioral, and embryological—alter classification. New data may show that groups of species or genera once assigned to a single family are actually more closely related to another group or not related to the former family. For example, salamanders of the families Dicamptodontidae and Rhyacotritonidae were previously placed in Ambystomatidae. Moreover, African tree frogs, currently treated as the family Hyperoliidae, were formerly included in Rhacophoridae, and frogs once considered the family Pseudidae are now treated as a subfamily of Hylidae.

Systematics (the study of evolution and classification of organisms) is a dynamically developing field, and the relationships of many groups remain unresolved. Depending on the type of evidence used, results can differ substantially, and various classifications may be proposed. Relationships of some modern amphibian groups have not been established with sufficient confidence. For instance, the group of endemic Madagascan frogs has been treated as the family Mantellidae, a subfamily of Ranidae, and a subfamily of Rhacophoridae (as adopted in this volume). In the classification used in this book there is the order Gymnophiona (caecilians) with five families, the order Caudata (tailed) with 10 families, and the order Anura (tailless) with 28 families.
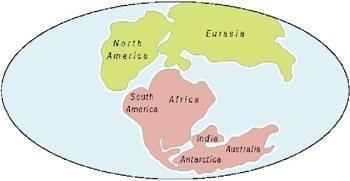
Hierarchical classification of modern amphibians and their relationships Historical biogeography The distribution of amphibian families reflects Earth’s history, especially the breakup of the supercontinent Pangaea, which began about 190 million years ago. Early separation produced two large continental masses: Laurasia, comprising North America, Europe, and most of Asia, and Gondwana, which included present‑day South America, Africa, Madagascar, the Indian subcontinent, Australia, New Zealand, and Antarctica. The ancestors of lissamphibians were probably widespread in Pangea before continental breakup. Continental arrangement in early Mesozoic (130 million years ago) Although fossil caecilians are known from the Jurassic of North America, today these amphibians live in regions that were parts of Gondwana. Two families are restricted to the Indian subcontinent (one also in adjacent Southeast Asia), one family is endemic to Africa, and one to South America. Tailed amphibians evolved in Laurasia. One family is limited to Asia, four families are shared across Eurasia and North America, where five endemic families occur. Only one Plethodontidae group dispersed from North to South America. Biogeography of tailless amphibians is somewhat more complex. One early branch, including modern Ascaphidae in North America and Leiopelmatidae in New Zealand, is linked to Jurassic South American fossils, indicating that this line split before the breakup of Pangaea. Fossil finds represent the distribution of other primitive tailless groups (archaobatrachians) in Laurasian continents: Bombinatoridae, Discoglossidae, Megophryidae, Pelodytidae and fossil Paleobatrachidae in Eurasia; Pelobatidae in Eurasia and North America, and Rhinophrynidae in North America. Nevertheless, the historical biogeography of most tailless amphibians (both primitive and advanced) is tied to Gondwana, whose fragmentation on existing continents played a major role in differentiating many tailless lineages. Many lineages are confined to a single continent: six families in South America, three in Africa, two in Australia, and one each in Madagascar and the Seychelles. Others are shared by two or more Gondwanan continents: one (Pipidae) in Africa and South America, one (Hylidae) in South America and Australia (also widespread in North America and Eurasia), one (Hyperoliidae) in Africa, Madagascar, and the Seychelles, and another (Rhacophoridae) in these three regions plus the Indian subcontinent and adjacent Southeast Asia. Microhylidae occurs on all Gondwanan continents except the Seychelles, and they dispersed into Southeast Asia and southern North America. True frogs (Ranidae) are worldwide except northern Australia and northern South America, and toads (Bufonidae) inhabit all continents except Australia (one species introduced). Regional diversity As a group, amphibians are distributed worldwide, excluding polar regions, most oceanic islands, and some desert areas. However, the pattern of their distribution differs among the three modern amphibian groups.
