
Ecology: Biology of Interaction. V-14. The Concept of Effective Temperatures
V-14. The Concept of Effective Temperatures
We have already noted on numerous occasions that temperature is one of the most important ecological factors. One reason for this is that the rate of chemical reactions depends substantially on temperature. For an approximate assessment of this effect, the Van't Hoff rule may be applied: the rate of chemical reactions doubles or triples with every 10°C increase in temperature. Environmental temperature has a particularly pronounced effect on poikilothermic organisms. Changes in the rate of certain biological processes conform to the Van't Hoff rule. For example, the intensity of carbon dioxide release by soil (which depends on the activity of soil bacteria), the locomotion rate of centipedes, and intestinal peristalsis in caterpillars all follow this pattern closely.
In certain cases, the temperature regime regulates development. The phenomenon of vernalisation is well known in wheat and other plants. To explain this phenomenon, it should be noted that wheat (and indeed rye, cabbage, and similar plants) occurs in both winter and spring forms. Under warm and moist conditions, winter-form plants begin to germinate but subsequently arrest their development, awaiting the onset of winter cold. After exposure to cold for one to three months, winter-form plants resume development and ultimately proceed to flowering and fruiting. Spring-form plants lack this phase of their life cycle and are capable of completing their entire development at positive temperatures. Vernalisation consists in the fact that exposure to low positive temperatures (for example, holding at temperatures between +1°C and +10°C for several days) induces winter-form plants to resume normal development culminating in flowering and fruiting. For a number of organisms, exposure to low temperatures (cold diapause) is a prerequisite for the onset of development of seeds (in plants), eggs (in insects or crustaceans), or other resting stages.
However, even once the developmental process has been initiated, its rate remains dependent on temperature. The development of many poikilothermic organisms is well described by the concept of effective temperatures, which treats the quantity of heat as a resource necessary for development.
The study of the effect of heat quantity on development was initiated by the eminent physicist René-Antoine Ferchault de Réaumur as early as 1735. Having been tasked with determining why fruit crops develop differently in different regions of France, he established that the quantity of heat required for plant development can be calculated by simply summing the mean daily temperatures over the warm period of the year. If the summation yields the quantity of heat required for a given variety, it will ripen; if not, it will fail to do so in time.
Initially, in investigations of the required heat quantity, positive temperatures (i.e., those exceeding 0°C) were summed; however, it gradually became apparent that a different temperature threshold must be employed for certain species. This eventually necessitated the introduction of the concept of the physiological zero. The physiological zero (T0) is the temperature above which development of a poikilothermic organism commences.
Although the physiological zero values for many organisms are close to 0°C, animals inhabiting snowy environments grow and develop at negative temperatures, while certain other organisms cease growth and development at positive temperatures. The physiological zero can be determined more precisely by examining the dependence of the rate of development on temperature, as shown in Fig. V-14.1.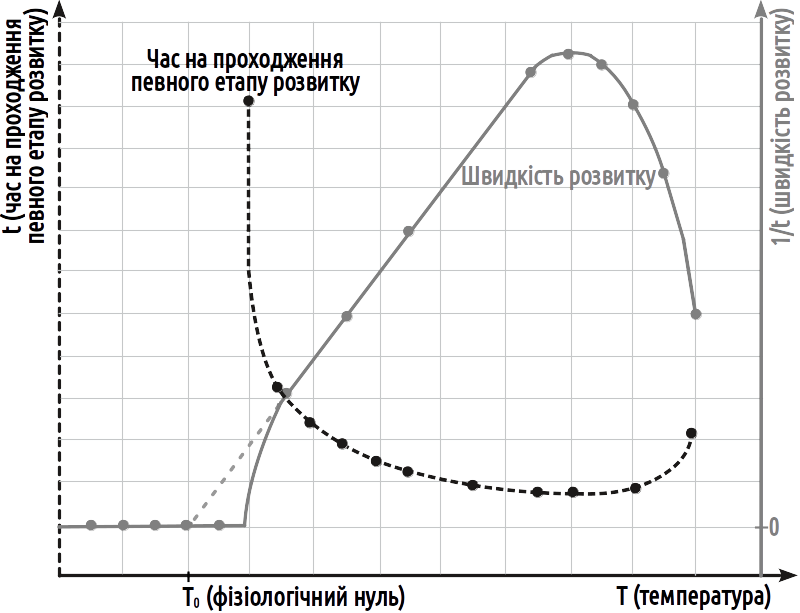
Fig. V-14.1. Dependence of the rate and duration of development on temperature in poikilothermic organisms. A certain portion of the dependence of the rate of development on temperature may be regarded as linear; the duration of development over this portion changes hyperbolically. Within this portion, calculation of the rate and duration of development using the concept of effective temperatures is feasible. The physiological zero is the point at which this line intersects the zero rate of development level.
