
Ecology: biology of interactions. V-22. (supplement) Factors influencing organism development
V-22. (supplement) Factors Influencing Organism Development
Heredity, environment, and randomness are the three things that govern our fate.
Akutagawa Ryunosuke
A person's position in society is determined by the trio — talent, labor, and chance.
Volodymyr Shkoda
The close interconnection of organism and environment manifests also in the influence of environment on development. Currently, the notion that hereditary traits of an organism (primarily associated with DNA) establish the "reaction norm" — the range of possible developmental outcomes — has become widespread. The role of environment in these concepts is to determine which variant of this reaction norm will be selected. Variability associated with changes in reaction norm is traditionally considered hereditary, genotypic (mutational) variability and is equated with Darwin's indeterminate variability. Diversity within the reaction norm is traditionally regarded as a manifestation of non-hereditary, phenotypic (modification) variability. It is considered what Darwin called definite variability. The presented description is fully consistent with the viewpoint of the Synthetic Theory of Evolution (STE). In the opinion of the authors of this work, this theory poorly aligns with modern data and significantly lags behind one of its alternatives — the Epigenetic Theory of Evolution (ETE).
First of all, it should be noted that hereditary variability is not at all indeterminate, dependent on ecological need for it. Classic experiments demonstrating that changes in genes are independent of the need for them were conducted by J. Lederberg on intestinal bacilli that lost the ability to break down lactose (milk sugar) due to mutation. Such bacteria were grown on a medium without lactose and then transferred to where this substance was the only food source. Most of these bacteria died, but some (in which a reverse mutation occurred enabling them to feed on lactose) survived. The experimenters showed that the mutation allowing breakdown of the required substance occurred on the original, lactose-free medium, i.e., it was random and undirected.
It seemed everything was clear. But in 1988, Nature published an article by J. Cairns, who only slightly altered the conditions of the classic experiment. In Cairns' experiment, bacteria unable to feed on the new medium did not die. They remained alive, but due to insufficient nutrition could not divide (M.D. Golubovsky, 2001). Under these conditions, bacteria intensively restructured their hereditary apparatus, and some soon acquired the necessary traits. A mutant faced with the choice of dying or returning to the norm manages to "pick up" a reverse mutation and become normal!
The second simplification presented above is that all acquired traits are non-hereditary. At one time, the search for hereditary changes caused by environmental influence was conducted very straightforwardly. For example, August Weismann proved the non-hereditary nature of acquired traits simply by cutting off mice tails. Indeed, over a considerable number of generations, the tail length in descendants of such mice remained constant. I.I. Mechnikov noted on this matter that tormenting mice made no sense: generation after generation, women before leaving descendants lose their virginity, but generation after generation, the hymen (maidenhead) forms again in their daughters. If the results of injuries are not inherited, this does not mean that no traits are inherited at all.
T.D. Lysenko fed cows chocolate at his experimental farm. The cows gave very fatty milk (no wonder!), and Lysenko hoped this trait would transmit to their offspring. It did not. But in 1984, the journal Science published an article by J.L. Marks, in which he mentioned the "Lysenko ghost." When flax plants were grown on soil with excess mineral nutrition, they grew tall and powerful. However, when their descendants were grown on ordinary soil, they still retained part of the parental tallness for many generations. As shown in a number of articles (review — see Grant, 1991), with excess nutrition, additional copies of the gene accelerating growth were incorporated into the flax chromosomes (and stably transmitted to descendants).
In 2005, it was discovered (article by R. Pruitt in Nature) that in ordinary thale cress (or Arabidopsis thaliana), a classic object of molecular genetics, when crossing two individuals homozygous for the defective version of the hothead gene, 10% of offspring have a normal state. Apparently, in this case, it is a matter of DNA correction by RNA molecules that were synthesized on normal genes of the mutant parent plants and stored in their genes.
