
Ecology: Biology of Interactions. V-15. Clinal Variation and Some Ecological Rules
Appendices: Syllabus. Questions. References. Personalities. Glossary. R Commands.
V-15. Clinal Variation and Some Ecological Rules
By comparing conditions in different parts of the Earth's surface, we can confirm that many important ecological factors change gradually, forming a gradient (smooth) sequence of changes. For example, travelling in a given direction, we may observe how elevation above sea level decreases gradually while humidity and temperature increase. At some boundary, a discrete (abrupt) change in environmental conditions will occur — for instance, we pass from land to sea. Yet as we continue our movement, now across the sea, we again observe that many factors (depth, light intensity) change in a gradient fashion.
When comparing organisms that inhabit a region characterised by gradient change in any given factor, we can detect a regular change in various traits of those organisms. When referring to intraspecific variation, such gradual geographic variability is termed clinal. The traits themselves may be said to form clines. For example, in broadleaved forests, in the forest-steppe, and even in steppe and semi-desert zones, we can find oak woodlands — oak forests. The transition between these zones is determined primarily by the amount of water available to plants. In the broadleaved forest zone, oaks grow on level surfaces unconnected with river channels, whereas in steppe and semi-desert they are confined to depressions (ravines) and river valleys — localities with elevated moisture. Comparing oaks from these woodlands, we find that numerous traits vary among them: height, stem diameter, leaf size, and features of leaf blade structure. These differences are linked to plant responses to gradually changing climatic factors.
The study of clinal variation is of particular interest for elucidating the nature of organism-environment relationships (Fig. V-15.1). Clinal variation is the result of selection adapting each population to local environmental conditions, and of interactions between neighbouring populations (crossbreeding, migration) that smooth out differences between them. In effect, clinal variation "reveals" and renders visible and accessible for study these processes that are usually hidden from the researcher.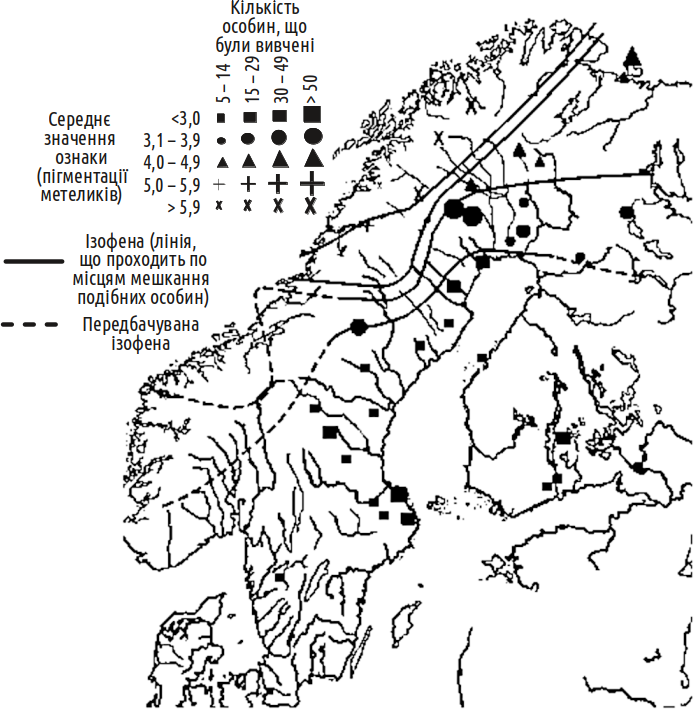
Fig. V-15.1. This diagram shows the clinal variation in wing pigmentation of one butterfly species, the green-veined white (Pieris napi), based on a study conducted at the beginning of the twentieth century. Isophenes are lines connecting areas of occurrence of organisms with the same degree of expression of the trait under investigation.
As a rule, clinal variation in organisms is not considered a basis for distinguishing subspecies. Geographic subspecies (forms within a species having independent systematic status) are recognised in cases of discrete (discontinuous, stepwise) variation. Nevertheless, transitional zones — intergradation zones — sometimes exist between subspecies or even between species capable of hybridisation.
In the majority of cases the concept of the cline is applied to describe intraspecific variation, although analogous changes can be recorded when examining the diversity of different species within a single genus or even family, provided they follow a similar mode of life and inhabit an environment exhibiting a gradient in any factor important to those organisms.
Some rules describing clinal and also interspecific geographic variation were formulated quite long ago. For example, Bergmann's Rule and Allen's Rule, which pertain both to intraspecific variation and to differences between closely related species with similar modes of life, were already formulated in the nineteenth century.