The dependence of the duration of development of a poikilothermic organism on temperature is hyperbolic in character. However, the rate of development (the quantity inverse to duration) depends linearly on temperature over a fairly wide range of values of this factor. Incidentally, it is evident from the figure that this character of dependence is not always preserved at very low temperatures close to T0. This means that the definition of the physiological zero given above is not entirely precise. On the graph, this definition corresponds to a temperature exceeding 5°C, whereas T0 = 4°C. The physiological zero should be calculated by extending the linear portion of the dependence of the rate of development on temperature to the zero rate of development level. Moreover, it should be noted that the physiological zero may differ for different developmental stages (this circumstance is not reflected in the figure).
Having clarified the concept of the physiological zero, the concept of the effective temperature may now be defined. The effective temperature (Te) is the difference between the physical temperature (T) and the physiological zero (T0): Te = T − T0.
The essence of the concept of effective temperatures is that the completion of each developmental stage of a poikilothermic organism requires a specific sum of mean daily effective temperatures, referred to as the thermal constant or heat constant (TConst). Examples of physiological zero values and thermal constants for certain organisms are presented in Table V-14.1.
Table V-14.1. Examples of thermal developmental parameters for certain organisms
|
Table 5.20.1. Examples of thermal parameters for the development of selected organisms |
Organism | Physiological Zero (T₀) | Developmental Stage | Thermal Constant (TConst)T0) |
Table 5.20.1. Examples of thermal parameters for the development of selected organisms |
Colorado potato beetle | +13.5 °C | Full cycle | 285 degree‑days |
Thermal constant (TConst) |
|
|
Cod (roe) | –3.6 °C | Roe development | 150 degree‑days |
Colorado potato beetle |
Cod (roe) | –3.6 °C | Roe development | 150 degree‑days |
Colorado potato beetle |
Complete developmental cycle |
|
|
Eurasian Scops Owl Agrotis segetum |
Agrotis segetum moth |
||||
|
+10°C |
Atlantic cod (eggs) |
Eurasian Scops Owl Agrotis segetum |
Colorado potato beetle |
Complete developmental cycle |
|
|
Glacier (Boreus westwoodi) |
Scorpionfly (Boreus westwoodi) |
||||
|
Are calculations based on the effective‑temperature concept always accurate? No. The described logic applies only to narrow temperature ranges, not to all organisms or all processes within them. The most significant drawback of this concept is the discrepancy between development times under constant versus variable temperatures. For instance, a chicken egg develops in 21 days at 40–41 °C; increasing temperature yields no gain. The Colorado potato beetle develops in 23 days at 20 °C, 15 days at 25 °C, and 19 days under fluctuating temperatures, regardless of whether the mean temperature is 20 °C or 25 °C. Variable temperatures are more natural! |
Wheat |
0°C |
Educational model: Influence of Heat Accumulation on Development of Poikilothermic Organisms |
Egg development |
|
|
D. Shabanov, M. Kravchenko. Ecology: Biology of Interaction |
Scots pine |
||||
|
5.21. Clinal Variability and Some Ecological Rules |
Pea |
||||
|
–2°C |
Chicken egg |
||||
V-14. The Concept of Effective Temperatures
We have already noted on numerous occasions that temperature is one of the most important ecological factors. One reason for this is that the rate of chemical reactions depends substantially on temperature. For an approximate assessment of this effect, the Van't Hoff rule may be applied: the rate of chemical reactions doubles or triples with every 10°C increase in temperature. Environmental temperature has a particularly pronounced effect on poikilothermic organisms. Changes in the rate of certain biological processes conform to the Van't Hoff rule. For example, the intensity of carbon dioxide release by soil (which depends on the activity of soil bacteria), the locomotion rate of centipedes, and intestinal peristalsis in caterpillars all follow this pattern closely.
