
Ecology: biology of interaction. IV-18. (supplement) Strategies of intraspecific interaction
Almost all offspring in a troop of baboons belongs to several dominant males (leaders) who, by combining their efforts, keep subdominant males in a subordinate state (in particular, thereby prohibiting them from mating with females). Periodically, as the leaders age, «revo...
IV-18. (supplement) Strategies of intra‑population interaction
It can be said that a large part of organisms lives in two fundamentally different environments: the external one and the intra‑population one. The intra‑population environment is the set of relationships among relatives that are not exhausted by the examples given below.
One of the most important mechanisms of intra‑population regulation of numbers is territoriality—competition among individuals of a population for the use of space with all its resources. Territoriality often manifests in the defense of an individual’s territory. Most often the area of this territory is larger than the minimum required for the survival of the individual or its family. In different organisms territoriality is expressed in different ways. Sometimes only males guard individual patches, sometimes pairs, sometimes both males and females compete with each other for individual patches.
Have you ever wondered why songbirds sing? At the pair‑formation stage, males sing to attract females. Yet after pairs are formed, nests built, chicks hatched, males continue to sing... At this stage the male’s song indicates that the family patch is occupied. Strangely enough, biologically this behavior is quite similar to that of a domestic dog, which at the other end of the leash, during a walk with its owner, marks every corner of the house, a post or a tree with its urine. In most cases territorial individuals know where their patch ends and where neighbors’ patches begin. The position of the border is the result of a balance of effort by the owners of adjacent territories to expand their own patches. Violation of another’s territory provokes clashes, and the outcome of these clashes usually depends on whose territory they occur in.
“The complex of territorial behavior is not exhausted by direct attacks, fights, chases, etc. Moreover, such harsh forms of defense in their pure form are quite rare; almost always aggression is accompanied by ritualized behavioral forms: threat postures, specific acoustic signals, demonstration of attack without actual physical contact, and so on. The significance of the threat includes, for example, a certain torso tilt toward the horizontal in a range of fish species, ‘elevated’ postures of some tailless amphibians, head‑bobbling, raising and lowering the forebody, and spreading of the throat fold in iguanid lizards, various forms of display in different mammals. Mostly, threat displays are perceived by the individual that has entered a foreign patch as a signal to flee” (I.O. Shylov, 1998).
Most often in a territorial animal population not all individuals reproduce, but only those that possess an individual patch. “Rovers” are forced to remain childless, waiting for an opportunity to occupy a territory that becomes vacant as a result of predator action or to wrest a patch from an aging territory holder. This behavior is an effective way to stabilize population size: at any moment the number of reproducing individuals equals the number of individual patches existing on that territory. Losses of patch‑holding individuals are not critical: they are immediately replaced by individuals from the “rover” pool, the population reserve. Territoriality is not the only intra‑population mechanism limiting population growth. For example, in murid rodents fertility regulation according to density is mediated hormonally. At excessive population density, the number of contacts among relatives exceeds a certain threshold, triggering stress responses and hormonal rearrangements that lead to reduced fertility.
From the foregoing one might get the impression that a population is an arena of ruthless, all‑against‑all competition. Of course, this is not the case. Biology knows many examples of mutual assistance and support within a population. It is especially interesting to understand the reasons for altruistic behavior, i.e., behavior in which an individual harms itself or even sacrifices its life for others. The easiest way to grasp such behavior is, for example, a mother protecting her brood. Even if the mother dies, a situation is possible in which the total reproductive value (that is, the future value of the population) of the offspring exceeds the value of the mother herself. For instance, if a gene existed that compelled a mother to sacrifice her life for her children, such a gene could spread in the population thanks to the higher survival of the offspring of altruistic mothers.
The above reasoning fully corresponds to the spirit of sociobiology—the science that explains animal (and, in particular, human) behavior as the result of selection on genetically determined traits. The founder of sociobiology is considered to be the American insect‑behavior specialist Edward Wilson, who published the 1975 book “Sociobiology: A New Synthesis”. The logic of this science is most simply illustrated by the example of kin selection described by one of the classics of sociobiology, William Hamilton. Hamilton was able to explain why altruistic behavior is most often recorded in social Hymenoptera—bees, wasps and ants. The point is that these insects have a rather unusual chromosomal sex‑determination system. Females of these insects can lay unfertilized eggs (with a single chromosome set), which develop into males, and fertilized eggs (with two sets), which develop into females. Usually the genetic similarity between parents and offspring, and even among offspring of the same parents, is 1/2 (half the genes are similar, half are not). However, in Hymenoptera the situation is different. The similarity between a mother and her daughters is 1/2, but the similarity among sisters is 3/4 (because the father contributes a single chromosome set, which all daughters receive in full). This means that a female ensures better survival of her own genes by caring for her sisters rather than for her own daughters! Evidently, these mechanisms are indeed the cause of the formation of social insect families, where one female reproduces (queen or mother), while most individuals are sterile females that care for the queen’s offspring—that is, for their own sisters. The described mechanism that underlies altruistic behavior operates only in females. And indeed, for example, in bees males (drones) do not participate in the collective activities of the colony.
Nevertheless, sociality in insects can arise not only from the described genetic mechanism. Evidence for this is found in termites. Their sociality likely has another cause—collective digestion, in which individuals must repeatedly eat each other’s feces to break down cellulose. By the way, in termites both females and males make equal contributions to “public” work.
While sociobiology explains many remarkable aspects of insect behavior very well, it works somewhat less effectively when it comes to, for example, mammals. This may be due to several reasons, including the fact that in animals with flexible behavior, genes that rigidly determine a particular mode of action in a given situation may be absent. The applicability of sociobiological conclusions to human behavior remains a subject of fierce scientific and pseudo‑scientific debate.
For example, almost all offspring in a troop of baboons belong to several dominant individuals (leaders) who, by pooling their efforts, keep subordinate males in a submissive state (including preventing them from mating with females). Periodically, as the leaders age, “revolutions” occur within the baboon group: subordinates jointly “overthrow” the old leaders and gain priority access to food and females. However, in the face of external danger (e.g., a leopard attack) both dominants and subordinates combine their defensive efforts and often sacrifice their lives to protect the group (see Fig. IV‑18.1). From a sociobiological perspective, a young male who gives his life for the group before he has produced offspring acts “incorrectly”. Yet if we view the situation not from the perspective of selection on individual genes but from the perspective of the male’s psyche as a leader who has not yet attained full leader status, the willingness of the monkey to drive a predator away from the troop at any cost becomes understandable.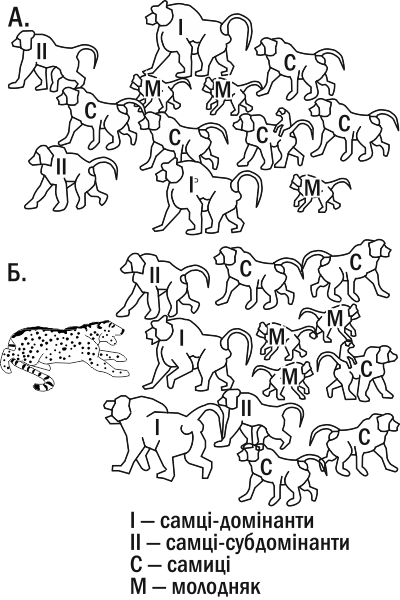
Fig. IV‑18.1. In baboons, a propensity of childless males to self‑sacrifice in defense of the troop has been recorded. The diagram simplifies the derived order of baboons (A.) and their formation during a predator attack (B.).