In 2006, the same journal published the results of work by M. Rassolzegan and other researchers at INSERM in France. They worked with genetically modified brown hamsters (Scotinomys), using a line with an artificially induced mutation of the Kit gene. Hamsters with two mutant versions of the Kit gene die; carriers of mutant and normal versions bear characteristic white marks on their fur; and carriers of two normal genes have (should have!) normal appearance. However, it turned out that external signs of mutation were preserved also in those genetically normal hamsters whose father, mother, or more distant ancestor were hamsters with mutant genes. This phenomenon is called paramutation and consists in the fact that genetically normal but externally altered individuals transmit manifestations of anomaly in offspring for several generations (though with gradual weakening). Apparently, RNA synthesized on mutant genes breaks into fragments but is preserved in the cell, binding with some carriers. Particularly many such fragments are found in sex cells. In cells of genetically normal descendants, these RNA molecules interfere with the work of normal genes and reproduce molecules similar to themselves. Probably, in their presence, RNA synthesized on the normal gene turns out to be abnormal.
Australian immunologists E. Steele, R. Lindley, and R. Blunden in a book with the characteristic title "What if Lamarck Was Right? Immunogenetics and Evolution" (2002) presented a well-founded concept according to which immune cells transmit to gametes information about antigens they have encountered, ensuring inheritance of acquired traits. Although in direct form the assumptions of these authors were not confirmed, there is a characteristic return to ideas that for a century were considered "outdated."
On the other hand, genuine mutations manifest and are inherited very unstably. Their manifestation in individuals with identical heredity is very unstable and varies from their complete manifestation to absence. An exception to this rule is mutations associated with blocking the synthesis of some product of gene activity that is present in normal cells. For example, blocking pigment synthesis will lead to albinism. On the other hand, the appearance of a gene sequence that can ensure synthesis of a new pigment does not necessarily lead to a change in coloration. Proponents of the Synthetic Theory of Evolution (STE) illustrate the action of mutations with examples where a given species has two stable possible developmental pathways and a well-functioning genetic switch between them (pea plants with yellow/green and smooth/wrinkled seeds in Gregor Mendel's experiments). Mutations that arise for the first time do not have such properties.
The instability of mutation embodiment can be observed even on different sides of the body in bilaterally symmetric organisms, or on different segments of metameric beings. The right and left sides, different segments differ from each other in the degree of mutation manifestation. From the viewpoint of the epigenetic theory of evolution, the reason for this is that mutations have not undergone selection for stability of embodiment in ontogenesis.
Conversely, if even unstable traits undergo stabilizing selection, their embodiment in ontogenesis becomes increasingly stable. This was convincingly shown in the experiments of C.H. Waddington. In these experiments, selection of Drosophila for the ability to form dumpy or bithorax aberrations in response to temperature or toxic influences led to them beginning to develop under normal conditions as well (Waddington, 1957). In the experiments of G.H. Shaposhnikov (1978), as a result of selection of aphid aberrations caused by changing the food plant, new, morphologically and ecologically distinctive forms of experimental animals arose, reproductively isolated from the original ones (essentially, a new species of organism was obtained).
As was first shown by R. Goldschmidt, external influence on a developing organism can cause an effect equivalent to a mutation. External influence in the development of a mutant organism can ensure the formation of a normal phenotype. This means that ontogenesis can lead to a fixed number of end states, the choice of which depends both on heredity and environmental influences. For their demonstration, the model proposed by C.H. Waddington is convenient. In this model, organism development is compared to a ball rolling down a surface of complex shape — an epigenetic (epi — above, after) landscape. The emergent property of the development control system is the presence of canalized (from channel — directed path) developmental variants — creodes. On the epigenetic landscape, creodes correspond to depressions (grooves; Fig. V-22.1).
Fig. V-22.1. Waddington's epigenetic landscape. A. — Typical genotype with well-regulated development; the most stable creode leads to the normal phenotype. B. — Genotype change caused restructuring of the epigenetic landscape and enabled development along one of the aberrant creodes. C. — External influence can cause aberrant development even with a normal genotype. D. — External influence can cause normal development even with an altered genotype
The epigenetic landscape models the work of the epigenetic system — the development control system. The epigenetic system can be described as the distribution of probability of different developmental outcomes for a certain value of factors acting on this development (hereditary, associated with DNA; hereditary, not associated with DNA; environmental).