In certain cases, the temperature regime regulates development. The phenomenon of vernalisation is well known in wheat and other plants. To explain this phenomenon, it should be noted that wheat (and indeed rye, cabbage, and similar plants) occurs in both winter and spring forms. Under warm and moist conditions, winter-form plants begin to germinate but subsequently arrest their development, awaiting the onset of winter cold. After exposure to cold for one to three months, winter-form plants resume development and ultimately proceed to flowering and fruiting. Spring-form plants lack this phase of their life cycle and are capable of completing their entire development at positive temperatures. Vernalisation consists in the fact that exposure to low positive temperatures (for example, holding at temperatures between +1°C and +10°C for several days) induces winter-form plants to resume normal development culminating in flowering and fruiting. For a number of organisms, exposure to low temperatures (cold diapause) is a prerequisite for the onset of development of seeds (in plants), eggs (in insects or crustaceans), or other resting stages.
However, even once the developmental process has been initiated, its rate remains dependent on temperature. The development of many poikilothermic organisms is well described by the concept of effective temperatures, which treats the quantity of heat as a resource necessary for development.
The study of the effect of heat quantity on development was initiated by the eminent physicist René-Antoine Ferchault de Réaumur as early as 1735. Having been tasked with determining why fruit crops develop differently in different regions of France, he established that the quantity of heat required for plant development can be calculated by simply summing the mean daily temperatures over the warm period of the year. If the summation yields the quantity of heat required for a given variety, it will ripen; if not, it will fail to do so in time.
Initially, in investigations of the required heat quantity, positive temperatures (i.e., those exceeding 0°C) were summed; however, it gradually became apparent that a different temperature threshold must be employed for certain species. This eventually necessitated the introduction of the concept of the physiological zero. The physiological zero (T0) is the temperature above which development of a poikilothermic organism commences.
Although the physiological zero values for many organisms are close to 0°C, animals inhabiting snowy environments grow and develop at negative temperatures, while certain other organisms cease growth and development at positive temperatures. The physiological zero can be determined more precisely by examining the dependence of the rate of development on temperature, as shown in Fig. V-14.1.
Fig. V-14.1. Dependence of the rate and duration of development on temperature in poikilothermic organisms. A certain portion of the dependence of the rate of development on temperature may be regarded as linear; the duration of development over this portion changes hyperbolically. Within this portion, calculation of the rate and duration of development using the concept of effective temperatures is feasible. The physiological zero is the point at which this line intersects the zero rate of development level.
The dependence of the duration of development of a poikilothermic organism on temperature is hyperbolic in character. However, the rate of development (the quantity inverse to duration) depends linearly on temperature over a fairly wide range of values of this factor. Incidentally, it is evident from the figure that this character of dependence is not always preserved at very low temperatures close to T0. This means that the definition of the physiological zero given above is not entirely precise. On the graph, this definition corresponds to a temperature exceeding 5°C, whereas T0 = 4°C. The physiological zero should be calculated by extending the linear portion of the dependence of the rate of development on temperature to the zero rate of development level. Moreover, it should be noted that the physiological zero may differ for different developmental stages (this circumstance is not reflected in the figure).