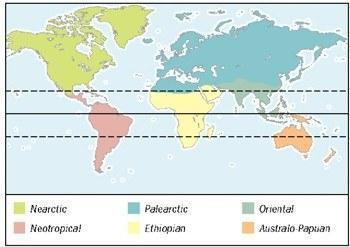
Tailless amphibians are worldwide but most diverse in the tropics; tailed amphibians are more diverse in northern continents; caecilians are limited to the tropics. Number of families/genera/species of amphibians in the world’s biogeographic regions. Non‑Arctic: 17/40/243; Neotropical: 19/185/2782; Palearctic: 15/34/192; Ethiopian: 13/95/770; Oriental: 12/75/825; Australo‑Papuan: 6/58/450 Globally, except for the Arctic and Antarctic (which lack amphibians), six biogeographic regions are recognized. The largest, the Palearctic (Europe and temperate Asia), has the fewest amphibian species (192), followed by the Non‑Arctic (temperate North America) with 243 species. Historically, these regions are part of former Laurasia and have the greatest diversity of tailed amphibians, especially in the Non‑Arctic. In contrast, the amphibian fauna of southern continents is mainly composed of tailless forms. The Australo‑Papuan region (Australia, New Zealand, New Guinea and associated islands) has 450 species of tailless amphibians, but none of tailed or caecilian groups. The Ethiopian (Afrotropical) region (sub‑Saharan Africa and Madagascar) has 770 species, including 29 caecilians. The Oriental region (tropical and subtropical Southeast Asia, India and associated islands) shelters 825 species, including 29 tailed salamanders and 44 caecilians. Undoubtedly, the Neotropical region (South America, tropical Mesoamerica, and the West Indies) has the greatest amphibian diversity, with 82 caecilian species, about 200 salamander species, and over 2 500 tailless species. Although caecilians are pantropically dispersed, they are absent from Madagascar. Ichthyophiidae and Uraeotyphlidae are endemics of the Oriental region, Scolecomorphidae of the Ethiopian region, and Rhinatrematidae of the Neotropical region. The large family Caeciliidae is most diverse in the Neotropical region (14 genera and 73 species) and also occurs in Africa (6 genera, 17 species), the Oriental region (2 genera, 4 species) and the Seychelles (3 genera, 7 species). Most tailed amphibians inhabit the Northern Hemisphere; they are absent from the Australo‑Papuan and Ethiopian regions. At the family level, the greatest diversity is in the Non‑Arctic region, where all families (except Hynobiidae) occur, and five families (Ambystomatidae, Amphiumidae, Dicamptodontidae, Rhyacotritonidae, and Sirenidae) are endemic. Cryptobranchidae and Proteidae are each represented by a single genus in the Non‑Arctic and Palearctic regions. Salamandridae is the most widespread tailed family with five genera in the Palearctic and two genera in the Non‑Arctic. Hynobiidae is the only tailed family confined to the Palearctic. Undoubtedly, the largest tailed family is Plethodontidae with 25 genera in the Non‑Arctic; one of these (Hydromantes) is shared with Europe. However, the greatest diversity of tailless amphibians occurs in tropical America, especially Central America and southern Mexico, where two genera exceed 200 species; two of these genera also occur in South America, one south of Bolivia.

Yellow‑legged frog (Rana boylii) metamorphosing from tadpole in Mendocino County, California Broad‑snouted frog (Amietia vertebralis) found at high altitude—over 10,000 ft (3,000 m)—in South Africa.