This or that creode is practically never the result of the work of one gene. A system with a very large number of degrees of freedom self-organizes, significantly reducing their number at the output. As early as the 1920s, S.S. Chetverikov found that individuals with normal phenotype are genetically very diverse. For the overwhelming majority of traits, there is no "gene of the norm," normality is determined by the entire genotype (Fig. V-22.2). When establishment of normal phenotype is impossible, aberrations (deviations) appear. Individuals with identical developmental deviations turn out to be genetically diverse (like normal individuals). The fact that individuals with different genotypes develop normally is the consequence of prolonged stabilizing selection for normality. The thing is that the norm is a phenotype corresponding to the most commonly encountered conditions. Despite the mythology of STE, an individual's life success or failure depends not on the presence or absence of a "good" or "bad" allele in it, but on its phenotype as a whole. If from generation to generation, predominantly carriers of normal phenotype survive, this leads to increasingly diverse genotypes beginning to develop normally.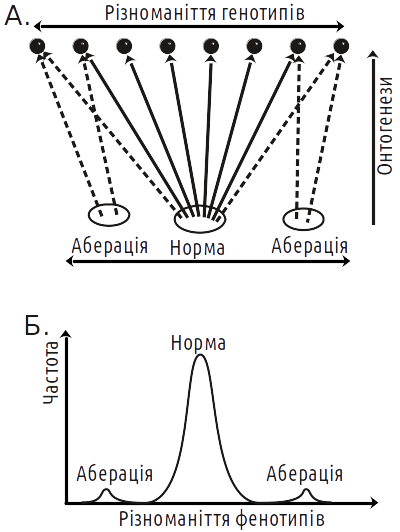
Fig. V-22.2. If ontogenesis were not a self-organizing process, under constant conditions, phenotypic diversity would repeat genotypic diversity (taking into account random environmental influence). The emergent interaction of various factors in the development process leads to the fact that phenotypic distribution differs greatly from normal. The canalization of development leads to the fact that different genotypes correspond to the phenotypic norm. At the same time, the development outcome of each individual zygote cannot be predicted with 100% probability
As can be seen from the examples given, "hereditary" and "non-hereditary" traits are merely extreme points on a single scale with built-up traits whose development as a result of stabilizing selection is regulated better (and occurs in a wide range of conditions) or worse (and occurs only in certain cases). Finally, as the creator of the modern version of the epigenetic theory of evolution, M.A. Shishkin, states, the concepts of genotypic and phenotypic variability generally relate to different situations. Genotypic variability is usually spoken of when comparing the development of different genotypes in identical conditions, and phenotypic — when comparing the development of identical genotypes in different conditions. As experimental data show, in all cases, the developmental outcome of an organism can be predicted only probabilistically, through describing the set of possible developmental outcomes and the probability of each of them.
The main form of selection from this viewpoint is stabilizing selection (Fig. V-22.3). As long as environmental conditions correspond to the norm, individuals that passed through the most probable creode in their development will have maximum chances for survival and leaving offspring. At that, it does not matter what contribution to normal development is assigned to "hereditary" and "non-hereditary" traits: it is determined by the entire epigenetic landscape. "Moving" selection, as its proponents in STE represent it, is a fiction that was never recorded in experiments. Selection against the norm leads to destabilization of the norm and expansion of the variability range. Under changing conditions, this effect precisely makes it possible to "feel out" those developmental variants that can become the new norm. At that, it does not matter on what basis these new variants will develop. Even if their development corresponds to the classic scheme of "modifications" (entirely depends on specific environmental influence), selection for the ability to develop such phenotype will lead to stabilization of the ontogenetic pathway leading to it.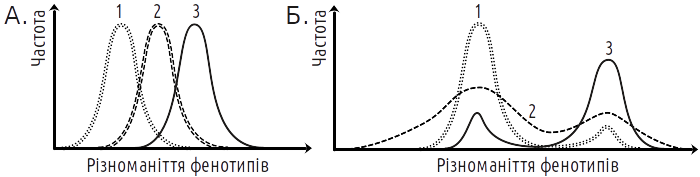
Fig. V-22.3. Selection results (for the greater value of the trait shown on the abscissa axis) according to different views. A. — Result expected from the viewpoint of the Synthetic Theory of Evolution (STE). Selection causes shift of the reaction norm. B. — Result expected from the viewpoint of the Epigenetic Theory of Evolution (OTE). Selection leads to destabilization of development and subsequently — to its stabilization on a new norm. Results recorded in experiments correspond to the second variant to a greater extent