Having clarified the concept of the physiological zero, the concept of the effective temperature may now be defined. The effective temperature (Te) is the difference between the physical temperature (T) and the physiological zero (T0): Te = T − T0.
The essence of the concept of effective temperatures is that the completion of each developmental stage of a poikilothermic organism requires a specific sum of mean daily effective temperatures, referred to as the thermal constant or heat constant (TConst). Examples of physiological zero values and thermal constants for certain organisms are presented in Table V-14.1.
Table V-14.1. Examples of thermal developmental parameters for certain organisms
In essence, the sum of effective temperatures constitutes a measure of the physiological time of a poikilothermic organism.
Accordingly, if the development of a poikilothermic organism proceeds at a variable temperature, the mean daily effective temperature must be calculated for each day. The condition for the completion of a developmental stage is that the sum of effective temperatures reaches the value of the thermal constant. At a constant developmental temperature, the calculations are simplified: one may simply state that TConst = t × Te, and also TConst = t × (T − T0), where t is the duration of development (in days).
The unit of measurement for the sum of effective temperatures and the thermal constant is degree-days (less precisely, degree-days), the product of degrees Celsius and the number of days.
Naturally, when calculating the quantity of heat received by an organism, it is meaningful to account for effective temperatures only for those days on which T did not fall below T0, since when the temperature drops below the physiological zero, development is arrested. However, if during part of a given day the temperature exceeded the critical threshold, time should be measured not in days but in other units (for example, in hours). Hours rather than days should also be employed for processes proceeding at high rates.
"For assessing the rate of development of microorganisms, the use of 'degree-hours' is possible, as any housewife familiar with yeast fungi knows. At higher temperatures they develop more intensively, and therefore the dough or kvass will be ready sooner than at low temperature. Temperature also affects the intensity of reproduction of lactic acid bacteria: milk that remains fresh for a long time in a refrigerator turns sour within a few hours in a warm room" (B.M. Mirkin, L.G. Naumova, 2005).
From the foregoing it follows that, knowing the duration of development of an organism at two different temperatures (corresponding to the linear portion of the dependence in Fig. V-14.1), we can determine the duration of its development at other temperatures falling within this linear range. Suppose that at temperature T1 the organism develops over t1 days, and at temperature T2 over t2 days. Since t1 × (T1 − T0) = t2 × (T2 − T0) = TConst, it follows that t1T1 − t1T0 = t2T2 − t2T0, and therefore t2T0 − t1T0 = t2T2 − t1T1. Having determined T0, TConst is readily determined. The duration of development at temperature T3 can be found using the formula t3 = TConst / (T3 − T0).
For example, we know that grasshopper eggs complete their development at 20°C in 17.5 days, and at 30°C in 5 days. Substituting the corresponding values into the formula t1 × (T1 − T0) = t2 × (T2 − T0), we can establish that 17.5 × (20 − T0) = 5 × (30 − T0), from which it follows that T0 = 16°C and TConst = 70 degree-days. On this basis, it can be concluded that development at 23°C would take, for instance, 10 days.
Are calculations based on the concept of effective temperatures always accurate? No. The logic described above can be applied only to certain temperature ranges, not to all organisms, and not to all processes within those organisms. The most significant shortcoming of this concept lies in the difference between developmental durations at constant and variable temperatures. Thus, a chicken egg develops in 21 days at a temperature of 40–41°C, and an increase in temperature yields no gain. The Colorado potato beetle develops at 20°C in 23 days, at 25°C in 15 days, and under variable temperature conditions in 19 days, regardless of whether the mean value equalled 20°C or 25°C. Variable temperatures are more natural!
Nevertheless, even simple mathematical models confer an advantage in simplifying the management of biosystems. Imagine that you need to plan an agronomic crop-protection measure against a particular pest, and that this measure will be most effective at a specific developmental stage of the organism (for example, before its transition to pupation). Knowing the time of mass oviposition and the weather forecast, you will be able to determine in advance the approximate date on which your efforts will be most effective. Does it matter that your calculations will contain some degree of imprecision?