A nictitating membrane in each eye protects against ultraviolet radiation Only four of the 28 tailless families are simultaneously present in the Old and New Worlds. Bufonidae is globally distributed, absent only from Australia, New Zealand, and Madagascar. Ranidae shows a similar pattern but also occurs in Madagascar and northern Australia. Microhylidae has several representatives in the Non‑Arctic and Palearctic regions and is more diverse on southern continents, including Madagascar and New Guinea, but not New Zealand. Hylidae is most diverse in the Neotropical region and secondarily in the Australo‑Papuan region. Two genera are endemic to North America, and only a few Hyla species occur in the Oriental and Palearctic regions. Except for Pelobatidae in the Non‑Arctic and Palearctic, other tailless amphibian families are restricted to the New World or the Old World, and only some occur in the Northern Hemisphere. Ascaphidae are endemic to the Non‑Arctic, Megophryidae to the Oriental, Discoglossidae and Pelodytidae to the Palearctic, and Bombinatoridae to the Palearctic and Oriental. The greatest diversity is characteristic of southern landmasses. Leiopelmatidae are endemic to New Zealand, and Limnodynastidae and Myobatrachidae are endemic to the Australo‑Papuan region. The Ethiopian region has six endemic tailless families: Arthroleptidae, Heleophrynidae, Hemisotidae, Hyperoliidae (Africa, Madagascar, and the Seychelles), Scaphiophrynidae (Madagascar only) and Sooglossidae (Seychelles only). The highest diversity of Rhacophoridae is in the Oriental region, but this family is also diverse in Madagascar and has one genus with three species in Africa. The Neotropical region has the world’s greatest tailless diversity. In addition to many genera and species of Bufonidae, Hylidae, and Microhylidae, it hosts seven endemic families: Allophrynidae, Brachycephalidae, Centrolenidae, Dendrobatidae, Leptodactylidae, Rhinodermatidae, and Rhinophrynidae. Four of them (Allophrynidae, Brachycephalidae, Rhinodermatidae, and Rhinophrynidae) contain only eight species, but Centrolenidae and Dendrobatidae together comprise over 300 species, and Leptodactylidae includes more than 1 000 species, with the genus Eleutherodactylus being the most numerous and widespread. Other families in the Neotropical region are Pipidae (shared with Africa) and Ranidae (shared with most of the world). Marsupial marsupial frog (Gastrotheca riobambae) does not produce and lay egg clutches like most tailless amphibians, but has a different reproductive mode.

The male fertilizes the eggs externally and then places them on the female’s back. The female carries the eggs until they reach the tadpole stage, then deposits them in water. This female carries the eggs in a pouch. Amphibians in ecosystems Although amphibians are generally confined to moist habitats such as wet forests, swamps, ponds, and streams, many species venture far from open water and inhabit trees, rocks, and soil beneath the ground surface. In such varied habitats amphibians feed on a great multitude of small organisms, mainly invertebrates, with insects being the most numerous component of the diet of both tailless and tailed groups. Nevertheless, their diets also include earthworms (especially in caecilians), small snails, spiders, and other tiny invertebrates. Body size plays an important role in prey selection. Some aquatic tailed species feed on tadpoles, and several large aquatic salamanders eat fish; eel‑like aquatic amphibians feed almost exclusively on crustaceans. Many frog species are less than 1 inch (25 mm) long, and their diet is limited to small insects and spiders. In tropical forests many of these small tailless frogs specialize on ants and termites, which are encountered at every turn. Large frogs with wide mouths generally eat larger prey, which may include other frogs, lizards, small snakes, birds, and mammals. Tadpoles feed primarily on decaying vegetation, algae, and plankton in ponds and streams. The feeding habits of amphibians play a crucial role in ecosystems because, as adults, they consume huge numbers of insects and thus help maintain ecological balance. Areas where native tailless amphibians have been eliminated have experienced significant increases in certain insect species, and mountain streams that were once relatively free of algae can become clogged with algae when algae‑grazing tadpoles disappear. Because of their abundance and relative ease of capture, amphibians become prey for many animals, especially small mammals, birds, and many snake species. Wetland birds feed on tadpoles and metamorphosing frogs in shallow waters. Several snake species specialize on tailed amphibians, and many tropical snakes feed almost exclusively on frogs. Small tailed and tailless amphibians fall prey to spiders. Even subterranean caecilians cannot escape predation by some snakes, especially coral snakes of the genus Micrurus. Thus amphibians are an important component of food webs in most terrestrial ecosystems on the planet. In the late 1980s biologists recorded a decline in amphibian populations worldwide.Gradual, and especially rapid, declines will not only lead to the potential loss of amphibian species, but will also have a significant impact on the populations of their prey, their predators, and animals positioned higher in the food chain. The long‑term consequences of such declines have not yet been determined.