Bergmann's Rule (1847): among related forms of homeothermic animals with similar modes of life, those inhabiting colder climates have larger body sizes.
Allen's Rule (1877): among related forms of homeothermic animals with similar modes of life, those inhabiting colder climates have relatively smaller protruding body parts: ears, legs, tails, and so on.
For instance, the geographic variation of the common red fox conforms to both of these rules. The linear body dimensions of southern foxes are approximately 10–15% smaller than those of northern foxes. Furthermore, southern foxes have relatively longer ears and tails. If we go beyond this species and examine the more northerly and more southerly relatives of foxes, we see that they obey the same rule. Arctic foxes have a fairly large body with short limbs, muzzle, tail, and ears. Desert fennec foxes are rather small, with long limbs, muzzle, and tail and very large ears.
The largest tigers are Amur tigers, which inhabit the harshest climate, while the smallest are Sumatran tigers. Wolves inhabiting the tundra can reach 50 kg in weight, whereas in deserts their weight amounts to only 35 kg. Bergmann's Rule also applies to interspecific geographic variation. Bears conform to it admirably — from the gigantic polar bear and grizzly to the smaller Asiatic black bear. The largest penguins (the emperor and king) inhabit Antarctica, while the smallest are found on the Galapagos Islands.
Bergmann's Rule and Allen's Rule share a similar physiological basis: they illustrate the fact that in cold climates, animals with a smaller relative body surface area find it easier to maintain a constant body temperature. Heat loss through the body surface is proportional to its surface area, while heat capacity and heat production are proportional to body volume. The relative surface area can be reduced either by increasing the linear body dimensions or by "rounding" the body — that is, by reducing its protruding parts.
Like all rules, Bergmann's Rule and Allen's Rule have numerous exceptions. Fossorial mammals, for example, do not conform to them. Their body size and proportions are substantially influenced by the requirements of locomotion in the soil environment, which, among other things, protects them from cold air.
For instance, elephants conform to Allen's Rule but not to Bergmann's Rule. African elephants are larger than Indian elephants, yet they inhabit a hotter climate. This is related to the fact that African elephants live predominantly in open spaces (savannah), while Indian elephants inhabit forests. Notably, forest-dwelling African elephants are smaller than Indian elephants! On the other hand, as would be expected, African elephants have greater difficulty dissipating excess heat, and therefore have much larger ears than Indian elephants.
Can animals following dissimilar modes of life be compared with one another? Can we assert that Bergmann's Rule is disproved by, for example, tropical fruit bats (flying foxes, flying dogs, etc.), which are considerably larger than bats of temperate latitudes? Of course, such a conclusion would be incorrect. These animals follow fundamentally different modes of life. Fruit bats feed primarily on fruit, while our bats specialise in catching nocturnal flying insects. The different feeding habits lead to substantial differences in the thermal balance of these animals, which in turn determines the differences in their body size.
Can Bergmann's Rule and Allen's Rule be applied to intraspecific variation in humans? Although all people belong to a single species, our ecological plasticity is so great that in different parts of the range humans lead different modes of life. These differences impede the manifestation of Bergmann's Rule. At the same time, when many peoples are compared, manifestations of Allen's Rule can be discerned. Inuit and other indigenous peoples of the Far North, for example, have stocky bodies with short arms, legs, and necks. Inhabitants of the open spaces of equatorial Africa (e.g., the Maasai) are slender and have relatively long and thin legs and arms. Of course, the manifestations of Allen's Rule can be observed only in indigenous peoples living in close interaction with their native environment. Representatives of modern global humanity, many of whom have migrated throughout the world and live in artificially modified environments, do not typically exhibit the manifestations of these ecological rules.
Another well-known ecological rule describing clinal variation is Gloger's Rule, proposed in 1833 by the ornithologist C. W. L. Gloger. It states that among related forms (subspecies or species) of homeothermic animals, those inhabiting warm and humid climates are more deeply pigmented than those inhabiting cold and dry climates.
Plausible reasons leading to this pattern of variation may include considerations related to cryptic coloration: the colour of the soil (or the snow surface) in cold and dry climates is generally lighter than in warm and humid ones. However, this circumstance alone is insufficient to explain Gloger's Rule, since it extends even to nocturnal animals. It is possible that a warm and humid climate promotes the synthesis of animal pigments. To some extent, Gloger's Rule can also be applied to humans.